
杉木3种交配设计试验种子品质的遗传分析
2011-06-19 04:42石志玲陈晓阳
华南农业大学学报 2011年4期
杨 萍,石志玲,陈晓阳
(1黎平县国营东风林场,贵州黎平557300;2黎平县国营花坡林场,贵州黎平557300;3华南农业大学,广东省森林植物种质创新与利用重点实验室,广东广州510642)
揭示种子品质性状变异规律及其与遗传基础的关系,对于提高林木种子园种子品质具有重要的意义.杉木 Cunninghamia Lanceolata(Lamb.)Hook是我国南方重要的工业用材树种,我国于20世纪60年代开始营造杉木种子园,迄今,种子园仍然是杉木良种繁育的主要途径.然而,杉木种子园种子播种品质不尽人意.据统计,各地杉木种子园的健全种籽率一般在30%左右,另外60%左右的种子是空籽或涩籽[1-8].如何改善种子的播种品质一直是杉木种子园经营管理中的重要问题.20世纪80年代初,叶培忠等[1-2]和陈岳武等[3]就杉木种子播种品质是否受遗传控制、双亲哪方影响更大等问题进行了研究,后来还有一些相关研究对影响杉木种子播种品质的因素进行了观察分析[4-8].但由于试验规模、试验设计和亲本不同等原因,结论并不完全一致.为进一步研究不同交配方式对种子品质的影响,本文利用贵州省黎平县国营东风林场杉木种子园不同交配设计试验的种子检验结果进行统计分析,试图验证杉木杂交中异交、自交、一般配合力(GCA)、特殊配合力(SCA)对杉木涩粒率、空籽率的影响,为进一步提高种子播种品质提供科学依据.
1 材料与方法
1.1 试验方法
亲本材料均为贵州省黎平县国营东风林场杉木种子园子代生长优良的无性系.对5×5全双列交配、11个亲本的不连续半双列交配和6×5的测交系交配试验的种子,采用X线法检验和剖切法检验进行种子涩籽、空籽和健全籽检验.检验重复4次,每个重复约含种子100~200粒.
1.2 统计方法
在林木交配试验中,经常遇到组合缺失问题.为了更为客观和精确分析,本文采用一般线性模型,通过求解因素效应值,进而求出各因素的离差平方和(SS)及均方,并通过一般表型方差分量模型,求出各因素均方成分[9-10].其中,双列交配设计的方差分析方法适用于该类各种交配试验分析.采用Turbo C 2.0编程软件自编程序计算,并利用实例进行了检验.对空籽率等百分率数据采取反正弦转换后再进行统计分析.
配合力分析采用表型线性方差分量模型:
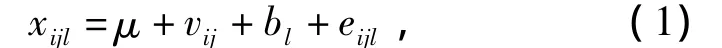
式(1)中,μ为总平均;vij为组合效应;bl为重复效应;eijl为机误.
全双列交配的组合效应可按式(2)分解,包括一般配合力(gi)、特殊配合力(sij)和正反交效应(rij):

测交系交配试验的组合效应可按式(3)分解,包括父本一般配合力(gMi)、母本一般配合力(gFi)和特殊配合力(sMFlj):
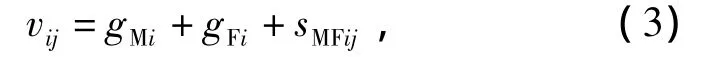
式(1)~(3)模型有如下假设条件:
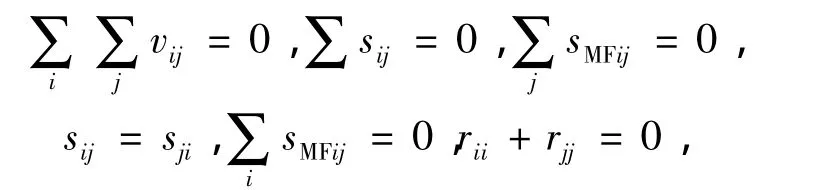
据此,可求出各因素的效应值.
