
弥漫性脑创伤后Bcl-2家族与线粒体途径细胞凋亡的相关性分析
2011-08-20 10:40王金林尹立国
中国实验诊断学 2011年12期
王金林,尹立国,李 军
(1.唐山市工人医院,河北唐山 063000;2.遵化市人民医院;3.玉田县第二医院)
随着凋亡机制的深入研究,线粒体在细胞凋亡过程中的中心地位逐渐得到确认,线粒体通透性转换孔(mitochondrial permeability transition pore,MPTP)由于其在线粒体和细胞存活和死亡中的重要作用而备受广大学者关注。近年来的研究显示,bcl-2家族与线粒体膜通透性转换孔关系密切。Bcl-2和Bax是Bcl-2家族中最主要的两个成员,分别具有抑制和促进细胞凋亡的作用,本研究以Marmarou方法建立大鼠弥漫性脑创伤模型,试图通过相关性分析进一步说明其在线粒体途径细胞凋亡中的作用。
1 材料与方法
1.1 脑创伤模型的制备与分组雄性成年SD大鼠90只,体重300-350 g,随机分为实验对照组、颅脑创伤组 。每组设伤后 1 h 、3 h、6 h、24 h 、48 h、72 h 6个时相点。对照组每个时相点3只大鼠,创伤组每时相点6只大鼠。各组未达到规定时相点死亡的大鼠从实验中剔除。动物置乙醚麻醉缸中,以乙醚吸入方式深度麻醉动物,麻醉后的大鼠俯卧于落体致伤海绵垫上,70%酒精消毒颅顶部皮肤,沿正中线矢状切开头皮,剥离骨膜,显露人字缝与冠状缝,将一不锈钢垫固定在大鼠冠状缝与人字缝之间将大鼠头部固定于打击模型架下的头部固定装置中,使重450 g,直径18 mm的铜柱沿垂直玻璃管于1.2 m高度自由落下,撞击大鼠颅骨顶部的钢垫,造成大鼠中度弥漫性颅脑损伤。撞击后即刻移开大鼠,以免铜柱反弹造成头部二次打击伤,给予颅脑损伤后大鼠,头皮切口常规消毒、缝合。对照组仅给予相同的麻醉处理及沿中线矢状切开头皮、剥离骨膜、在冠状缝与人字缝之间粘钢垫,置致伤海绵垫上,但不行自由落体致伤,而后常规消毒、缝合切口.各组未到规定时相死亡的大鼠予以剔除。
1.2 细胞色素c、Bax和Bcl-2检测标本甲醛固定,石蜡切片,按武汉博士德提供检测试剂盒说明书方法进行检测。
1.3 原位细胞凋亡检测(TUNEL法)按Roche公司提供TUNEL检测试剂盒进行操作。
1.4 阳性细胞免疫反应强弱采用Motic Med 6.0计算机真彩色图象分析系统以平均光密度进行半定量分析;实验数据用SPSS11.5 for windows软件进行统计学处理。
2 结果
2.1 伤后1 h至24 hCytc表达与TUNEL阳性细胞数密切相关(r=0.958,P<0.05)(图1)。
2.2 伤后1 h至24 h Bax和Bcl-2二者的比例与Cytc免疫反应相关(r=0.996,P<0.01)(图2)。
2.3 伤后1 h至72 h Bax和Bcl-2二者的比例与TUNEL阳性细胞数密切相关(r=0.925,P<0.01)(图3)。
2.4 但Cytc表达峰值(24 h)早于Bax和Bcl-2二者的比例高峰值(48 h)(表1)。

图1 Cytc表达与TUNEL阳性细胞数的相关性

图2 Cytc表达与bax/bcl-2的相关性
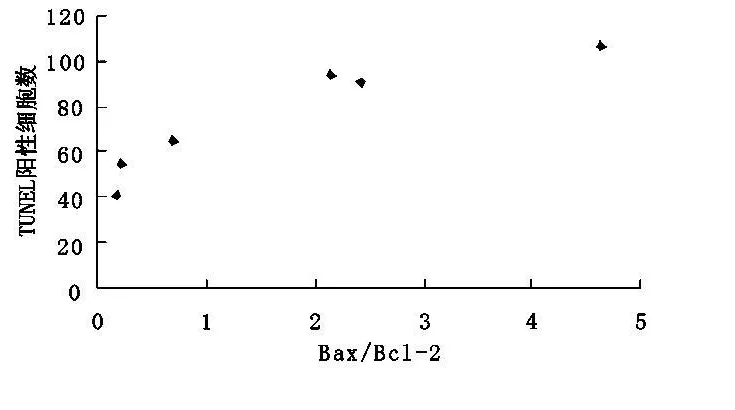
图3 bax/bcl-2与TUNEL阳性细胞数的相关性

表1 脑创伤后 cyt-c、bax、bcl-2与细胞凋亡数的动态变化
3 讨论
