
植被性状与高原鼢鼠栖息地适合度的关系
2011-04-25 09:46葛庆征张卫国江小雷
草业科学 2011年5期
张 军,葛庆征,张卫国,江小雷
(兰州大学草地农业科技学院,甘肃 兰州 730020)
高原鼢鼠(Myospalaxbaileyi)栖息地的适合度取决于气候、植被、土壤、地形、天敌生物以及种内及种间关系等因素的综合效应[1-7]。在诸多因素的权衡中,由于植被条件直接决定着鼠类食物的数量和品质[8-9],同时也在很大程度上体现了其他环境因素的现实状况。因而,植被性状在鼠类栖息地适合度的构成要素中占有更高的权重,以至于可以凭借植被的某些性状指标直接对鼠类栖息地的适合度做出估测和评价[10]。植物多样性是指示植被演替进程的重要指标[11],它的变化不仅是植被现时状态的具体体现,而且也直接反映某些重要植物种在群落中的地位以及鼢鼠食物资源的丰富度[12]。因此,植物多样性对于判断高原鼢鼠栖息地的适合度具有重要的指示意义。目前,国内外有关植物多样性与啮齿动物栖息地相互关系的研究已有大量报道[13-14],但涉及高原鼢鼠的相关报道却非常有限,而且内容多以鼠丘植被的演替以及鼢鼠活动对植被多样性的干扰效应为聚焦点[15-18],而涉及多样性与栖息地适合度相互关系的研究鲜见报道。本研究以植被性状与高原鼢鼠栖息地适合度的相关关系为命题,以期探明二者相关关系及其作用机理,为有效开展高原鼢鼠的生态防治提供理论依据。
1 研究方法
1.1研究地概况 研究地设于甘南藏族自治州玛曲县以南玛(玛曲)-阿(阿万仓)公路约28 km处的公路南侧。地理坐标为33°51′ N,101°55′ E,海拔3 426 m。年平均降水量530.2 mm,年平均气温1.1℃,年日照时数2 583.9 h。地貌为宽谷滩地,土层深厚,土壤为亚高山草甸土。植被为亚高山草甸,属永久性天然放牧地,长期作冬季牧场利用,因过度放牧而呈中度退化态势。属典型高原鼢鼠适生地,鼠群密度8~25只/hm2,在自东向西的1 000 m区间内呈梯度递减之势,其南侧地势稍高处为高原鼠兔分布地,并有约20 m左右的共生区。
1.2试验设计 2005年4月,在研究地对南北向约100 m、东西向约500 m的区域实施围栏保护,设为试验区。在试验区内的东西方向上沿高原鼢鼠的密度梯度每隔50 m预设一个样地,并自东向西依次编号,样地面积50 m×50 m,共预设10个样地。4月15日数取并记录各样地历年鼠丘数,5月15日再次数取各样地当年形成的新鼠丘数,以同一样地“2×新鼠丘数+旧鼠丘数”代表该样地的鼠群密度[19]。在预设的10个样地中,确定5个存在显著密度差异的样地,形成5个密度梯度水平,并作为8月中旬进行植被分析的对应样地。由于鼠群密度与生境适合度高度正相关,5个密度梯度同时也代表着5个生境适合度水平。