本文配合力方差分析均采用固定模式,方差分析模式见表 1、表 2.表 1、2中,Kx=(tr(Ux×C'x×A-1)×Cx×Ux)/υx,σ 为 x均方成分,x:分别取 6、v、g、s、r、f和 m;Ux:x 因素效应值的设计矩阵;Cx:x因素效应值的设计矩阵变换矩阵;Cx':Cx的转置矩阵;A-1:x因素效应值的系数满秩逆矩阵;υx:x因素的自由度;tr:矩阵求迹.

表1 双列交配设计方差分析表1)Tab.1 Variance analysis of sowing traits of seeds derived from diallel mating
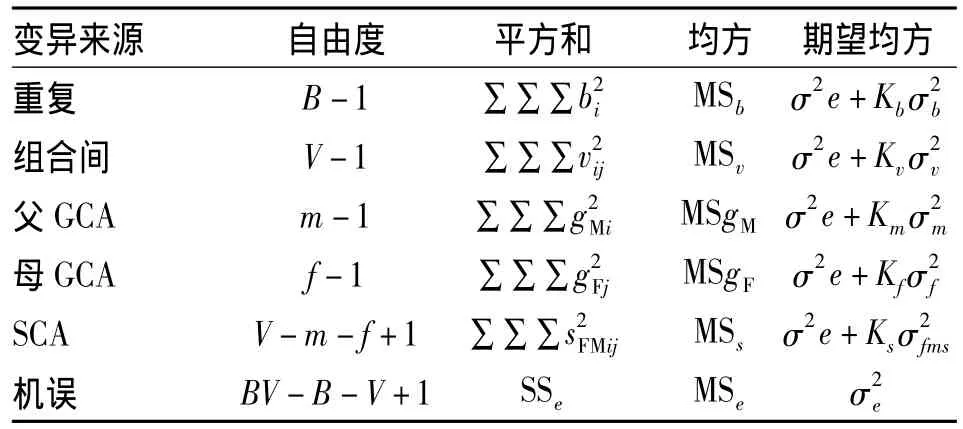
表2 测交系交配设计方差分析表1)Tab.2 Variance analysis of sowing traits of seeds derived from tester mating
2 结果与分析
2.1 完全双列交配试验
从表3可以看出,不同交配组合种子在健全籽率、空籽率、涩籽率和种子千粒重上的差异很大.例如,交配组合4×4的种子平均健全籽率仅0.5%,而4×74高达48.4%,两者相差95.8倍;96×74的空籽率为0.4%,而 4×96高达 60.3%,两者相差150.8倍;4×96的涩籽率仅10.0%,而4×4高达88.4%,两者相差7.8倍;88×74的千粒重2.925 g,而96×88为5.940 g,两者相差1.03倍.不同母本间的种子品质也有明显的差异.96等无性系的种子品质较好,而89等较差.同一母本不同组合间的差异也很大.如无性系4与其他4个父本交配,健全籽率为8.7% ~48.4%,空籽率为8.6% ~60.3%,涩籽率为10.0%~82.7%.这说明种子播种品质在很大程度上取决于具体的交配组合.从表3还可看出,正交与反交的差异也很大.如4×74和88×89的健全籽率分别为48.4%和21.4%,而74×4和89×88的健全籽率分别为16.6%和14.8%.
