
沿黄河下游湿地不同土地利用方式植物多样性1)
2011-08-09 11:07:42韦翠珍张佳宝周凌云
东北林业大学学报 2011年8期
韦翠珍 张佳宝 周凌云
(土壤与农业可持续发展国家重点实验室(中国科学院南京土壤研究所),南京,210008)
湿地作为地球之肾,是地球三大生态系统之一,具有世界上最丰富的物种资源[1]。但是湿地资源的开发利用,使许多生物种尤其是一些稀有动植物种的生存受到威胁。在加拿大的南部,已有50%的湿地用于发展农业生产并建立了排灌系统,由于原生境消失和污染物的增加,使许多湿地动物(包括爬行类、两栖类、鸟类、哺乳类和鱼类)受到影响,湿地中约有1/3野生生物种被列为濒危生物物种[2]。另据统计,荷兰约有50%的湿地资源因受到人为的围垦而遭到破坏;爱尔兰约45%的湿地资源由于人为的破坏而消失[3];波兰由于盲目的利用和开发,境内90%以上的湿地资源已受到严重污染和破坏。中国是世界上湿地类型齐全、数量丰富的国家之一[4],但改革开放以后,由于不合理利用和破坏,湿地的面积急剧缩减,到20世纪90年代中期,已有50%的海滨滩涂不复存在,近1 000个天然湖泊消亡,黑龙江三江平原78%的天然沼泽湿地丧失,七大水系63.1%的河段水质因污染失去了饮用水的功能[5]。这些都不同程度地导致生物多样性的丧失。沿黄河下游湿地是黄河健康生命体系的重要组成部分,其具有蓄水滞洪、净化水体和调节气候的功能,对下游防洪安全起着重要作用;同时其生态环境复杂,适于各类生物如甲壳类、鱼类、两栖类、爬行类及植物繁衍,特别适于珍稀鸟类的生栖[6]。黄河中下游湿地地处暖温带,是亚洲候鸟迁徙的中线,大量水禽在此越冬停歇。河南豫北黄河故道湿地鸟类国家级自然保护区水域、滩涂广阔,野生动植物资源丰富,鸟类众多,是黄河中下游平原人口稠密区交通发达地带遗存下来的较大的一块湿地,动植物中的北方物种、南方物种和广布种十分丰富,是冬候鸟的越冬北界,共有鸟类130余种,其中有国家一级、二级保护鸟类34种[7]。
但是沿黄河下游人口密集,人类农业活动强度大,下游湿地被不同方式利用着,湿地资源受到严重干扰。湿地植物作为湿地构成的三大要素之一[8],是湿地动物生物链的重要一环。湿地不同利用方式严重影响着湿地植物物种组成及物种多样性。研究沿黄河下游湿地不同利用方式下植物物种变化,对黄河下游湿地生物多样性保护、湿地恢复与建设、构建黄河健康生命具有重要的作用。目前,沿黄河下游湿地不同利用方式下植物多样性的研究尚未见报道,为此,文中以河南豫北黄河故道湿地鸟类国家级自然保护区内不同利用方式下植物群落为研究对象,研究沿黄河下游湿地不同利用方式下植物多样性,旨在为中原地区生物多样性的保护提供依据。
1 研究区概况