1.3测定项目及方法 植被性状测定主要包括植被组成、盖度、草丛高度和地上生物量,各项测定皆按常规法[20]进行,样方面积0.5 m×0.5 m,各梯度样地重复5次。
1.4数据分析
1.4.1植物多样性 采用Shannon-Wiener指数(H′)表示,其计算公式如下:
H′=-∑(PilgPi);
Pi=Ni/N。
式中,Ni表示第i个物种在样方中的生物量,N表示样方的总生物量。
1.4.2物种丰富度(R) 样区内的植物种数。
1.4.3数据处理 数据皆由SPSS 13.0版统计软件分析整理,图表用Excel 2003制作。
2 结果与分析
2.1种群密度的梯度差异 新旧鼠丘数及其梯度变化如表1所示,在预设的10个样地中,鼠群密度基本按样地排序呈依次递减的梯度变化。其中,样地3和4及样地8和9的密度值相等,故可分别归入同一梯度级,样地1、2,样地5、6和样地7、8、9因差异相对较小也予以合并,最终将样地1、2,3、4,5、6,7、8、9,10确定为5个供试梯度样地。
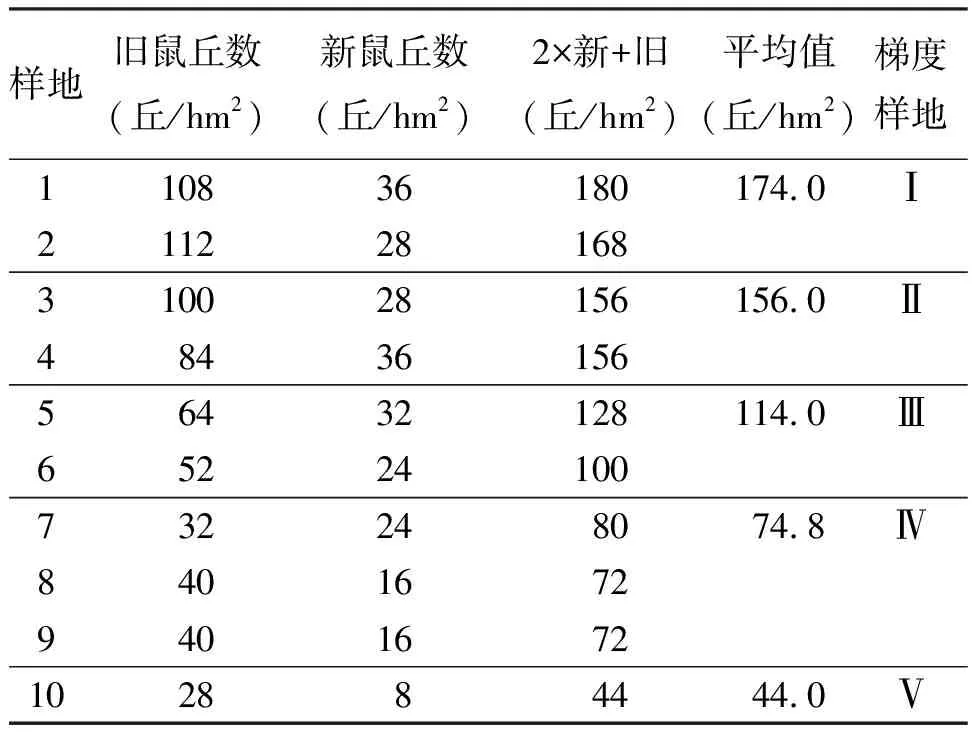
表1 各样地鼠群密度的梯度差异
2.2植被性状与鼠群密度的关系 在鼠群密度的5个梯度序列上,植被盖度依次为95.2%、89.5%、82.5%、70.1%和67.9%,呈随鼠群密度上升而平缓下降的变化趋势,与鼠群密度表现为显著负相关关系(P<0.05),决定系数为0.958 4(图1a)。其中,相邻梯度间除样地Ⅱ与样地Ⅲ差异显著(P<0.05)外,其他相邻梯度间差异均未达到显著级水平(P>0.05),但各相间梯度间则存在极显著差异(P<0.01)。相反,植被高度在各梯度样方的测定值依次为31.1、34.4、38.2、44.1和45.3 cm,除样地Ⅰ、Ⅱ间无显著性差异,Ⅱ、Ⅲ间差异极显著(P<0.01)外,其余相邻样地间的差异均达显著水平(P<0.05),整体上表现为随鼠群密度增加而上升的趋势(图1b),呈显著正相关关系,决定系数为0.970 7。地上生物量在各梯度样方的测定值依次为432.88、485.00、401.32、374.24和356.00 g/m2,各梯度样地间差异显著性与植被高度的表现基本一致,但生物量的最高值和最低值分别出现在样地Ⅳ和样地Ⅰ,生物量在鼠群密度梯度上的变化规律表现为非线性(图1c)。
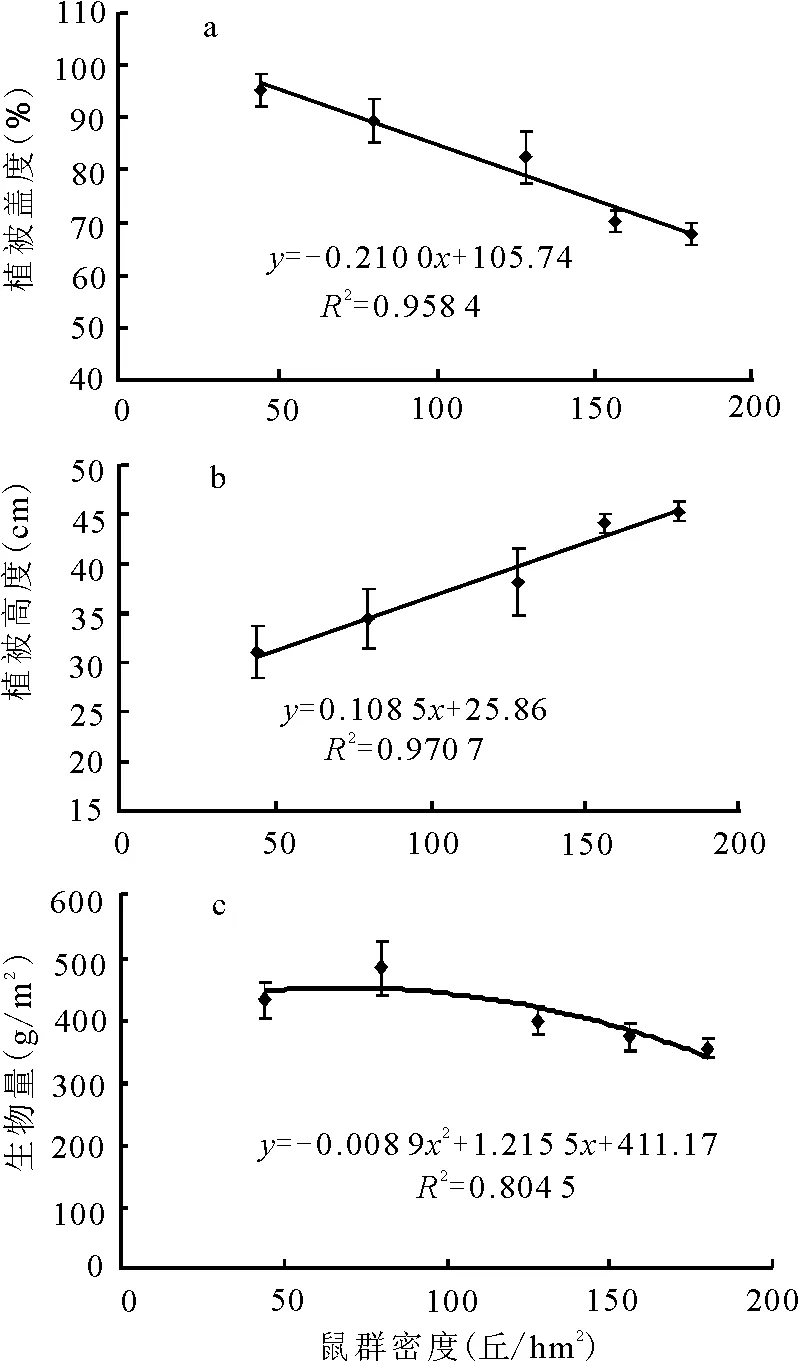