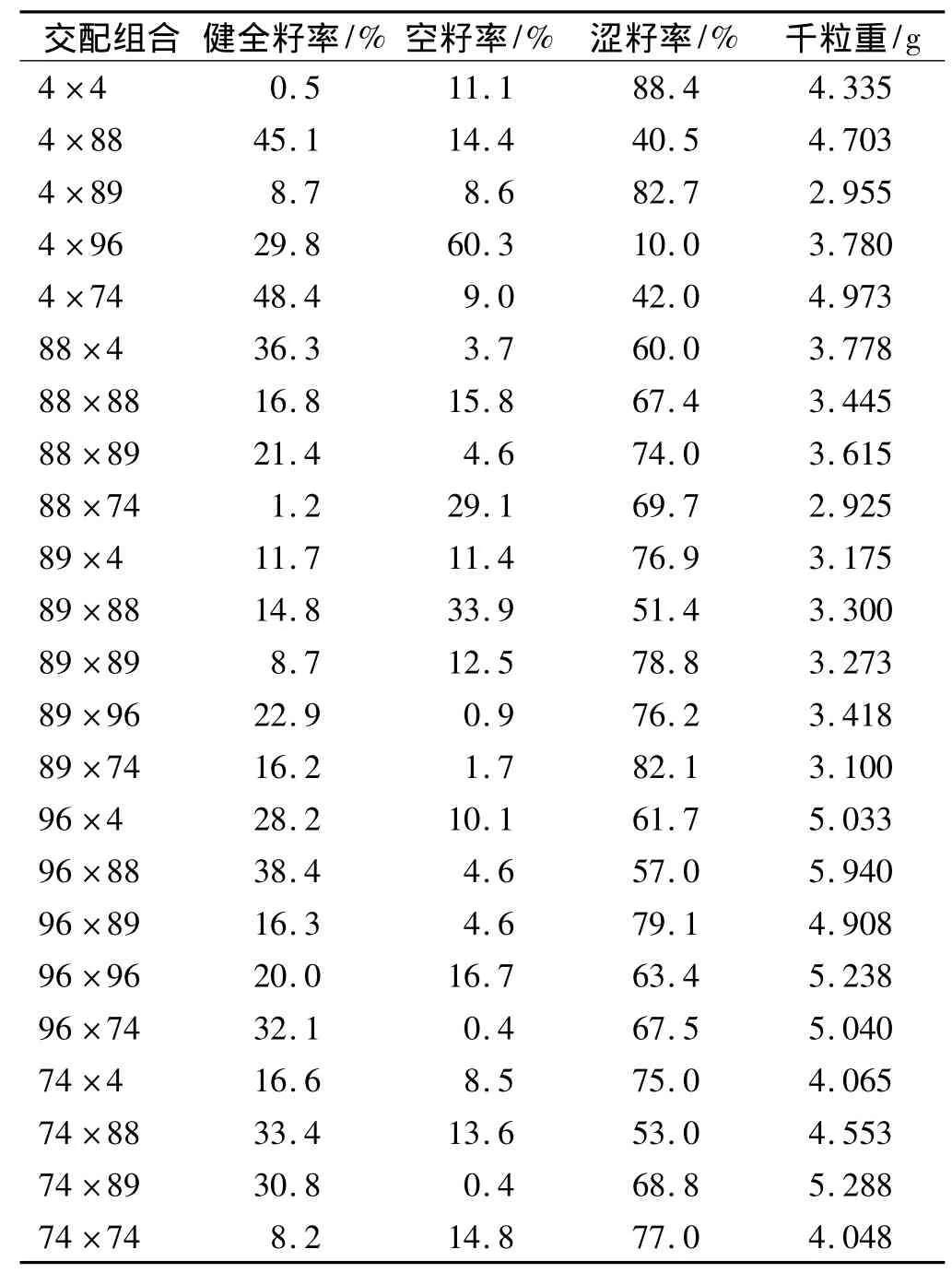
表3 全双列交配种子检验结果Tab.3 Sowing traits of seeds derived from complete diallel mating
经方差分析(表4),组合间在各指标上的差异达到极显著水平,亲本间GCA和SCA的差异均达到了极显著水平,从方差分量来看,两次试验各指标的SCA都高于GCA.正反交差异达到了显著或极显著水平,尤其是空籽率和千粒重,除遗传原因外,还可能与授粉状况有关.
2.2 不连续半双列交配试验
不连续半双列交配试验结果也表明,不同组合差异也非常大,健全籽率、空籽率、涩籽率和千粒重分别变动于 4.4% ~33.2%、13.2% ~68.0%、21.3% ~61.9%和0.492~l.156 g.种子品质与母本有明显的关系.例如无性系63作母本,健全籽率为20.8%,明显高于其他无性系,无性系93和121种子品质较差,健全籽率平均只有8.5%和9.4%.经方差分析(表5),种子品质各指标在交配组合、GCA和SCA都达到极显著水平,GCA的F值大于SCA.除涩籽率外,GCA的方差分量比SCA的大,这与全双列交配试验结果有所不同.
2.3 测交系交配试验
由表6可以看出,30个组合间健全籽率、空籽率、涩籽率和200粒重的变动幅度分别为11.0%~45.3%、14.0% ~58.8%、24.5% ~57.0%和0.681 ~1.497 g.母本间的差异更明显,如593无性系的健全籽率平均为18.82%,而594无性系平均为37.1%,两者相差18.3%.
