
四川道路边坡自然恢复的植被多样性研究
2011-06-21 10:14孙海龙李绍才李成俊
水土保持研究 2011年6期
罗 双,孙海龙,刘 冲,李绍才,,庞 亮,李成俊
(1.四川大学 生命科学学院,成都 610064;2.四川大学 水力学与山区河流保护国家重点实验室,成都 610065)
四川道路边坡自然恢复的植被多样性研究
罗 双1,孙海龙2,刘 冲1,李绍才1,2,庞 亮1,李成俊1
(1.四川大学 生命科学学院,成都 610064;2.四川大学 水力学与山区河流保护国家重点实验室,成都 610065)
采用样线法对四川地区路域边坡自然恢复后的植被进行调查,研究道路边坡对植物多样性影响。调查区域内分布植物66科159属190种。其中灌木81种,草本109种;选用了丰富度Margalef指数、Shannon-Wiener多样性指数、Simpson指数、Pielou均匀度指数等α多样性指数及Codyβ多样性指数来研究边坡的坡度、坡向、海拔、恢复时间与植物群落多样性关系。结果表明:45°~60°坡度、阴坡、500~700m海拔及>10a以上的恢复年限的边坡,群落的多样性较高,对边坡植被恢复有利;而在>60°坡度、阳坡、>700m的海拔高度和<10a的恢复年限内的多样性较低;在不同坡度和海拔的环境梯度上,Codyβ指数与α多样性分析结果一致。在生态恢复中,群落多样性低的边坡,适当增加物种丰富度、密度及有利的人为干扰,有助于提高群落多样性。
四川地区;公路生态;自然恢复;植物多样性
在铁路、公路工程建设过程中,经常开挖形成大量裸土边坡,不同程度地损坏原有植被,使得群落结构和多样性及其功能特征发生变化,影响整个区域的生态过程[1],导致道路沿线水土流失、景观破碎、生态抗逆性减弱,生物多样性锐减等系列问题,对自然生态系统产生显著影响,研究表明这种影响涉及到全球陆地面积的15%~20%[2-3]。边坡植被生态恢复是路域生态建设的主要内容之一,在维护边坡稳定实践中,已成为控制侵蚀和稳定边坡的一个有效措施[4],而植物因素起着关键性作用,采用生态工程方式,利用速生物种快速恢复生物环境,但大多形成单一群落或群落多样性低。实践表明,只重视发展人工植被,而忽视保护和恢复天然次生植被,难以达到较好的植被恢复效果[5-6]。植物多样性指数是评价植被覆盖变化的一个重要指标[7],自然恢复下的植物适应性好,植物群落多样性高。在边坡生态恢复时,借鉴当地自然植物群落的结构,保持群落多层次和多样性结构,有利于形成较稳定的植物群落,提高边坡稳定性。
物种多样性指数能较好地反映群落种类组成、结构水平及稳定性和复杂性[8-11],能比较系统和清晰地表现各群落的一定生态习性[12]。植被的多样性研究有利于预测公路边坡植物群落的演替方向、群落稳定性及边坡稳定性。通过不同环境因子与多样性的差别对比,探讨植物群落与边坡之间的关系。现有文献对于路域边坡植被多样性研究大部分针对人工生态恢复,自然恢复情况下植物物种组成及多样性分析等方面的研究文献较少,仅延庆山区公路边坡[13],青藏铁路温性草原区路域[14],长白山区道路[15],四川铁路路堑边坡[16]等有部分对公路边坡自然恢复多样性的研究,但相关研究缺乏大范围内的坡度、坡向、恢复时间等对边坡多样性的影响。本文通过调查四川地区8条公路和铁路,从边坡因素和恢复年限出发,研究对植物物种组成及多样性的影响规律和变化特征,完善边坡植被生态恢复的物种选择和道路建设生态效应理论基础,为边坡人工植被生态恢复提供参考。
1 研究区域与研究方法
1.1 研究区概况