研究区位于河南豫北黄河故道湿地鸟类国家级自然保护区内,该保护区总面积24 780 hm2,地处河南省新乡市东部,卫辉市和延津县接壤的黄河故道以及封丘县境内的黄河滩涂和背河洼地。前者为古黄河改道后遗留下来的一段槽型凹地,地理位置为东经 114°07',北纬 34°54',长约 30 km,宽约3.5 km,面积 10 500 hm2;后者地理坐标为东经 114°29',北纬35°24',长约 50 km,宽 3 km,面积 14 280 hm2。区内平均气温为14.1℃,年平均降水量为600 mm,区内主要水源为当地降水和浅表地下水。保护区内鸟类资源丰富,有34种鸟类被列为国家重点保护对象,其中国家Ⅰ级重点保护鸟类7种,国家Ⅱ重点保护鸟类27种。该保护区主要保护对象是大天鹅、鹤及湿地生态环境。
2 研究方法
植物群落调查方法:沿黄河下游湿地由滩地、堤根洼地、湖泊湿地、人工湿地(水稻田)组成,其中滩地大多被开垦为耕地,种植农作物,堤根为护堤林(树种为杨树或柳树,胸径22~48 cm,林下均为草本植物)。故本研究选择林下草本、滩地中的耕地(花生地和玉米地)、人工湿地(水稻田)、湖滨湿地为研究对象,于2009年7—8月、2010年7—8月调查沿黄河下游湿地不同土地利用方式下的植物群落,采样方法为生态群落调查样方法。林下草本、玉米地、花生地、水稻田4种利用方式样方面积10 m×10 m,沿黄河大堤,每隔2 km每个利用方式设置一个样方,共40个样方,每个样方内设置3个1 m×1 m的小样方,共120个小样方;湖泊湿地以河南豫北黄河故道湿地鸟类国家级自然保护区的青龙湖湿地为研究对象,以主观采样方法沿青龙湖水深60 cm处往外向保护区边线进行样方调查,湿地植物均为草本植物,故设置的样方面积为1m×1 m,共81个样方。5种湿地利用方式调查总共201个1 m×1 m的样方。调查样地内的植物种类,记录各种植物的名称、植物个体数、盖度、株高。
数据分析:采用生态学上应用最广泛的物种丰富度指数(S)、Simpson 多样性指数(D)、Shannon-Wiener指数(H)、均匀度指数Pielou(E)等来表征植物的多样性特征[9],同时统计各个演替群落的物种总数(ST),计算群落的Sorenson相似性指数(Cs)。重要值(VI)=(相对多度+相对盖度+相对频度)/3;物种丰富度(S)=样地内出现的物种数;Shannon-Wiener多样性指数(H)=-∑piln pi;Simpson多样性指数(D)=∑p2i;Pielou均匀度指数(E)=H/ln S;Sorenson相似性指数(Cs)=2j/(a+b)。式中:pi为第i种植物的重要值;j为两个群落或样地共有物种数;a和b分别为不同两个样地A和B的物种数。
采用单因素方差分析(ANOVA)对群落的丰富度指数、多样性指数、均匀度指数的差异进行检验,ANOVA在SPSS17.0上进行。