图1 植被性状与鼠群密度的关系
以上结果表明,在任何密度梯度上,鼠丘均存在着显著降低植物着生面积的负面作用,但只有在鼠群密度足够大时才会对植被地上生物量产生显著影响,而鼠丘对植被高度的正效应则在任何密度水平上均得到了显著体现。
2.3植物多样性与鼠群密度的关系 植物多样性和物种丰富度在鼠群密度的5个梯度序列上表现为由低到高、再由高到低的抛物线变化(图2),其峰值和最低值分别出现在样地Ⅲ和样地Ⅴ,在此密度区间,植物多样性的增势较大,样地Ⅴ与样地Ⅳ、样地Ⅳ与样地Ⅲ间的差异均极显著(P<0.01),而在高密度区间(样地Ⅲ至样地Ⅰ)则降势相对较缓,但相邻样地间的差异仍为显著(P<0.05)。
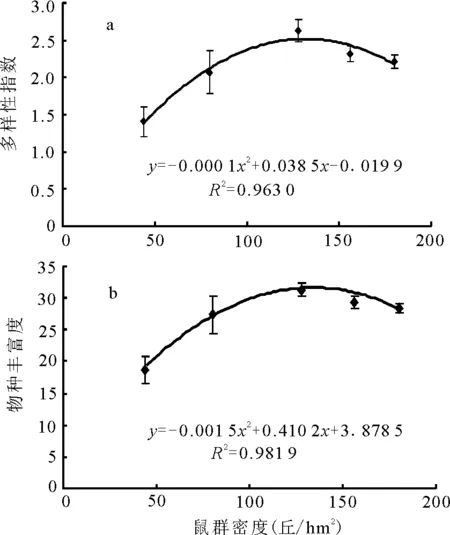
图2 植物多样性与鼠群密度的关系
这一结果表明,鼠丘对植物多样性的干扰效应符合“中度干扰理论”的基本原理。对于典型高寒草甸而言,适度的鼠群密度有利于植物多样性的维持,而过高或过低均可导致植物多样性的显著下降,且尤以高密度为甚。
3 讨论
植被是草原鼠类栖息环境最重要的组成部分,不仅以鼠类食物的主要来源影响其栖息条件的优劣[21],同时也以环境的感官效应[22]以及对其他相关生物的影响等途径对鼠类栖息地的适合度产生间接的影响。就高原鼢鼠而言,植被地表性状与鼠的活动,特别是掘土造丘活动密切相关[16]。鼠丘不仅以覆盖的方式影响植被盖度[23],而且也可通过改变土壤理化性质和土壤种子库性状的途径对植被的物种组成、草丛高度和生物量等产生间接的影响[1,23-26]。此外,高原鼢鼠持续不断的造丘活动可以在草地上形成大量异质性斑块,从而为植物多样性的维持和提高创造了必要的条件[15,27]。显然,鼠群密度越高,植被所受到的扰动也就越强烈。因而,植被性状的变化在很大程度上反映了鼠群密度的变化,二者之间存在着一定的对应关系。在本研究中,植被盖度与鼠群密度之间存在着极为显著的负相关性,这显然是鼠群密度与鼠丘密度的高度正相关性所导致的结果。因而,在高原鼢鼠的典型分布地,以植被盖度作为推测鼠群密度或生境适合度的指标不仅在理论上是可行的,而且在实践中也具有重要的参考价值。草丛高度尽管与鼠群密度存在着显著的正相关性,但由于影响植被高度的不确定因素较多,仅凭植被高度作为推测鼠群密度或生境适合度的指标是不可靠的,极有可能导致错误的判断。地上生物量的性质与植被高度类似,同样会受到许多不确定因素的干扰而失去其应有的稳定性,但对鼠群密度或生境条件的指示意义却明显高于草丛高度,当然,若能以杂类草在地上生物量中所占比重作为推测指标,无疑将会提高推测结果的准确性。