研究区位于四川省成都平原及周边地区(30°15.557′ - 32°15.559′ N,103°32.633′ -105°49.843′E),属亚热带湿润季风气候,年均气温18℃左右,1月平均气温5~7.5℃,7月平均温度25~27.5℃,>10℃年积温5 500~6 000℃,年均降水量1 000~1 200mm,干燥度1.0左右。
1.2 调查方法
由于在自然恢复的条件下,道路边坡植物中乔木很少,同时在边坡的人工生态恢复中,主要参考和选择灌木和草本物种,加之乔木样方较大,受道路沿线边坡条件限制,因此,本文主要对灌草群落与环境因素间(坡度、坡向、海拔、恢复年限)关系进行研究,分析道路边坡植物多样性。
综合考虑道路边坡的特点和调查指标的要求,选择以成都为中心的成渝铁路(成都-重庆)、成南高速公路(成都-南充)、成绵高速公路(成都-绵阳)、宝成铁路(宝鸡-成都)、老川陕公路(四川-陕西)、成洛公路(成都-洛带)、达成铁路(达州-成都)、都汶高速(成都-汶川)等8条公路或铁路,具体信息详见表1。采用样线法,每条道路抽取10个恢复效果较好的边坡植物群落为样点。每个样点设置灌木和草本各3个样方,灌木样方为5m×5m,草本样方为1 m×1m,共计80个样点,草本灌木各240个样方。记录各样点地理位置、海拔、坡度、坡向、物种、株高、冠幅、盖度等指标。本调查于2009年8-11月完成。

表1 四川地区8条道路信息
1.3 数据测定及处理
(1)重要值是某一物种在群落中的地位和作用的综合指数。

(2)物种α多样性。α多样性指数选用丰富度Margalef指数(DMg)、Shannon-Wiener多样性指数(H)、Simpson指数(DS)、Pielou均匀度指数(E)来反映群落水平的综合数值。

式中:S——群落中的物种总数;N——观察到的个体总数;pi——种i重要值[17-18]。
丰富度指数(DMg)是指在一定大小的样方中的物种数目,它是基于物种存在与否,不考虑种间个体数量,忽略富集种和稀疏种对群落多样性贡献的差异[19]。Shannon-Wiener指数(H)是一种较好反映个体密度、生境差异、群落类型、演替阶段的指数。Simpson指数(DS)是反映群落优势度的较好指标,反映各物种种群数量的变化情况,值越小表示优势度物种地位越突出,群落内数量分布越不均匀[20];均匀度Pielou指数(E)描述的是一个群落或生境中全部物种个体数目的分配状况,反映群落内物种个体数间的差异和各物种个体数目分配的均匀程度[21]。
(3)β多样性。β多样性可以定义为沿着环境梯度变化物种替代的程度[22],可直观地反映不同群落间物种组成的差异,揭示生境被物种分割的程度或不同地段的生境多样性,生境被物种分隔的程度[23-25],不同群落间共有种越少,β多样性越大,β多样性与α多样性一起构成了总体多样性或一定地段的生物异质性[22]。采用样地组间物种替代性(Cody指数)来反映β多样性空间分异。

式中:βC——Cody指数;g(H)——沿生境梯度H增加的物种数目;L(H)——沿生境梯度H失去的物种数目,即在上一个梯度中存在而在下一个梯度中没有的物种数目[26]。
每个样点取3个样方平均值。植物多样性分析采用Excel和R软件包处理。
2 结果与分析
2.1 研究区内植物群落特征及组成