3 结果与分析
3.1 沿黄河下游湿地不同利用方式下植物物种组成
沿黄河下游不同利用方式下的湿地由85个植物物种组成,分别隶属于26科66属(表1)。其中林地物种数最丰富,分布有37个物种,占研究区物种总数的43.5%;湖滨湿地次之,有33个物种;滩地中花生地和玉米地物种数相差不大,分别为24、21种,为总物种数的25%左右;人工湿地水稻田物种数最少,仅占物种数的15.3%。沿黄河湿地不同利用方式下植物科别组成中(表2),花生地、玉米地、水稻田等耕地禾本科包含的物种数最多,是该利用方式下的大科植物,菊科、莎草科次之;林地和湖滨湿地中菊科包含的物种数最多,禾本科次之。由此表明,湿地不同利用方式对物种分布有着重要的影响,林地和湖泊湿地两种非耕地的物种丰富,以菊科植物分布为多,人工湿地水稻田物种较为贫乏,花生地和玉米地两种旱作耕地物种数差别不大,包括水稻田在内的耕地中,禾本科植物分布丰富。
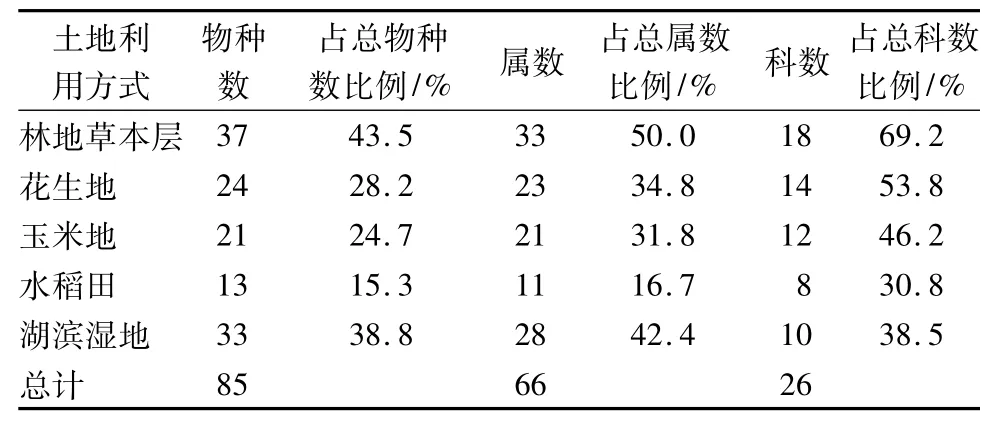
表1 沿黄河下游湿地不同土地利用方式下植物科属种组成
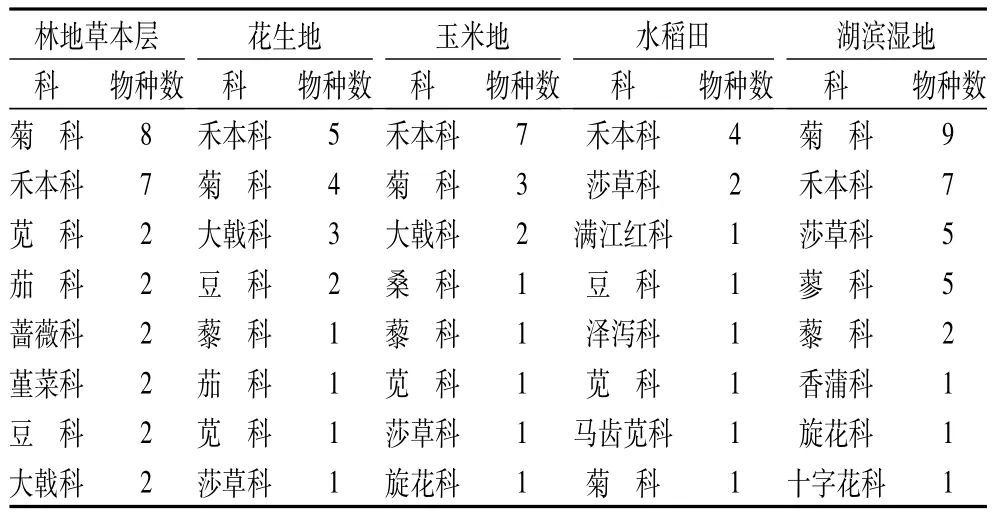
表2 沿黄河下游湿地不同利用方式下植物主要科别组成
3.2 沿黄河下游湿地不同利用方式下植物群落优势种变化
植物种类重要值是由 J.T.Curtis等[10]在森林群落分析中提出来的,其数值大小可作为群落中植物种优势度的一个度量标志,指出群落中每种植物的相对重要性及植物的最适生境。对沿黄河下游湿地不同利用方式下所有植物的重要值进行排序,可以看出各种利用方式下植物群落优势种的变化情况。由表3可知,林地中草本植物以狗尾草为优势种,其重要值为27.7,盐地碱蓬重要值为10.9,也是林地主要物种之一,主要的伴生种包括小飞蓬、藜、苦卖菜、葎草、毛马唐、蛇莓、三叶鬼针草等;湖滨湿地以芦苇为主要优势种,其重要值为26.9,水烛和扁杆藨草亦有较大分布,其重要值分别为21.8、14.6,女菀、莎草、旋覆花、马唐、刚毛荸荠、苣荬菜、酸模叶蓼为主要伴生种;花生地和玉米地的伴生种差别不大,均包括香附子、铁苋菜、醴肠、牛筋草、马唐、马齿苋、打碗花等;水稻田中绿萍生长较为丰富,重要值达到26.1,头状穗莎草、稗、醴肠等为主要伴生种。
3.3 沿黄河下游湿地不同利用方式下植物多样性
香农-威纳指数是根据信息论建立的,表示不确定的程度,Shannon-Wiener指数(H)值越高,不确定程度越大,多样性越高[11],总体而言,沿黄河下游湿地H偏低,不同利用方式H值略有差别,林地最高,但也仅为1.491,湖滨湿地最低,为0.982,花生地、玉米地、水稻田的 H 值为 1.03~1.23。单因素方差分析显示,林地与湖滨湿地之间H有显著差异,花生地、玉米地、水稻田间无统计意义的差异。

