
Bt棉对斑痣悬茧蜂寄主选择及其子代发育的影响*
2011-06-12 02:44王锦达王以一刘沛菡李保平
植物保护 2011年3期
王锦达, 王以一, 刘沛菡, 王 康, 李保平
(南京农业大学植物保护学院昆虫学系,农业部作物病虫害监测与防控重点开放实验室,南京 210095)
在棉田生态系统中,天敌昆虫不仅对优势害虫(包括靶标害虫)种群数量具有抑制作用,而且是若干非靶标害虫维持较低水平的重要因素。Bt棉的广泛种植对其靶标害虫棉铃虫(Helicoverpa armigera Hübner)具有很好的控制作用[1],但对棉铃虫寄生蜂也有不同程度的影响,例如,多项研究报道Bt棉田中棉铃虫幼虫的寄生率明显降低或寄生蜂数量减少[2-3]。其原因一般推测为,当转基因作物对靶标寄主幼虫具有很强的致死作用时,则必然对其寄生蜂产生不利影响[4]。但如果Bt作物对寄主幼虫(如非靶标害虫)具有亚致死作用,寄生蜂在寄主幼虫死亡前完成发育,则寄生蜂的繁殖未必受到不良影响,种群数量未必明显降低。例如,Bt棉对棉蚜茧蜂[Lysiphlebia japonica(A shmead)]的寄生率影响不大[5];崔金杰等[2]分别从双价棉、Bt棉和常规棉田采回被寄生的棉蚜僵蚜,羽化率均很高(90%以上),羽化出蜂体重无明显差异;但缘腹盘绒茧蜂(Cotesia p lutella)和佛州点缘跳小蜂(Copidosoma f loridanum)寄生大豆尺夜蛾(Pseudop lusia includens)幼虫后,寄生蜂的生长发育、生殖力和寿命等适合度特性受到不良影响[6],类似的结果也出现在寄生棉大卷叶螟[Notarcha derogata(Fabricius)]的螟蛉盘绒茧蜂(C.ru ficrus)[7]寄生蜂中。显然需要更多的研究来阐明,Bt棉的非靶标害虫对其寄生蜂、尤其是与靶标害虫共有的寄生蜂的影响。
斜纹夜蛾[Spodoptera litura(Fabricius)]和甜菜夜蛾[S.exigua(Hübner)]为转Bt棉的非靶标害虫[8],近年来在部分地区的转Bt基因棉田严重为害棉花[9-10]。斑痣悬茧蜂[Meteorus pulchricornis(Wesmael)]寄生多种大型鳞翅目幼虫,不仅是棉铃虫、而且是斜纹夜蛾和甜菜夜蛾幼虫的重要寄生性天敌[11-13]。本研究旨在用Bt棉饲喂斜纹夜蛾和甜菜夜蛾幼虫,观察斑痣悬茧蜂的寄主选择性以及子代蜂的发育表现,为评价Bt棉通过次靶标寄主对寄生蜂的影响提供依据。
1 材料和方法
1.1 供试植物及昆虫
供试棉花品种为‘泗棉3号’(常规棉)和‘国抗2号’(Bt棉),前者是后者转抗虫基因的亲本,其中Bt棉为转Cry1Ac基因,Bt毒素含量为(463±24.6)ng/g(ELISA酶联免疫技术测定)。供试斜纹夜蛾和甜菜夜蛾采自南京农业大学江浦农场大豆田,在室内分别用两种棉叶饲养,从孵化直至老熟化蛹。斑痣悬茧蜂从2008年9月采自南京农业大学江浦农场大豆田中的斜纹夜蛾幼虫育出,为产雌孤雌生殖品系,斑痣悬茧蜂属单寄生性内寄生蜂,单头寄生蜂幼虫在寄主幼虫体内完成发育到老熟后,在寄主幼虫腹部侧面咬一小洞爬出,在寄主幼虫附近结茧化蛹。在养虫室内(24℃±1℃、60%±10%RH和L∥D=14 h∥10 h)分别以甜菜夜蛾和斜纹夜蛾为寄主繁殖。非选择性试验所用蜂为4~6日龄(前期观察该日龄蜂产卵量最大)以及选择性试验中8~10日龄处女蜂(事前观察该日龄蜂搜索能力最强)。