本次调查统计植物共190种,隶属于66科159属,植被类型属于亚热带常绿阔叶林。其中灌木38科68属81种,占总种数的42.6%;草本40科90属109种,占总种数的57.4%。菊科(Asteraceae)、禾本科(Gramineae)、豆科(Leguminosae)、蔷薇科(Rosaceae)是种类最多的4个科,占种数的41.4%,表明4大科植物在四川道路植被的自然恢复演替过程中所起的作用最大,在该地区的植物区系中也占据着重要地位。灌木物种种类较多的科为:豆科、蔷薇科、漆树科(Anacardiaceae)、忍冬科 (Caprifoliaceae)、桑 科(Moraceae)、马鞭草科(Verbenaceae)、樟科(Lauraceae)、毛茛科(Ranunculaceae);草本物种种类较多的科为:菊科、禾本科、蔷薇科、伞形科(Umbelliferae)、莎草科(Cyperaceae)、茜草科(Rubiaceae)、唇形科(Labiatae)、豆科、毛茛科;种数最多的属为悬钩子属(Ro-saceae),其次为蒲公英属(Taraxacums)和女贞属(Ligustrum)。物种频度最高为艾蒿(Artemisiaargyi),其次为地果(Ficus tikoua)、黄荆(Vitex negundo)。艾蒿、三叶鬼针草(Bidens pilosa)、飞蓬(Erigeron acer)、短 叶黍 (Panicum brevifolium)、构 树(Broussonetia papyrifera)、马桑 (Coriaria sinica)是调查区域内的先锋物种,与相关研究结果相同[27-28];此次路域植被调查中还发现有入侵物种空心莲子草(Alternanthera philoxeroides)。其中艾蒿、白茅(Imperata cylindrica)、芒(Miscanthus sinensis)、三叶鬼针草、野菊花(Dendranthema indicum)、酢浆草(Oxalis corniculata)、飞蓬、苦荬菜(Ixeris polycephala)、牡蒿(Artemisia japonica)、地果、构树(Broussonetia papyrifera)、黄 荆 (Vitex negundo)、马桑(Coriaria sinica)、铁仔(Myrsine africana)在调查的样方中出现频率较高,是四川地区自然恢复演替过程中主要建群种或共建种,对边坡自然恢复和人工生态修复有重要的参考价值,可在道路边坡人工生态修复中引入,能最大程度上恢复本地植被多样性。
2.2 α多样性指数分析
2.2.1 不同坡度的多样性分析 不同坡度土壤养分含量以及机械组成不同,从而影响植物生长[29]。不同坡度上,草本和灌木的物种丰富度指数(DMg)、Shannon-Wiener指数 (H)、Simpson 多样性指数(DS)及灌木的Pielou指数(E)均在45°~60°表现出峰值,在>60°出现最低值(图1)。不同坡度上,灌木的DMg、H、DS、E变化趋势一致,灌木物种丰富度指数(DMg)仅在<30°的边坡上大于草本,其它坡度灌木层均小于草本层;草本Shannon-Wiener指数(H)均大于灌木,而草本的Simpson多样性指数(DS)仅在>60°的坡度上大于灌木,草本的Pielou指数(E)45°~60°反而最高,均匀程度最高。说明45°~60°坡度上较其它坡度草本和灌木物种丰富性最高,物种数最多,优势物种最少,灌木的均匀程度最高。在>60°坡度上,物种丰富度和多样性最低,优势种(草本:三叶鬼针草、短叶黍、地丁草(Corydalis bungeana);灌木:地果、马桑)地位突出,造成在群落内分布不均匀。在人工生态恢复中,保持45°~60°的开挖坡度有利于提高群落的多样性,而与相关研究结论相似,同时也有研究表明,坡度对物种多样性存在一定影响,但占比重较其它因素小[30-31]。
2.2.2 坡向多样性比较 从表2中可看出,阴坡草本和灌木的各多样性指数(DMg、H、E)都大于阳坡对应的草本和灌木,灌木多样性指数在阴坡和阳坡都低于对应的草本,这表明阴坡较阳坡的物种丰富,多样性高,均匀性高,与相关研究结论相近[32-34],这可能是坡向不同造成光照、水分、湿度等各因子产生差异,从而生境异质性造成物种多样性的差异[15],阳坡坡面温度高,蒸发量大,有机质积累少;阴坡蒸发量小,土壤较肥沃。阴阳坡的优势度差异不大,表明坡向与优势种突出无显著关系。