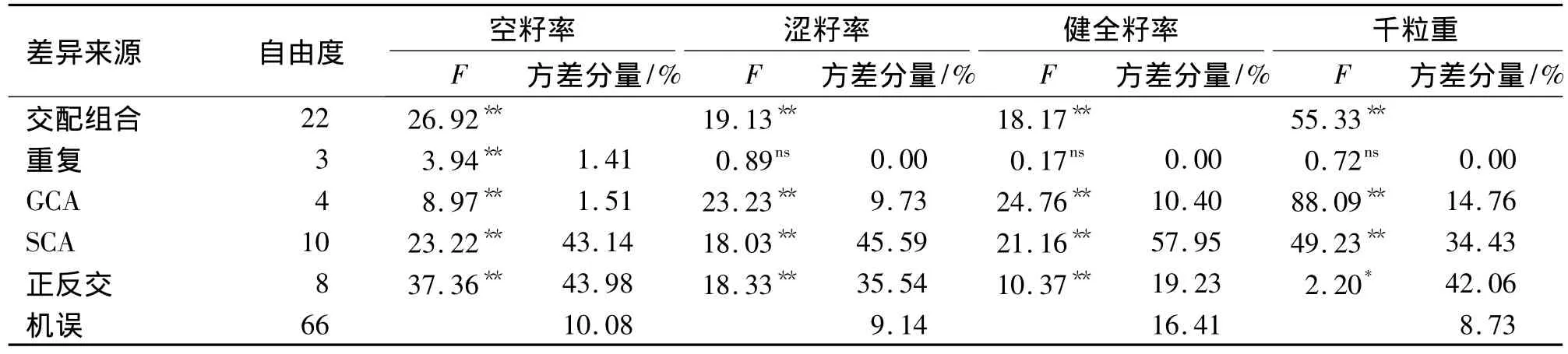
表4 全双列交配种子检验数据方差分析1)Tab.4 Variance analysis of sowing traits of seeds derived from complete diallel

表5 不连续半双列交配试验种子检验数据方差分析1)Tab.5 Variance analysis of sowing traits of seeds derived from disconnected half-diallel mating
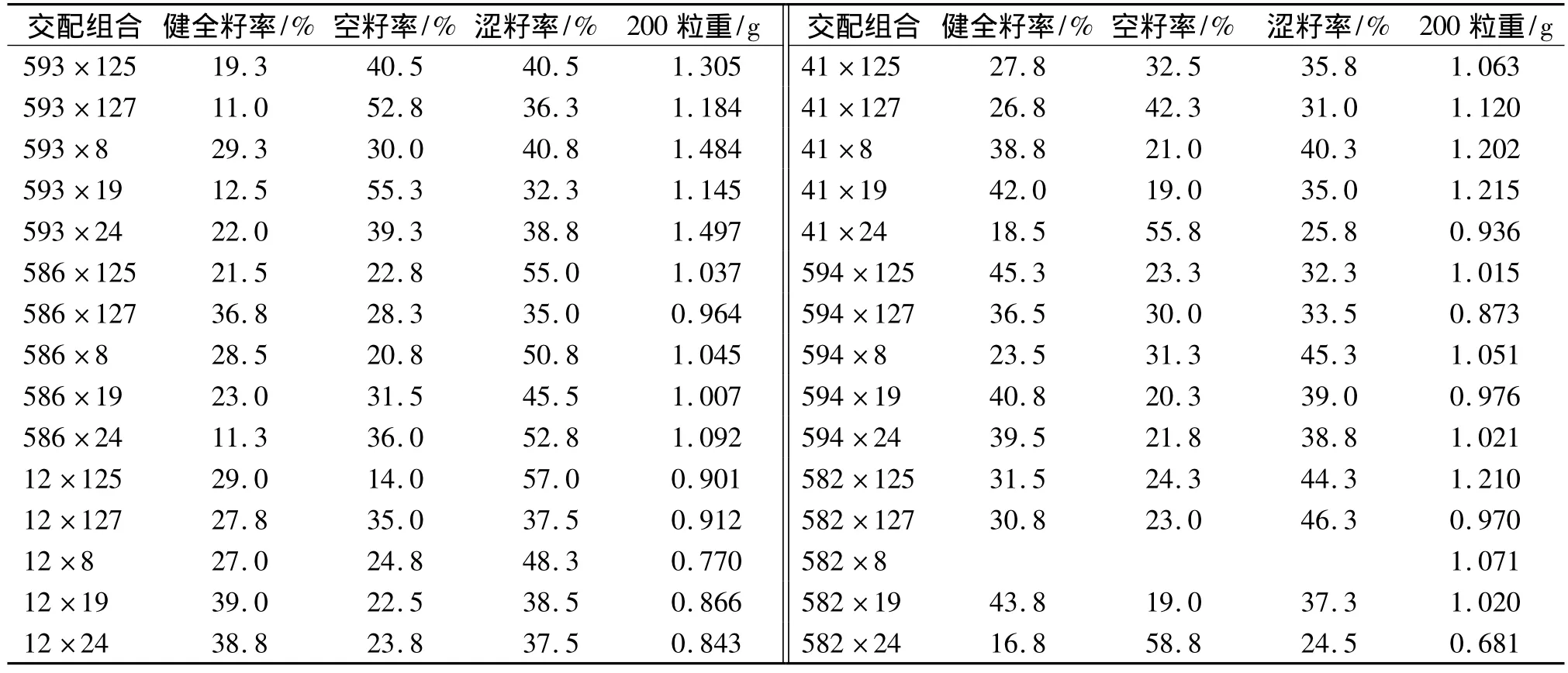
表6 测交系交配试验种子检验Tab.6 Sowing traits of seeds derived from tester mating