表3 沿黄河下游湿地不同土地利用方式主要物种重要值
Simpson指数(D)又称为优势度指数,是多样性的反面即集中性的度量[12],因此该指数在沿黄河下游湿地5种利用方式中的趋势与H相反,林地的D值最低,而湖滨湿地最高,花生地、玉米地、水稻田居中。
物种丰富度(S)即物种的数目,是最简单、最古老的物种多样性测度方法[13]。对于丰富度而言,林地包含的物种最丰富,丰富度指数达到6.1,而水稻田的 S值最低,仅为3.6,花生地、玉米地、湖滨湿地分别为 4.7、4.0、3.8。对物种丰富度进行方差分析,结果表明林地与玉米地、水稻田、湖滨湿地间存在显著差异,但除林地外的其他4种利用方式间物种丰富度差异不大。
均匀度指数(E)是群落中不同物种的多度(生物量、盖度或其他指标)(本文为重要值)分布的均匀程度[12],Pielou均匀度指数为群落实测多样性(H)与最大多样性(Hmax,即在给定物种数下的完全均匀群落的多样性)之比。沿黄河下游湿地不同利用方式下均匀度均为0.8左右,各利用方式间差异不显著。
总体而言,沿黄河下游湿地植物多样性指数较低。
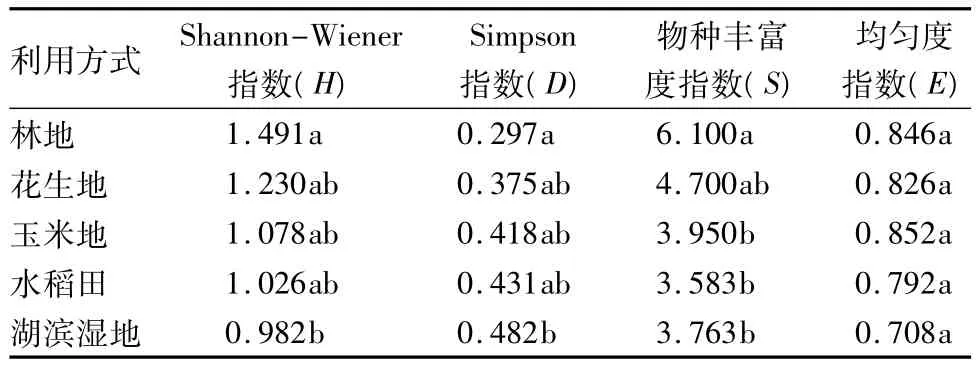
表4 沿黄河下游湿地不同利用方式植物多样性
3.4 沿黄河下游湿地不同利用方式植物群落的相似性
从沿黄河下游湿地不同利用方式植物群落物种组成的相似性来看(表5),花生地与玉米地的群落结构最为相似,Sorenson相似性指数(Cs)达0.596,林地与花生地、玉米地间有一定的相似度,两者间Cs分别为0.444、0.310,林地与水稻田群落结构差异很大,而湖滨湿地与其他4个群落的差异最大,其群落与其他4种利用方式下的群落没有相同的物种出现,Cs均为0。滩地中耕地(即花生地和玉米地)种植植物群落物种组成差别不大,林地土壤含水量与滩地耕地相似,其林下草本群落与耕地物种组成有一定的相似度,但是人工湿地水稻田为短期淹水群落,土壤含水量较高,其群落物种组成与上述群落有较大差异,而湖滨湿地是湿生、沼生、水生植物的理想场所,故其物种组成与其他利用方式下群落差异显著。
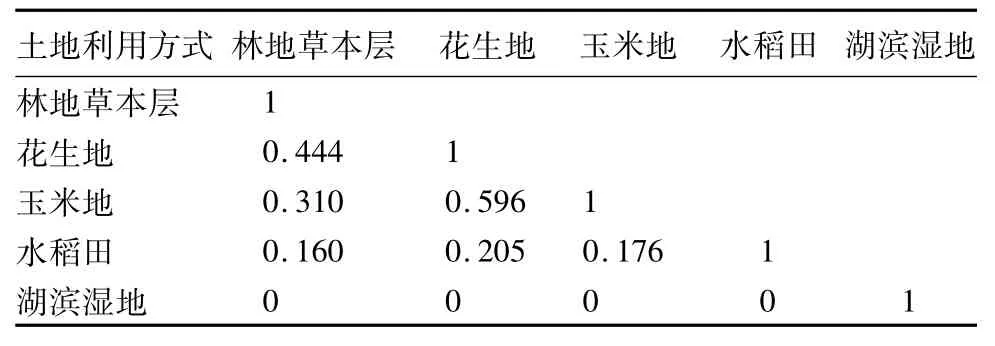
表5 沿黄河下游湿地不同利用方式植物群落相似性
4 结论与讨论