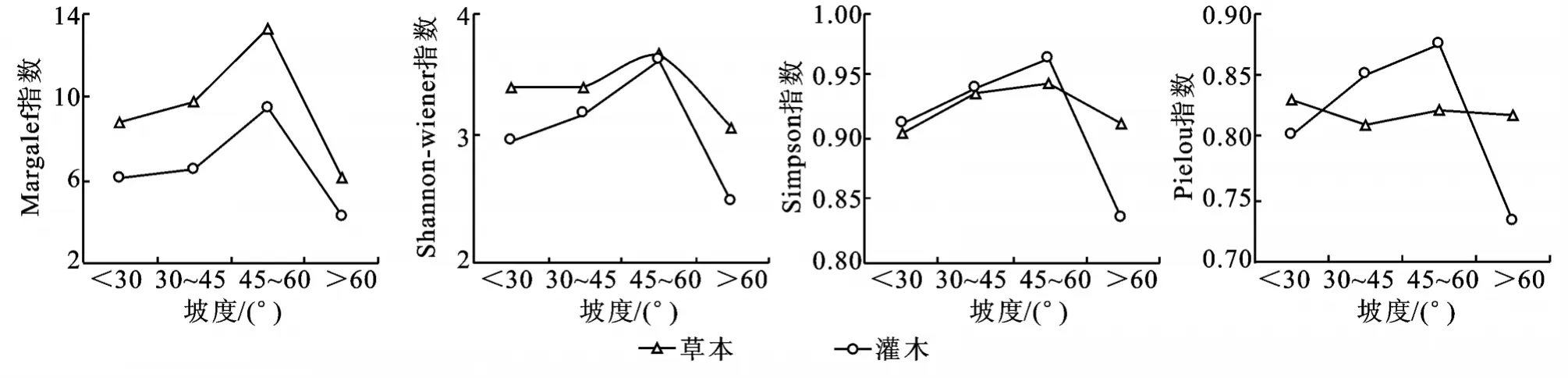
图1 不同坡度自然恢复植物多样性指数

表2 不同坡向的自然恢复植物多样性指数
2.2.3 不同海拔的多样性分析 由图2看出,草本和灌木DMg、H指数在<300m海拔位置均最低,草本和灌木的丰富度指数最高分别在海拔300~500m和500~700m中,说明低海拔的边坡物种数相对较少,多样性相对较低。>500m海拔上,草本与灌木丰富度相差不明显;在>700m海拔上H、DS值最小,多样性最低,优势种(地果、马桑、三叶鬼针草、芒、白茅)突出,均匀程度最低,说明高海拔不利于灌木群落多样性发展。在海拔500~700m,灌木的多样性指数DMg、H、DS、E值最高,且H、DS、E高于对应草本,说明在该海拔灌木比草本多样性程度、均匀性程度高,优势种不明显,灌木层占优势,群落发育较好,导致草本多样性相对降低。300~500m海拔范围内均匀程度低,优势种(地果、构树、黄荆)相对突出,灌木多样性仍能保持较高水平。草本群落则表现相反,在>700m的海拔上,多样性和均匀性最高,无明显优势种,物种数量分布较均匀,但物种丰富度相对较少,在海拔<300m、300~500m与500~700m,草本多样性相差不大,DS、E值低,说明除物种数外,海拔对草本多样性影响不大。高海拔(>700m)和低海拔(<300m)都能使物种丰富度降低,500~700m海拔上灌木多样性和丰富度最高,说明海拔的变化常常是影响山地生境差异的主导因子,海拔导致水势条件及其组合在空间上的分布差异[35-36]。

图2 不同海拔自然恢复植物多样性指数
2.2.4 不同恢复年限的多样性分析 不同恢复时间段上H、DS、E变化趋势相近,在<5a和>15a时间段上草本与灌木的丰富度指数(DMg)差异不明显,草本和木本在>15a时间段上DMg值都最大,而<5a的Margale指数都最小,但在10~15a时间段上反而降低,可能与形成优势种相关。在<5a时间段上,草本和灌木DMg值皆最小,物种数最少,多为先锋物种,但均匀度都最高;草本和灌木在5~10a中H、DS、E值都最小,表明物种多样性最低,优势种(草本白茅、艾蒿、蜈蚣草,灌木构树、黄荆、盐肤木、铁仔)表现明显[37],均匀程度低,但DMg值较<5a和10~15a高。而在10~15a和>15aH、DS、E值较高,与前人研究有相似的结果[13,16],在两个恢复时间段相差较小,多样性程度较高,无明显优势种,群落内物种数量分布均匀。
分析恢复年限对多样性影响的原因,可能是<5a内植被恢复时间短,大部分为先锋物种,具有较高的适应性,草本相对于灌木有较强的适应性,草本多样性高于灌木,但丰富度和多样性较低。而在5~10a处于群落演替竞争消长阶段,草本和灌木层形成了一定的优势种,多样性指数呈现最低,先锋物种为其他物种提供了适应的土壤和水分条件,物种数开始增加[28];10~15a处于趋于稳定阶段,一些无竞争优势的物种被淘汰,造成物种丰富度相对于5~10a时间段反而降低,但多样性提高,优势种也不明显,均匀程度提高;>15a自然恢复的群落结构已经达到稳定,灌木群落占优势,多样性趋于稳定,边坡土壤环境得到改善,使物种丰富性提高,物种数达到稳定状态,这一变化趋势与其他研究[37]相似。结果表明在5~10 a和10~15a两个阶段物种多样性波动较大,是自然恢复的两个重要阶段,10~15a对于形成灌木层更重要,在这个时间段内降低草本的多样性和提高灌木竞争优势,能够提高群落整体的稳定性和抗逆性,这两个阶段内物种的多样性将影响到以后群落物种的多样性和群落的稳定(图3)。