方差分析(表7)表明,种子品质各指标在交配组合、父本和母本的GCA以及SCA间的差异都达到了极显著水平.从方差分量可知,母本的GCA均大于父本的GCA,父本和母本的GCA方差分量总和大于SCA.
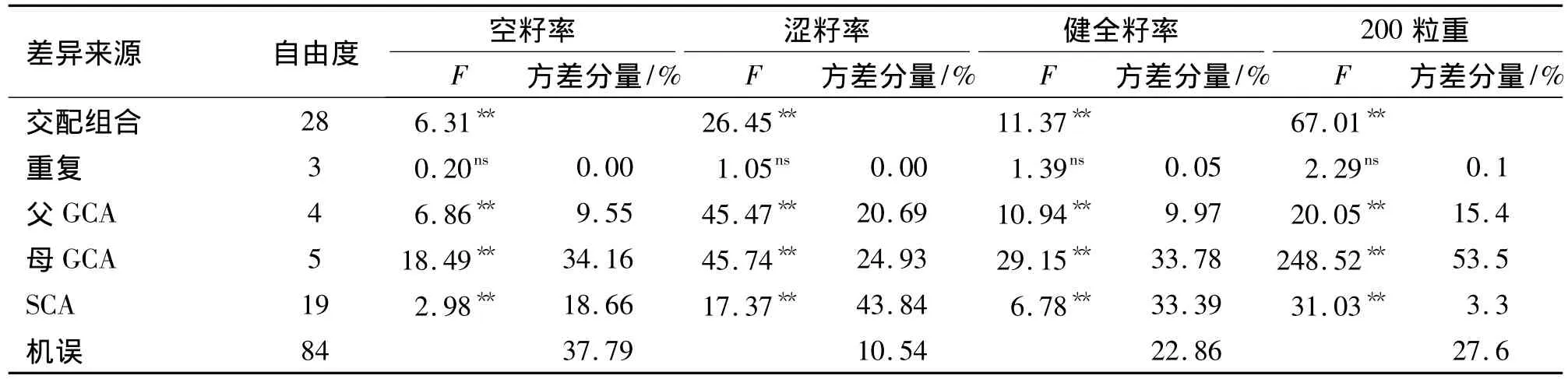
表7 测交系交配试验种子检验数据方差分析1)Tab.7 Variance analysis of sowing traits of seeds derived from tester mating
2.4 自交对种子品质的影响
表8数据表明:A组试验中,自交的平均健全籽率比异交的低15.4%,平均空籽率比异交的高4.4%,平均涩籽率比异交的高11.6%.B组试验也反映异交的种子品质比自交好.不同无性系自交能孕性是有差异的.如在两组试验中,无性系88健全籽率均高于89无性系.由表8还可看出,自交的涩籽率明显高于异交,但在空籽率上,两种交配方式相差不明显.