植物多样性与高原鼢鼠的造丘活动密切相关[1,15-16]。由于不同时间形成的鼠丘处于不同的演替阶段,并较为均匀地分布于整个次生演替序列的各个阶段,这就使环境的异质性在鼠丘斑块水平上得到最大化的发展,进而使对立地条件有着不同要求的物种都能获得生存的机会。由植物多样性的形成机制不难看出,在健康的草地系统中,高原鼢鼠种群密度由于系统调节机制和种群自身调节机制的作用而始终保持在一个正常的波动范围内,在此阈值内,植物多样性与鼠群密度的对应关系也相应地处于一种相对稳定的状态,不易受到其他因素的强烈干扰。此外,植物多样性对高原鼢鼠食物的丰富度具有重要的指示意义,在正常的范围内,植物多样性与高原鼢鼠的食物丰富度呈显著正相关关系[16],这也就是说,较高的植物多样性在一定程度上代表着较高的生境适合度。因此,以植物多样性指数去间接地推知高原鼢鼠的生境适合度具有较高的可信度。然而,当系统受到某种因素的强烈扰动(如过度放牧地、弃耕地等)而发生系统功能紊乱时,高原鼢鼠的种群密度就会出现背离正常消长规律的异常变化,此时,依靠植物多样性作为高原鼢鼠生境适合度的估测参数,其准确性就会受到一定的影响。在本研究所设定的密度范围内,植物多样性的变化规律与“中度干扰理论”相吻合,即中等密度水平有利于植物多样性的最大化,而高密度和低密度水平均可导致植物多样性的梯度性下降。因此,植物多样性指标在估测高原鼢鼠生境适合度的实践中具有较高的参考价值。
4 结论
1)高原鼢鼠的生境适合度与植被性状之间存在着显著的相关性,因此,植被性状的相关指标不仅可以用于推测高原鼢鼠的生境适合度,并可据此对其种群密度的发展趋势做出预测。
2)植被盖度与高原鼢鼠种群密度表现为极显著线性相关,因而,在高原鼢鼠的典型分布地,依据植被盖度推测高原鼢鼠的种群密度,进而对生境适合度作出评价,具有拟合度高、简便易行的特点。
3)植物多样性是高原鼢鼠种群密度、活动频度及食物丰富度的重要表征,在生境适合度评价、鼠群消长趋势预测等领域具有相关度高、稳定性好的特点。
[1] Zhang Y M,Liu J K.Effects of plateau zokors (Myospalaxfontanierii) on plant community and soil in an alpine meadow[J].Journal of Mammalogy,2003,84:400-407.
[2] 张堰铭,樊乃昌,王权业.高原鼢鼠种群密度环境因子的关系[J].高原生物学集刊,1999,14:101-109.