原生植物群落优势种对群落中其他物种的发生具有强大抑制作用,所以不同的群落优势种造成物种组成的差异。湿地的不同利用方式形成了不同的人为形成的植物群落,5种湿地利用方式形成了以林下草本、花生、玉米、水稻和湖滨草本为优势植物的不同群落。花生地、玉米地作为耕地类型,虽然优势种不同,但其主要组成物种差别不大,香附子、铁苋菜、醴肠、牛筋草、马唐、马齿苋等出现的频度较高,这与兰州引黄灌区玉米地[14]、豫北玉米地[15]、江苏旱连作棉田[16]、河北玉米地[17]杂草优势物种组成相类似,均为1年生草本植物,通过种子或根茎进行繁殖,繁殖能力强。林下草本层植物主要物种为狗尾草、盐地碱蓬等植物,均为耐旱物种;而湖滨湿地以喜湿的芦苇、扁杆藨草、水烛为优势种,伴生种亦是喜湿物种,偶见种中野大豆是国家二级重点保护植物,是渐危物种,但沿黄河下游湖滨湿地中野生大豆数量并不多,分布于湖滨湿地边缘。因为它是野生动物和家畜、家禽喜食的饲料,应控制放牧和刈割进行保护。
对5种利用方式下湿地植物群落物种组成相似度分析,旱作耕地群落间相似度高,而湖滨湿地作为天然湿地,物种组成与其他利用方式下植物群落物种组成毫无相似度。农业利用完全改变了沿黄河下游湿地物种组成。
湿地被认为是物种基因库,是物种最丰富的生态系统之一。但是湿地物种消失速度惊人,人为干扰是最主要的原因之一。中度干扰假说认为适度干扰能提高群落物种多样性,但是过度干扰会造成物种多样性的迅速下降[18]。对湿地的利用可以看成是对湿地生态系统的干扰。在研究的5种利用方式中,花生地、玉米地、水稻田是人为耕作土地,除了受到翻耕的干扰外,除草剂的施用亦是对植物群落的最重要的干扰因素,由于耕作方式和土壤含水量的差异,造成了这3种利用方式下植物多样性的变化。对于玉米地而言,除草方式为喷施除草剂,而花生地则为人工除草与喷施除草剂相结合,玉米地Shannon-Wiener多样性指数稍高于花生地,这与姚和金等[19]的研究结果相一致。姚和金等对浙西南柑橘园林下草本层植物多样性对不同除草方式的响应的研究结果表明,化学除草方式杂草多样性最低,人工除草杂草多样性最高。翻耕对植物多样性具有不同程度的影响,马长明等[20]认为深耕(20 cm)有增加Shannon-Wiener多样性指数的趋势,其研究结果认为深耕植物多样性最高,这与本研究结果不一致,相对于玉米地,研究区花生地为免耕地,但是花生地Shannon-Wiener多样性指数略高于玉米地,这可能与除草方式存在密切相关关系。水稻田是沿黄河下游湿地主要湿地类型之一,该湿地水文变化剧烈,同时受到翻耕和除草剂的影响,干扰因素多,强度大,物种丰富度最低。林地草本层和湖滨湿地是沿黄河下游相对比较天然的湿地群落,但是林地是人工次生林地,位于黄河大堤堤根,放牧强度大,而湖泊湿地四周村庄密集,同时受到放牧和人为刈割干扰,湖滨湿地水源为降水和浅层地下水,湿地水文波动大,故研究区林地草本层与湖滨湿地植物群落多样性偏低,但是物种绝对数量依然是湿地非耕地利用类型——林地草本层、湖滨湿地最多。
[1]张丽荣,成文娟,薛达元.《生物多样性公约》国际履约的进展与趋势[J].生态学报,2009,29(10):5636-5643.