1.2 试验方法
寄生选择:斜纹夜蛾幼虫寄生选择试验在纱网罩笼(木质包有透明纱网,长×宽×高=80 cm×80 cm×100 cm)中进行。笼中放置盆栽常规棉和Bt棉各1盆(高约40~50 cm,生长状况接近),每株分别接上取食常规棉和Bt棉叶的斜纹夜蛾3龄幼虫各10头,随机分布于各叶片,然后放入1头8~10日龄无寄生经验的寄生蜂,24 h后取出寄主,其中随机选择各处理幼虫5头单独称重(LAC114,Lavrock,常熟;精确到0.01 mg)。幼虫单独饲养每天观察记录寄生蜂幼虫爬出、结茧和羽化情况。重复15次。
甜菜夜蛾幼虫的寄生选择试验在养虫盒(直径8 cm,高9 cm)中进行,分别选取用常规棉和Bt棉叶饲喂至4龄的甜菜夜蛾幼虫各10头,盒底铺有1%的琼脂保湿,从塑料盒侧壁的小洞接入1头4~6日龄处女蜂,塑料盒内壁涂有10%的蜂蜜水供寄生蜂吸食;经8 h后移走寄生蜂,随机选择盒内寄主幼虫称重,并移入培养皿中用相应棉叶饲养,每日观察并记录寄生蜂的存活以及寄生蜂幼虫爬出寄主结茧化蛹情况等,并解剖死亡寄主幼虫检查是否被寄生。试验重复10次。
子代蜂发育表现:试验方法与选择试验中寄生选择试验甜菜夜蛾部分方法相同,记录寄主的存活以及寄生蜂幼虫爬出寄主结茧化蛹情况等,每日观察2次。羽化出的寄生蜂成虫单独置于试管内,不提供补充营养,观察其寿命。死亡的成蜂保存于75%的乙醇中随后测量体型大小。成蜂体型大小用后足胫节长度表示,在数码解剖镜(麦克奥迪SM2-143)下测量(精确到0.001 mm)。试验重复16次(斜纹夜蛾取食Bt棉叶)、17次(斜纹夜蛾取食常规棉叶)或10次(甜菜夜蛾)。
1.3 数据分析
寄生率为结茧的子代蜂数与解剖被寄生的幼虫数之和占全部供试幼虫的百分率;结茧率为结茧数占被寄生的寄主幼虫数的百分率;羽化率为羽化出蜂数占蛹数的百分率。寄生蜂子代的生长发育表现用发育历期和体型大小表示,发育历期为从雌蜂产卵到子代蜂羽化的时间,体型大小用后足胫节长度代表。用独立样本t测验比较取食2种棉叶的寄主幼虫体重差异,用非参数W ilcoxon秩和测验来比较寄生蜂的寄生、结茧和羽化的实际发生数量,用独立样本t测验比较子代蜂发育时间和体型大小(后足胫节长度);用非参数Kap lan-Meier方法估计寄主选择时间变量的生存函数,用log-rank测验比较生存函数的差异显著性。数据分析用R统计软件(R-2.7.1)[14]。
2 结果与分析
2.1 取食Bt棉对寄主幼虫体重的影响
取食常规棉叶的斜纹夜蛾寄主幼虫平均体重为(0.023 66±0.018 34)g,与取食Bt棉叶的斜纹夜蛾幼虫体重(0.021 65±0.013 71)g没有显著差异(独立样本t测验,p=0.208);而取食常规棉叶的甜菜夜蛾幼虫比取食Bt棉叶的重0.013 4 g,存在显著差异(独立样本 t测验,p=0.013)(图 1)。