图3 不同自然恢复年限的植物多样性指数
2.3 不同坡度和海拔的群落间β多样性变化
Cody指数(βc)侧重于物种的更替,即新增加和失去物种的数目,用来测度物种在环境梯度上的变化速率,反映群落间物种组成的差异,指数值越高,表示群落的相似性越小,差异性越大,群落间不同的物种越多[38]。从图4中可看出,沿着坡度递增,Cody指数起伏变化较大,在35°、45°左右形成峰值,在38°~45°间有最小值;在30°~50°间曲线波动起伏明显,相邻样地群落间物种替代速率变化悬殊,反映了这个坡度间的生境破碎化程度高,>60°后,Cody指数值急剧减小;在35°和45°左右群落间物种组成相似性大;45°~60°间的群落组成相似性小,群落β多样性高,这与45°~60°的α多样性高相一致。在海拔上,Cody指数在570m左右形成峰值,在300~370m间逐渐递减,在350~370m间最低,在>370m后逐渐起伏递增,波动的幅度较大,表明相邻海拔间各地群落间物种替代速率快慢变化悬殊,反映了随海拔变化边坡的生境破碎化程度高;在>500m海拔Cody指数较高,在570m左右有最大的Cody值,在350~370m群落间物种组成相似程度大,在海拔相对高(>500m)群落间相似性小,相似程度最小,间接说明了该海拔位置的群落β多样性高,这与500~700m的α多样性最高相一致。
结合图1、图3和图4,可以看出一个物种丰富、均匀度高的群落(α多样性高),不一定β多样性就高;反之,一个β多样性较低的群落,未必α多样性一定就低,这说明α多样性独立于β多样性[39]。应当指出的是,由于βc指数仅对物种的存在与否进行测度,而不能对物种水平的数量特征进行测度,使β多样性指数在反映群落内物种沿环境梯度的变化趋势上仍有一定的局限性。另外,生境梯度的变化也并非是均匀的,加之抽样调查的误差等都会在一定程度上对多样性的测度结果产生一定的影响[40],所以关于四川地区的边坡自然植被多样性还有待进一步研究。
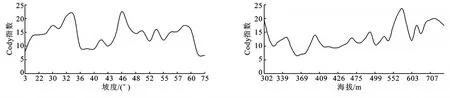
图4 相邻坡度和海拔群落间的Cody指数
3 结论
通过调查分析,四川地区边坡自然恢复的植物群落物种较丰富,统计共有190种,隶属于66科159属,其中灌木38科68属81种,草本40科90属109种。结果表明:在45°~60°坡度、阴坡坡向、500~700 m海拔高度、>10a的恢复年限中,四川地区自然恢复的边坡群落物种丰富性、多样性程度高,均匀性较高;而>60°坡度、阳坡、>700m的海拔高度、5~10a的恢复时间段内的多样性相对较低;高海拔(>700 m)和低海拔(<300m)都能使物种丰富度降低,β多样性在>500m海拔Cody指数较高,在570m左右有最大的Cody值。5~10a和10~15a两个时间段多样性波动较大,是植被自然恢复的两个重要阶段,10~15a对于灌木层形成优势更为重要。在生态恢复应用中,增加频度和优势明显的乡土物种,适当选择45°~60°的坡度,并在5~10a和10~15a恢复时间段内适当控制优势种和增加灌木物种,能提高群落多样性。建议对于导致多样性较低的坡度、坡向、海拔等因素,除了物种选择外,应适当增加物种丰富度和密度,有助于提高群落多样性。
[1]岳天祥.生物多样性研究及其问题[J].生态学报,2001,21(3):462-467.
[2]Shahid N,Li Shibin.Biodiversity enhances ecosystem reliability[J].Nature,1997,390:507-509.