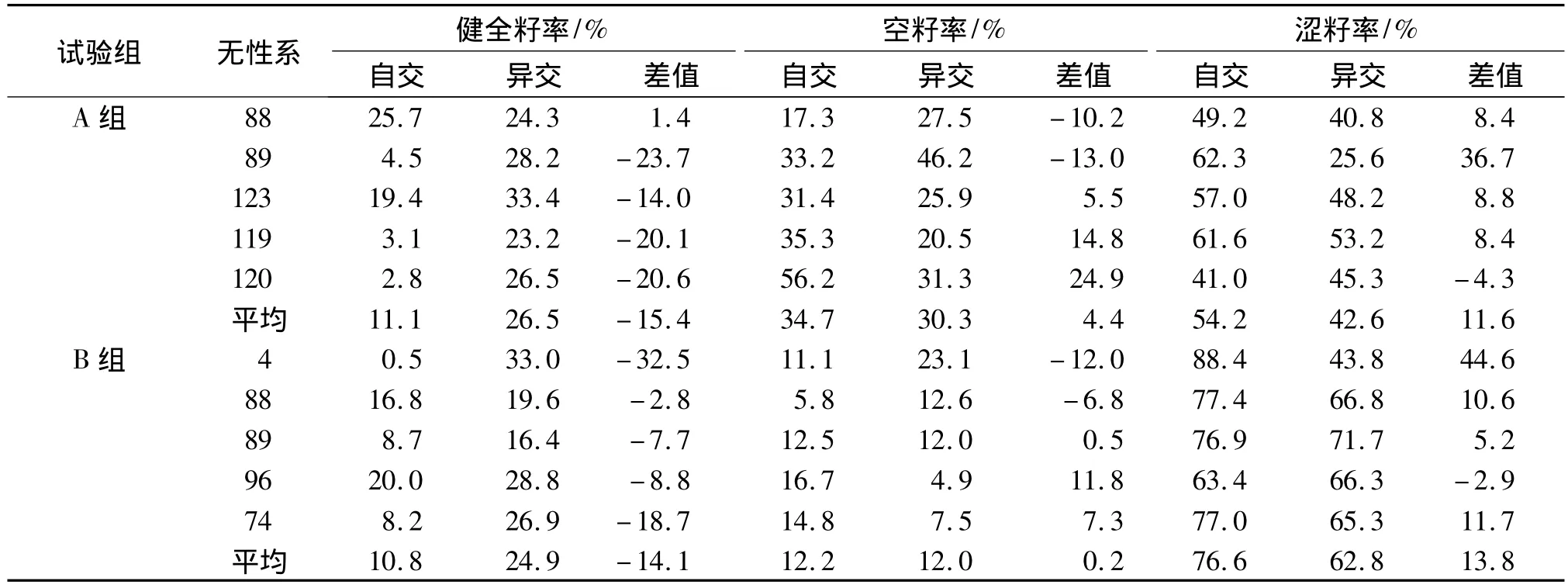
表8 自交与异交的种子品质Tab.8 Sowing traits of seeds derived from selfing and outcrossing
对18和19无性系做了本系花粉不同比例的授粉试验,结果(表9)表明,本系花粉比例与种子千粒重、空籽率关系不明显,而与健全籽率和涩籽率相关紧密,随着本系花粉比例增高,健全籽率降低,涩籽率提高,自交是通过产生涩籽来影响种子的播种品质.由表9还可看到,自由授粉的健全籽率和涩籽率十分接近本系花粉占20%的授粉试验结果.据此可以估计,3年生1.5代种子园的自交率达20%左右.