图1 取食不同棉叶斜纹夜蛾3龄和甜菜夜蛾4龄幼虫的体重
2.2 斑痣悬茧蜂的寄生选择
在大罩笼中进行的寄生选择试验表明,斑痣悬茧蜂未表现出对以Bt棉叶为食的斜纹夜蛾幼虫的显著偏好(W ilcoxon秩和测验,p=0.242),寄生率为(32.51±3.53)%,仅比对取食常规棉叶幼虫的寄生率高4.61%(图2)。

图2 斑痣悬茧蜂对取食不同棉叶斜纹夜蛾幼虫的寄生选择
甜菜夜蛾独立塑料盒的非选择试验,斑痣悬茧蜂在相同时间内对相同寄主数的取食不同食料的甜菜夜蛾寄生选择数量同样没有表现出差异(p=0.889),但是同样表现出寄生取食Bt棉叶的甜菜夜蛾的数量比寄生取食常规棉叶的寄主幼虫多了2.46%(图3)。
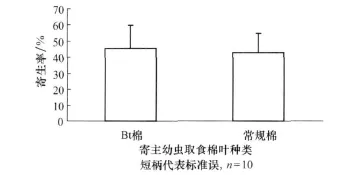
图3 斑痣悬茧蜂对取食不同棉叶甜菜夜蛾幼虫的寄生率
2.3 子代斑痣悬茧蜂的发育表现
在取食Bt棉叶的斜纹夜蛾幼虫体内发育的斑痣悬茧蜂子代,在发育时间、茧重、羽化蜂体型大小(后足胫节长度)等发育适合度相关特征方面,与寄生取食常规棉叶的寄主子代蜂没有显著差异(表1),子代蜂的存活时间也未表现出差异(图 4,logrank测验,p=0.898)。
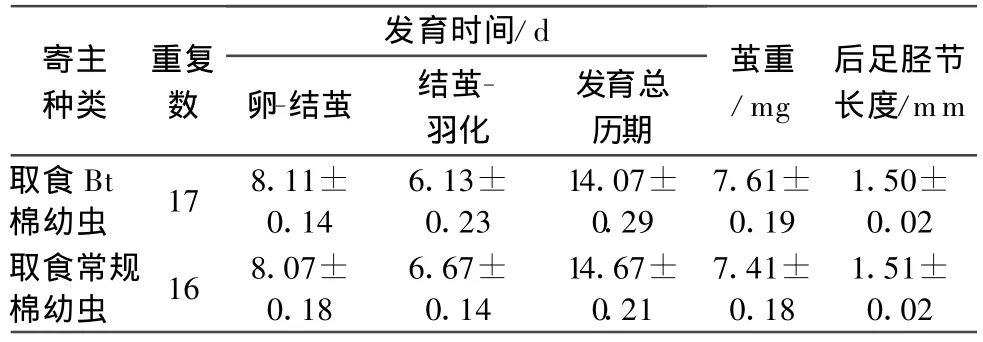
表1 在取食Bt棉和常规棉的斜纹夜蛾幼虫体内的子代蜂发育表现1)
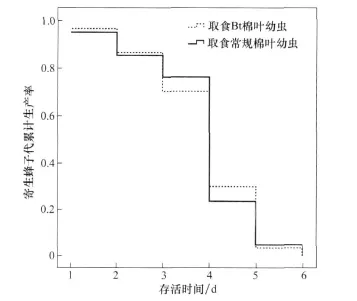
图4 在取食Bt棉和常规棉的斜纹夜蛾幼虫体内发育的子代蜂的Kaplan-Meier存活曲线
虽然取食两种不同棉叶的甜菜夜蛾体重不尽相同,但是斑痣悬茧蜂在取食两种不同棉叶寄主中发育出的子代蜂的后足胫节长度经过独立样本t测验比较发现差异不显著(p=0.476),其他相关的子代蜂发育适合度参数也表现出不明显的差异(表2)。
结茧率和羽化率也未在寄生斜纹夜蛾和甜菜夜蛾的子代蜂表现出显著差异,例如,在以Bt棉叶为食的斜纹夜蛾体内完成幼期发育的幼虫,结茧率为(61.88±36.56)%,羽化率为(63.10±37.37)%;而在以常规棉为食的斜纹夜蛾体内完成幼期发育的结茧率为(74.11±21.45)%,羽化率为(69.23±36.58)%(图 5)。
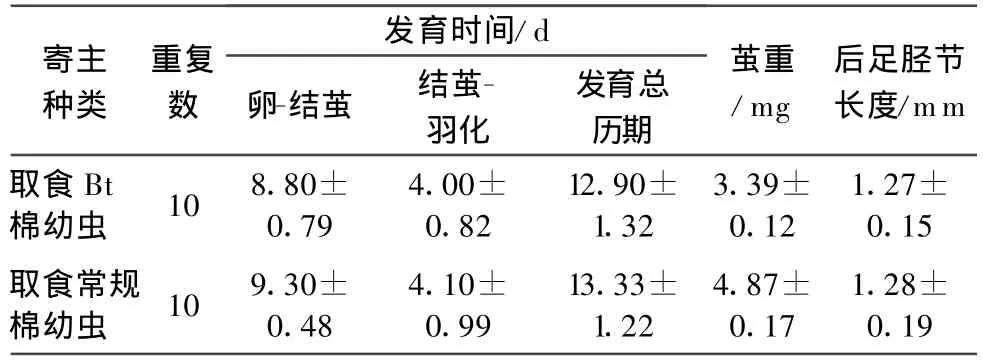
表2 寄生取食不同棉叶甜菜夜蛾幼虫的斑痣悬茧蜂子代生长发育表现1)
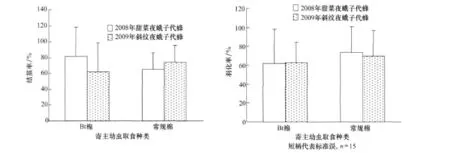
图5 取食Bt棉和常规棉的斜纹夜蛾和甜菜夜蛾幼虫体内发育的子代蜂结茧和羽化表现
3 讨论