[3]Forman R T T,Sperling D,Bissonette J A,et al.Road Ecology:Science and Solutions[M].Inland:Inland Press,2002:286-293.
[4]Davide G.Biodiversity impact assessment of roads and approach based on ecosystem rarity[J].Environmental Impact Assessment Review,2003,23:343-365.
[5]吴钦孝,杨文治.黄土高原植被建设与持续发展[M].北京:科学出版社,1998.
[6]董林水,张旭东,周金星,等.晋西土石山区森林次生演替过程中群落结构及复杂性[J].应用生态学报,2007,18(3):471-475.
[7]崔莉,田国行,王凯.高速公路绿化生物多样性途径探讨[J].中外公路,2007,27(4):311-313.
[8]常学礼,邬建国.科尔沁沙地沙漠化过程中的物种多样性[J].应用生态学报,1997,8(2):151-156.
[9]彭少麟,周厚诚,陈天杏,等.广东森林群落的组成结构数量特征[J].植物生态与地植物学学报,1989,13(1):10-17.
[10]彭少麟,方炜,任海,等.鼎湖山厚壳桂群落演替过程的组成和结构动态[J].植物生态学报,1998,22(3):245-249.
[11]马克平,黄建辉,于顺利,等.北京东灵山地区植物群落多样性研究.Ⅱ:丰富度、均匀度和物种多样性指数[J].生态学报,1995,15(3):268-277.
[12]朱晓勇,胡海波,鲁小珍,等.太湖西区公路两侧植物物种多样性的研究[J].南京林业大学学报:自然科学版,2006,30(3):85-88.
[13]陈敬贤.延庆山区公路边坡植物群落物种多样性特征[J].中国水土保持,2010(2):3-5.
[14]淮虎银,魏万红,张镱锂.青藏铁路温性草原区路域植被自然恢复过程中群落组成和物种多样性变化[J].山地学报,2005,23(11):657-662.
[15]陈建业,陆旭东,王倜.长白山区公路对路域植物物种组成及多样性的影响[J].生态环境学报,2010,9(2):373-378.
[16]于燕华,辜彬,艾应伟.四川省铁路路堑边坡植物多样性分析[J].中国水土保持科学,2006,4(12):27-30.
[17]马克平.生物群落多样性的测度方法:多样性的测度方法(上)[J].生物多样性,1994,2(3):162-168.
[18]马克平.生物群落多样性的测度方法:多样性的测度方法(下)[J].生物多样性,1994,2(4):231-239.
[19]刘灿然,马克平,吕延华,等.生物群落多样性的测度方法.Ⅵ:与多样性测度有关的统计问题[J].生物多样性,1998,8(6):229-230.
[20]钱宏.长白山高山冻原植物群落的生态优势度[J].生态学杂志,1990,9(2):24-27.
[21]朱守谦.贵州部分森林群落物种多样性初步研究[J].植物生态学报,1987,11(4):286-296.
[22]Wilson M V,Schmida A.Measuring beta diversity with presence-absence data[J].J.Ecol.,1984,72:1055-1064.
[23]Magurran A E.Ecological diversity and its measurement[M].New Jersey:Princeton University Press,1988.
[24]郝占庆,于德永,吴钢,等.长白山北坡植物群落β多样性分析[J].生态学报,2001,21(12):2018-2022.
[25]马克平,刘灿然,刘玉明.生物群落多样性的测度方法.Ⅱ:β多样性的测度方法[J].生物多样性,1995,3(1):38-43.
[26]钱迎倩,马克平.生物多样性研究的原理与方法[M].北京:中国科学技术出版社,1994.
[27]宗浩,苏光麒,张翔,等.四川龙门山铜尾矿库植被调查及植被恢复的物种选择探讨[J].四川师范大学学报:自然科学版,2009,32(6):796-802.
[28]方志强,高信芬,孙成仁.冕宁县山体滑坡地段早期植物群落组成研究[J].西华师范大学学报:自然科学版,2008,29(3):250-257.
[29]吴文轩.大沟流域植物群落物种组成、群落结构及物种多样性的研究[D].重庆:西南大学,2007.
[30]黎云祥,刘钊,陈利,等.南充市近郊退化灌丛草坡群落物种多样性与环境因子灰色关联度分析[J].四川师范学院学报:自然科学版1998,19(2):189-192.