Bcl-2属于一类新的癌基因家族,主要包括抗凋亡蛋白(bcl-2、bcl-xl、bcl-wl等)和促凋亡蛋白(bax、bak、bolt等)。近年来研究显示Bcl-2家族蛋白对PTP的开放和关闭起着关键的调节作用,促凋亡蛋白bax等可以通过与ANT或VDAC的结合介导PTP开放,而抗凋亡蛋白bcl-2、bcl-xl等可以通过与bax竞争性的与ANT结合,或直接阻止bax与ANT、VDAC的结合来发挥其抗调亡作用[1,2]。在纯化的线粒体中重组Bax、Bak具有开放PT孔的作用。可能是因为其具有与通道蛋白相似的三维空间结构,能形成足够大的通道来转运ctyc,平时Bax与Bcl-2或Bcl-xl形成异二聚体,使PT处于关闭状态。当细胞发生调亡时,与Bcl-2,Bcl-xl相连的Bax含量增加,形成Bax同型二聚体,PT通道开放,使ctyc释放[3]。目前大量的研究显示细胞内还存在着以bax为核心的不依赖与PTP复合体的凋亡调控机制。体外实验中发现bax能在人工构建的脂质体膜上形成通过大分子物质的孔道而不需要PTP复合体的参与,但必须依赖于bid蛋白和心肌磷脂的存在[4],提示bax可能具有不依赖PTP复合体来介导细胞色素c等物质释放的能力。而在生理状况下bax以单体的形式于胞浆中,研究表明只有在受到凋亡信号刺激后,bax转位到线粒体膜并暴露其BH3结构域后,才能介导细胞色素c释放等下游事件[5]与此同时,相关研究还发现bax单体无法在线粒体膜上形成孔道和释放细胞色素c,只有在形成低聚物后才能介导细胞色素 c释放。进一步的研究显示体外培养的HeLa细胞在外界凋亡诱导因子的作用下细胞内的bax形成了两种结合于线粒体膜的高分子量低聚复合物,而此低聚复合物中并不含有构成PTP复合体的主要成分VDAC和ANT。还有学者发现,发生膜转位的bax在线粒体膜上形成了四聚体到十聚体组成的低聚复合物,同样此低聚复合物中并不含有构成PTP复合体的主要成分VDAC和ANT显著,而bcl-2则可以显著抑制此低聚复合物的形成[6]。另有研究显示bax在凋亡信号介导下发生膜转位后,形成数千个bax分子簇,bcl-xl超表达没有阻止bax膜转位发生,但可以抑制分子簇的形成和凋亡的发生。有此可以看出bax对细胞色素c释放起关键性作用,在凋亡信号刺激下bax从胞浆转位到线粒体膜上从而启动了线粒体介导的调亡途径。同时转位到线粒体膜上bax和bak低聚化或者分子簇形成很可能决定凋亡是否发生。这其中,bax很可能决定凋亡发生与否的必要因素,而bak则是其调控因素[7]。抗凋亡类bcl-2蛋白可以通过阻止促凋亡蛋白在线粒体膜上形成高分子低聚物或是在线粒体膜附近形成分子簇发挥其抗调亡作用。当然上述研究大部分限于体外实验,我们Marmarou方法建立弥漫性脑创伤模型对大鼠体内cytc、Bcl-2、Bax蛋白进行动态观察和相关分析,结果发现伤后1 h至24 h Bax和Bcl-2二者的比例与Cytc免疫反应相关(r=0.996,P<0.01)说明bcl-2家族与cytc释放关系密切,细胞色素c从线粒体释放到胞浆后,在dATP/ATP存在下,首先与凋亡激活因子1(apoptosis protease activeing factor,Apaf-1)结合,形成多聚复合体[8,9]。后者充分聚合胞质中的procaspase-9,并导致其自动活化[10]成caspase-9,然后启动级联反应,继续活化下游的Caspase,如Caspase-3和Caspase-7,整个过程为一正反馈,活化的Caspases能对其底物特异性的切割,DNA片段化,细胞发生凋亡,并出现凋亡形态学改变[11,12,13]。凋亡诱导因子(AIF)从线粒体释放后直接进入胞核内,独立的将DNA裂解引起染色体和DNA的大规模片断化,导致核凋亡[14,15]。我们的相关结果也显示cytc阳性表达与TUNEL阳性细胞数密切相关,Bax和Bcl-2二者的比例与TUNEL阳性细胞数密切相关,结合前人成果可以确认脑创伤cytc释放、bcl-2家族比例变化引发了细胞凋亡。同时我们也对时相变化进行分析,结果发现但Cytc表达峰值(24 h)早于Bax和Bcl-2二者的比例高峰值(48 h),说明另有其它因素作用于线粒体膜通透性转换孔引起cytc释放。
[1]Zomora M,Granell M,Mampel T,et al.Adenine nucleotide translocase 3(ANT3)overexpression inducws apoptosis incultured cells[J].FEBS Lett,2004,563(1-3):155.