表9 本系花粉比例与种子播种品质Tab.9 Quality evaluation of seeds pollinated with various proportions of self-pollen
3 讨论与结论
自交降低种子品质和产量已在花旗松Pseudotsuga menziesii、欧洲云杉 Picea abies、欧洲赤松 Pinus sylvestris、火炬松 Pinus taeda、湿地松 Pinus elliottii、油松Pinus tabuleaformis等树种中得到证实[11].叶培忠等[1]通过杉木3×3全双列交配试验表明,各亲本自交时,其健全籽率比异交降低20% ~60%,比自由授粉降低50% ~90%,其涩籽率比异交增高40% ~100%,比自由授粉增高34% ~110%,空籽率与交配方式关系不明确.本研究也表明,自交的涩籽率明显高于异交,但在空籽率上,两种交配方式差异不大.由于自交是通过生成涩籽来影响种子的播种品质,因此降低自交率可以显著提高种子的播种品质.
叶培忠等[1]根据杉木交配试验分析认为,杉木种子活力主要受基因的显性作用,陈益泰等[4]研究显示,杉木近交使得种子发芽率和苗木高生长量大幅度下降.近交衰退随着近交系数的增加而加大.Johnsson[11]对欧洲赤松空籽率研究得出,特殊配合力是主要的.本文的3个试验结果分析均表明,GCA、SCA和正反交效应都显著.但是,加性效应与非加性效应哪个更大,不同交配试验的结论不一致.全双列交配试验反映SCA方差分量大于GCA方差分量,即非加性效应大于加性效应.而测交系交配试验结果显示,GCA方差分量较大,加性效应更明显.为了解释这个问题,将全双列交配试验自交组合剔除后,再作统计分析,结果表明,空籽率GCA和SCA的F值分别为29.86和10.33,方差分量分别为35.87%和24.38%,涩籽率GCA、SCA的F值分别为38.14和4.08,方差分量分别为25.14%和8.92%.这表明,GCA的方差分量大于SCA.由于全双列交配设计试验中包含了自交,而自交组合种子品质较差,因而SCA的方差分量相对较大,但当剔出自交组合后,结论恰好相反,即GCA的方差分量大于SCA.此外,由于不同试验用不同的亲本,而不同亲本选择受精的状况有一定差异,因而结论也有所差别.
[1]叶培忠,陈岳武,蒋恕,等.杉木种子生活力变异的研究[J].南京林产工业学院学报,1981,5(3):22-32.
[2]叶培忠,陈岳武,刘大林,等.配合力分析在杉木数量遗传研究中的应用[J].南京林产工业学院学报,1981(3):1-21.
[3]陈岳武,施季森,刘大林,等.杉木种内杂种优势及亲本配合力的分析[J].南京林产工业学院学报,1982(2):1-20.
[4]陈益泰,何贵平,李恭学.杉木种子发芽率和苗木高生长的近交效应[J].林业科学研究,1989,2(5):420-426.
[5]刘仁东,陈晓阳,秦向华.气象因子差异对杉木种子质量的影响[J].四川林业科技,2007,28(3):78-79.
[6]吴承祯,洪伟,林思祖.杉木种子涩籽的空间特征分析[J]. 山地学报,2006,24(1):117-122.
[7]刘芳.无性系、立地、郁闭度对杉木种子园产量和质量的影响[J].林业科技开发,2002,16(5):24-26.
[8]孙鸿有,郑勇平,翁春媚,等.杉木种子园种子品质性状变异及遗传参数[J].浙江林学院学报,2005,22(1):61-65.
[9]NAMKOONG G.Introduction to quatitative genetics in forestry[M].[S.l.]:Castle House,1981.
[10]王松桂.线性模型的理论及应用[M].合肥:安徽教育出版社,1987:348-444.
[11]JOHSSON H.Contributions to the genetics of empty grians in the seed of pine(Pinus silvestris)[J].Silvae Genetica,1976,25(1):10-15.
猜你喜欢
农业科技通讯(2023年1期)2023-02-12
辽宁林业科技(2021年3期)2021-07-08
四川蚕业(2021年1期)2021-02-12
山西水土保持科技(2020年2期)2020-12-22
昆虫学报(2020年6期)2020-08-06
昆虫学报(2020年1期)2020-03-03
猪业科学(2018年5期)2018-07-17
河北农业科学(2016年2期)2016-09-09
广西林业科学(2016年1期)2016-03-20
广西林业科学(2016年1期)2016-03-20