[2]Leighton F A.Disease considerations in habitat conservation and management[C]//Holroyd G L.Proceedings of the Second Endangered Species and Prairie Conservation Workshop.Edmonton:Natural History Section Provincial Museum of Alberta,Natural History Occasional Paper,1991:15-33.
[3]周启星.湿地资源保护和合理利用的生态学[M]//陈晓峰.生态环境研究与可持续发展.北京:中国环境科学出版社,1997:15-20.
[4]陈克林.中国的湿地与水鸟[J].生物学通报,1998,33(4):2-4.
[5]赵学敏.湿地:人与自然和谐共存的家园——中国湿地保护[M].北京:中国林业出版社,2005.
[6]郝伏勤,高传德,黄锦辉,等.黄河下游河道湿地浅析[J].人民黄河,2005,27(4):5-8.
[7]王新民,王玉生,郜国玉.豫北黄河故道湿地鸟类自然保护区科学考察与研究[M].郑州:黄河水利出版社,1995.
[8]William J M,James G G.Wetlands[M].4th ed.New Jersy:John Wiley & Sons Inc,2001.
[9]章家恩.生态学常用实验研究方法与技术[M].北京:化学工业出版社,2006:91-92.
[10]Curtis JT,McIntosh R P.An upland forest continuum in the prairie-forest border region of Wisconsin[J].Ecology,1951,32(3):476-496.
[11]尚玉昌.普通生态学[M].北京:北京大学出版社,2002:276-277.
[12]马克平,刘玉明.生物群落多样性的测度方法Ⅰ:α多样性的测度方法(下)[J].生物多样性,1994,2(4):231-239.
[13]马克平.生物群落多样性的测度方法Ⅰ:α多样性的测度方法(上)[J].生物多样性,1994,2(3):163-168.
[14]马丽荣,蔺海明,陈玉梁.兰州引黄灌区玉米田杂草群落及生态位研究[J].草业学报,2007,16(2):111-117.
[15]张爱芹,樊学民.豫北玉米田杂草群落结构及多样性动态[J].中国农村小康科技,2007(8):55-88.
[16]强胜,沈俊明,张成群,等.种植制度对江苏省棉田杂草群落影响的研究[J].植物生态学报,2003,27(2):278-282.
[17]魏守辉,张朝贤,翟国英,等.河北省玉米田杂草组成及群落特征[J].植物保护学报,2006,33(2):212-218.
[18]Connell J H.Diversity of tropical rainforests and coral reefs[J].Science,1978,199:1304-1310.
[19]姚和金,金宗来,杨伟斌,等.不同除草方式对浙西南柑橘园杂草群落及其多样性的影响[J].应用生态学报,2010,21(1):23-28.
[20]马长明,翟明普,袁玉欣.耕翻干扰对丰宁坝上孤石牧场种子库及地上植被的影响[J].草业科学,2006,23(11):73-78.
猜你喜欢
风流一代·青春(2022年7期)2022-07-22 22:57:11
江苏安全生产(2022年5期)2022-06-16 07:51:18
青年生活(2020年31期)2020-10-14 09:07:08
学生天地(2020年17期)2020-08-25 09:28:44
作文成功之路·小学版(2020年5期)2020-06-11 12:48:26
初中生世界·九年级(2016年12期)2017-01-06 14:43:17
西南农业学报(2016年5期)2016-05-17 05:42:32
现代农业(2015年5期)2015-02-28 18:40:49
中国水利(2015年1期)2015-02-28 15:11:49
湖泊科学(2014年2期)2014-02-27 10:30:03