[31]陈光升.四川小寨子沟森林群落物种多样性的环境梯度分析[J].热带亚热带植物学报,2010,18(2):182-188.
[32]李裕元,邵明安.子午岭植被自然恢复过程中植物多样性的变化[J].生态学报,2004,24(2):252-260.
[33]庄树宏,王克明,陈礼学.昆箭山老坟阳坡与阴坡半天然植被植物群落生态特征的初步研究[J].植物生态学报,1999,23(3):238-249.
[34]马克明,傅伯杰,周华锋.北京东灵山地区森林的物种多样性和景观格局多样性研究[J].生态学报,1999,19(1):1-7.
[35]岳明,张林静,高弟,等.佛坪自然保护区植物群落物种多样性与海拔梯度的关系[J].地理科学,2002,22(3):349-354.
[36]Minchin P R.Montane vegetation of the Mt.Field massif,Tasmania:a test of some hypotheses about properties of community patterns[J].Vegetatio,1989,83:97-110.
[37]魏振荣,肖云丽,李锐.巴山山地退耕地植被自然恢复过程及物种多样性变化[J].中国水土保持科学,2010,8(2):99-104.
[38]高贤明,马克平,黄建辉,等.北京东灵山地区植物群落多样性的研究.Ⅺ:山地草甸β多样性[J].生态学报,1998,18(1):24-32.
[39]邱波,任青吉,罗燕江,等.高寒草甸不同生境类型植物群落的α及β多样性研究[J].西北植物学报,2004,24(4):655-661.
[40]白永飞,邢雪荣,许志信,等.内蒙古高原针茅草原群落β多样性研究[J].应用生态学报,2000,11(3):408-412.
Study on Vegetation Diversity of Road Verge after Natural Restoration in Sichuan Province,China
LUO Shuang1,SUN Hai-long2,LIU Chong1,LI Shao-cai1,2,PANG Liang1,LI Cheng-jun1
(1.School of Life Science,Sichuan University,Chengdu610064,China;2.State Key Laboratory of Hydraulics and Mountain River Engineering,Sichuan University,Chengdu610065,China.)
In this paper,we investigated road verge vegetation to study the plant diversity after natural restoration with line transect surveys in Sichuan Province.There are 66families and 159genera of 190plant species,including 81species of shrubs,109species of herbs.Margalef richness index,Shannon-Wiener diversity index,Simpson index,Pielou evenness index and Cody diversity index were selected to examine the relationships among plant community diversity,slope,aspect,elevation,recovery duration.The results showed that habitats with slope from 45°to 60°,the south faced slope,the elevation of 500~700mand the recovery duration>10years slope had higher community diversity,contributing to the vegetation restoration.However,habitats with slope>60°,the north faced slope,at an altitude>700m,and the recovery duration<10 years had the lower diversity.In addition,Cody index was used to analyzeβ-diversity along the slope and elevation environmental gradient,which was consistent with the results ofα-diversity.During the course of ecological restoration of road verge vegetation in this region,habitats with lower diversity resulted from adverse environmental factors,well-adapted species,higher species richness and density,with additional artificial interferences should be adopted to enhance the community diversity.
Sichuan Province;road ecology;natural vegetation restoration;plant diversity
X171.4;X176
A
1005-3409(2011)06-0051-06
2011-05-31
2011-06-24
国家科技支撑计划重点项目课题(2007BAD39B04);环保公益性行业科研专项(200909060)
罗双(1986-),男,湖南省长沙人,硕士研究生,主要从事生态工程和生态恢复方面研究。E-mail:luoshuang324@163.com
孙海龙(1976-),男,黑龙江省海林市人,博士,讲师,主要从事生态工程方面研究。E-mail:lizist@vip.sina.com
猜你喜欢
青年文学家(2021年16期)2021-08-05
少儿美术(2020年8期)2020-11-06
城市道桥与防洪(2019年5期)2019-06-26
中国特种设备安全(2019年4期)2019-05-20
中国特种设备安全(2018年11期)2019-01-08
中央民族大学学报(自然科学版)(2018年4期)2018-03-19
乡村科技(2018年8期)2018-02-13
现代园艺(2017年13期)2018-01-19
诗潮(2017年2期)2017-03-16
医学研究杂志(2015年4期)2015-06-10