[2]Belzacq AS,Helena LAV,Guido K,ed al.The adenine nucleotide translocator in apoptosis[J].Biochimic,2002,84(2-3):167.
[3]Degli Esposti M,Dive C.Mitochondrial Membrane permeabilisation by Bax/Bak[J].Biochemical&Biophysical Communications,2003,304(3):455.
[4]Kuwana T.Mackey MR.Perkins G,et al.Bid,Bax,and lipids cooperate to form supramolecular openings in the outer mitochondrial membrane[J].Cell,2002,111(3):331.
[5]Jia L.Patwari Y.Srinivasula SM,et al.Bax translocation is crucial for the sensitivity of leukaemic cells to etoposide-induced apoptosis[J].Oncogene,2001,20(35):4817.
[6]Mikhailov V,Mikhailov M,Donna J et al.Bcl-2 prevent Bax Oligomerization in the Mitochondrial Outer Membrane[J].J Biol Chem,2001,276(21):18361.
[7]Mikhailov V,Mikhailov M,Degenhardt et al.Association of Bax and Bak Homo-oligomers in Mitochondria[J].J Biol Chem,2003,278(7):5367.
[8]Hu Y,Benedict MA,Ding L,et al.Role of cytochrome c and dATP/ATP hydrolysis in Apaf-1-mediated caspase-9 activation and apoptosis[J].EMBO J,1999,18(13):3586.
[9]Yoshida H.Apaf-1 isrequired for mitichondrial pathways of apoptosis and brain development[J].Cell,2002,94:739.
[10]Hatai T,Matsuzawa A,Inoshita S,etal.Execution of apoptosis signal-regulating kinase-1(ASK1)-induced apoptosis by the mitochondrial-dependent caspase activation[J].J Biol Chem,2000,275(34):26576.
[11]Li Jia,Srinivasa M.Srinivasula,Feng-Ting Liu,et al.Apaf-1 protein deficiency confers resistance to ctyochrome c-dependent apoptosis in human leukemia cells[J].Blood,2001,98:414.
[12]Kawane K,Fukuyama,Yoshida H,et al.Impaired thymic development in mouse embryos deficient in apoptotic DNA degradation[J].Nat Immunol,2003,4(2):138.
[13]Pawlowski J,Kraft AS.Bax-induced apoptotic cell death[J].Proc Natl A-cad Sci USA,2000,97(2):529.
[14]Patterson SD,SD,Spahr CS,Daugas E,et al.Mass spectrometric identification of proteins released frommitochondria undergoing pemeability transition[J].Cell Death Differ,2000,7(2):137.
[15]Susin SA,Daugas E,Ravagnan L,et al.Two distinct pathways leading to nuclear apoptosis[J].J Exp Med,2000,192(4):571.
猜你喜欢
中华养生保健(2020年5期)2020-11-16
中国病理生理杂志(2020年3期)2020-04-03
野生动物学报(2020年1期)2020-02-21
中成药(2018年7期)2018-08-04
中成药(2018年3期)2018-05-07
中国病理生理杂志(2018年6期)2018-01-22
中成药(2017年5期)2017-06-13
中国高原医学与生物学杂志(2017年4期)2017-03-08
中华老年口腔医学杂志(2016年3期)2017-01-15
植物营养与肥料学报(2012年5期)2012-10-26