寄生蜂在搜索寄主时常依赖于受害植物所发出的挥发性次生化合物[15-16],而害虫对植物的取食程度对这种功能的发挥有着至关重要的作用[17]。本试验中斑痣悬茧蜂在选择寄主时,对以Bt棉叶和常规棉叶为食的斜纹夜蛾幼虫的寄生未表现出显著的偏好,由于试验中未发现两种棉叶被取食程度存在差异,寄主幼虫体重也相近,因此推测可能是转Bt棉释放的挥发性物质与常规棉没有差异。阎凤鸣[18]对转Bt基因的棉花品种‘GK 12’与常规棉‘泗棉3号’的挥发性化学物质组成和含量的研究,没有发现两者存在明显差异。类似的结果也出现在其他研究中。例如,Orr等人的[19]研究表明,在Bt玉米田中套种小块常规玉米(由于欧洲玉米螟幼虫不能在转Bt玉米上存活),并人工接种欧洲玉米螟[Ostrinia nubila lis(Hübner)]卵,对玉米螟幼虫的总体寄生水平没有显著影响。在室内大罩笼中混种转Bt基因油菜和常规油菜,接种蚜虫后释放菜少脉蚜茧蜂(D iaeretiella rapae),寄生率在转基因与常规油菜间无显著差异[20]。
研究结果表明,斑痣悬茧蜂子代发育表现(历期、体型大小和存活率等)并未受寄主幼虫取食Bt棉叶的影响,也不受寄主幼虫体重差异的影响。由于斑痣悬茧蜂是容性寄生蜂,如果寄主幼虫被寄生后体内的营养物质足以支持其完成世代发育,则寄生蜂能够正常生长。因此我们推测取食Bt棉的非靶标寄主幼虫体内的营养物质足够支持寄生蜂生存所需要求,而可能存在的少量Bt毒素对斑痣悬茧蜂发育没有不良影响。在对其他寄生蜂的测定中也得出了类似的结论,例如,从Bt棉田和常规棉田采回的棉蚜茧蜂之间在羽化率、历期及体重等方面没有显著差异[2];寄生取食转GNA基因马铃薯的番茄夜蛾(Lacanobia oleracea)幼虫的毛角姬小蜂(Eulophus pennicornis)在后代体型大小、抱卵量和寿命等方面均表现正常[21],Bt棉花粉对寄生欧洲玉米螟的拟澳洲赤眼蜂(Trichogramma con fusuMViggiani)没有影响[22]。
转基因抗虫作物对非靶标生物的风险评估是其安全性评价的重要内容,其中对天敌昆虫的潜在影响评价对于实施害虫综合治理具有重要意义[4]。本试验研究在室内进行,未发现取食Bt棉的非靶标害虫(斜纹夜蛾和甜菜夜蛾幼虫)对其寄生蜂(斑痣悬茧蜂)的寄生选择以及子代发育有不良影响。但转基因抗虫作物对非靶标生物的潜在影响受到多种环境因素的影响,有必要进行大规模、长时期的野外试验,才能最终获得充分的依据进行科学评估[23]。
[1]W u KongMing,Lu Yanhui,Feng H ongqiang,et al.Supp ression of cotton bollw orMinMultiple crops in China in areaswith Bt toxin-containing cotton[J].Science,2008,321:1676-1678.
[2]崔金杰,雒珺瑜,王春义,等.转双价基因棉对棉田主要寄生性天敌生长发育的影响[J].棉花学报,2005,17(1):37-41.
[3]余月书,杨益众,任璐,等.转基因棉对棉田寄生物非亲和效应的研究[J].扬州大学学报(农业与生命科学版),2004,25(2):65-67.
[4]李保平,孟玲,万方浩.转基因抗虫植物对天敌昆虫的影响[J].中国生物防治,2002,18(3):97-105.