[3] 张堰铭.高原鼢鼠对高寒草甸群落特征及演替的影响[J].动物学研究,1999,20(6):435-440.
[4] 韩天虎,张卫国,丁连生.高原鼢鼠栖息地的植被特征[J].草业学报,1999,8(2):43-49.
[5] 魏万红,王权业,周文扬,等.捕食干扰对Myospalaxbaileyi种群繁殖力的影响[J].生态学报,1998,18(2):176-180.
[6] 张卫国,江小雷.高原鼢鼠与高原鼠兔共栖关系的研究[J].草业学报,1998,7(2):43-47.
[7] 黄倩,花立民,曹慧,等.甘肃草原鼠害区划研究[J].草业科学,2009,26(02):91-99.
[8] Torres-Contreras H,Bozinovic F.Food selection in an herbivorous rodent:Balancing nutrition with thermoregulation[J].Ecology,1997,78(7):2230-2237.
[9] Turchin P,Batzli G O.Availability of food and the population dynamics of arvicoline rodents[J].Ecology,2001,82(6):1521-1534.
[10] Olson G S,Glenn E M,Anthony R G,etal.Modeling demographic performance of northern spotted owls relative to forest habitat in Oregon[J].Journal of Wildlife Management,2004,68(4):1039-1053.
[11] 贺金生,陈伟烈.陆地植物群落物种多样性的梯度变化特征[J].生态学报,1997,17(1):91-99.
[12] Hambäck P A,Beckerman A P.Herbivory and plant resource competition:a review of two interacting interactions[J].Oikos,2003,101:26-37.
[13] 刘伟,周立,王溪.不同放牧强度对植物及啮齿动物作用的研究[J].生态学报,1999,19(3):378-382.
[14] Smith A T,Foggin J M.The plateau pika (Ochotonacurzoniae) is a keystone species for biodiversity on the Tibetan Plateau[J].Animal Conservation,1999,2:235-240.
[15] 江小雷,张卫国,杨振宇.不同演替阶段鼢鼠土丘群落植物多样性变化研究[J].应用生态学报,2004,15(5):814-818.
[16] 张卫国,江小雷,王树茂,等.鼢鼠的造丘活动及不同休牧方式对草地植被生产力的影响[J].西北植物学报,2004,24(10):1882-1887.
[17] 张堰铭,刘季科.地下鼠生物学特征及其在生态系统中的作用[J].兽类学报,2002,22(2):144-154.
[18] 周雪荣,郭正刚,郭兴华.高原鼠兔和高原鼢鼠在高寒草甸中的作用[J].草业科学,2010,27(5):38-44.
[19] 何俊龄,张金沙,杨莹博,等.高原鼢鼠土丘空间格局及主要特征研究[J].草业学报,2006,15(1):107-102.
[20] 任继周.草业科学研究方法[M].北京:中国农业出版社,1998:11-15.
[21] Batzli G O,Lesientre C.The influence of high quality food on habitat use by arctic microtine rodents[J].Oikos,1991,60:299-306.
[22] Gebresilassie W,Bekele A,Belay G,etal.Microhabitat choice and diet of rodents in Maynugus irrigation field,northern Ethiopia[J].African Journal of Ecology,2004,42:315-321.
[23] Reichman O J,Smith S C.Impact of pocket gopher burrows on overlying vegetation[J].Journal of Mammalogy,1985,66:720-725.
[24] Mielke H W.Mound building by pocket gophers (Geomyidae):their impact on soils and vegetation in North America[J].Journal of Biogeography,1977,4(2):171-180.
[25] Rebollo S,Pérez-Camacho L,Valencia J,etal.Vole mound effects and disturbance rate in a mediterranean plant community under different grazing and irrigation regimes[J].Plant Ecology,2003,169(2):227-243.
[26] 孙飞达,龙瑞军,蒋文兰,等.三江源区不同鼠洞密度下高寒草甸植物群落生物量和土壤容重特性研究[J].草业学报,2008,17(5):111-116.
[27] Guo Q F.Microhabitat differentiation in Chihuahuan Desert plant communities[J].Plant Ecology,1998,139(1):71-80.
猜你喜欢
林业资源管理(2022年6期)2023-01-18
Zoological Research(2022年3期)2022-06-07
南方农业(2021年30期)2021-12-25
现代园艺(2021年23期)2021-12-01
落叶果树(2021年6期)2021-02-12
林业勘查设计(2020年1期)2021-01-18
新农业(2020年18期)2021-01-07
兽医导刊(2020年3期)2020-12-30
防护林科技(2019年7期)2019-03-25
体育科技(2016年2期)2016-02-28