[5]Veiders R M,崔金杰,夏敬源,等.In fluence of transgenic cotton on the cotton aphid(Aphisgossy pii)and its tw oMajor eneMies in China[J].棉花学报,2002,14(3):175-179.
[6]Baur ME,Boethel D J.Effect of Bt-cotton ex pressing CrylA(c)on the su rvivaland fecundity of tw o hyMenopteran parasitoids(Braconidae,Encyrtidae)in the laboratory[J].Biological Con trol,2003,26(3):325-332.
[7]张晖,王长永,陈建群.基于食物链关系评价转Bt基因棉对寄生性天敌的非靶效应[J].农村生态环境,2003,19(2):55-57.
[8]董双林.转Bt基因棉及其抗虫性研究与利用进展[J].棉花学报,1998,10(2):57-63.
[9]夏敬源,崔金杰,常蕊芹.转基因抗虫棉对甜菜夜蛾的抗性研究[J].中国棉花,2000,27(9):10-11.
[10]张跃.2003年棉花斜纹夜蛾大发生的原因及防治对策[J].安徽农业科学2004,32(5):91.
[11]刘亚慧,李保平.斑痣悬茧蜂对甜菜夜蛾幼虫龄期的选择及其生长发育的研究[J].南京农业大学学报,2006,29(2):66-70.
[12]Liu Yahui,Li Baoping.Developmental in teractions betw een Spodoptera exigua(Noctuidae:Lepidoptera)and its uniparental endoparasitoid Meteorus pulchricornis(Braconidae:Hymenoptera)[J].BiologicalControl,2006,38:264-269.
[13]Liu Yahui,Meng Ling,Li Baoping.Effects of Helicoverpa a rmigera(Noctuidae,Lepidoptera)host stageson Some developmental parameters of the uniparental endoparasitoid Meteorus pu lchrico(Braconidae,Hymenoptera)[J].Bulletin of Entomological Research,2008,98:109-114.
[14]C raw ley MJ.The RBook[M].England:John Wiley&Sons,Ltd,2005.
[15]BerenbauMMR.Evolution of specialization in insect umbellifer[J].Annual Review of EntoMology,1990,35:319-343.
[16]严善春,张丹丹,迟德富.植物挥发性物质对昆虫作用的研究进展[J].应用生态学报,2003,14(2):310-313.
[17]Poppy G.GMcrops:environmental risks and non-target effects[J].T rend s in Plant Science,2000,5(1):1360-1385.
[18]阎凤鸣,许崇任,Bengtsson M,等.转Bt基因棉挥发性气味的化学成分及其对棉铃虫的电生理活性[J].昆虫学报,2000,45(4):425-429.
[19]Or r D B,Landis D A.Oviposition of European corn borer(Lepidoptera:Pyralidae)and impact of natural enemy populations in transgenic versus isogenic corn[J].Journal of Economic EntoMology,1997,90:905-909.
[20]Schuler T H.The iMpact of insect resistant GMcrops on population of natu ral eneMies[J].An tenna,2000,24(2):59-65.
[21]Bell H A,Fitches E C,Dow n RE,et al.The effect of snowdrop lectin(GNA)delivered via artificial diet and transgenic p lan ts on Eulophus pennicorn is(H ymenoptera:Eu lophidae),a parasitoid of the tomato moth Lacanobia oleracea(Lepidoptera:Noctuidae)[J].Jou rnal of Insect Phy siology,1999,45(11):983-991.
[22]耿金虎,沈佐锐,李正西,等.常规棉花粉和转CrvlAc+CpTI棉花粉对拟澳洲赤眼蜂繁殖和存活的影响[J].生态学报,2005,25(7):1575-1582.
[23]Romeis J,Bartsch D,Bigler F,etal.Assessment of risk of insect-resistan t transgenic crop s to nontarget arthropods[J].Nature Biotechnology,2008,26(2):203-208.
猜你喜欢
少儿科学周刊·少年版(2022年10期)2022-06-27
蜜蜂杂志(2019年12期)2019-06-16
西南国防医药(2018年12期)2019-01-02
河南农业科学(2018年6期)2018-01-18
上海农业学报(2017年6期)2018-01-05
女友·家园(2016年4期)2016-04-27
安徽农业科学(2015年10期)2015-12-17
科学大众·小诺贝尔(2015年5期)2015-05-04
癌变·畸变·突变(2015年3期)2015-02-27
糖尿病新世界(2015年6期)2015-01-04
