
初始密度对广聚萤叶甲种群扩张的影响
2011-04-04 03:19:50郑兴汶周忠实郭建英万方浩陈红松王建国
环境昆虫学报 2011年1期
郑兴汶,周忠实,郭建英,万方浩*,陈红松,王建国
(1.江西农业大学农学院植物保护系,南昌 330045;2.中国农业科学院植物保护研究所植物病虫害生物学国家重点实验室,北京 100193)
广聚萤叶甲Ophraella communa属鞘翅目、叶甲科Chrysomelidae、萤叶甲亚科Galercinae,北美种,是入侵我国的世界性恶性杂草豚草Ambrosia artemisiifolia的一种重要天敌,以幼虫和成虫取食豚草叶片,可有效抑制豚草的扩散和蔓延。其生物学特性、寄主专一性、扩散能力及冷藏运输技术等前人做了大量的研究工作 (Zwǒlfer and Harris,1971;Palmer and Goeden,1991;Teshler et al.,2004)。Dernovici等评估了广聚萤叶甲对向日葵的安全性和潜在风险,认为其对野外向日葵的风险极低,是豚草的一种有效、安全的天敌 (Dernovici et al.,2006)。目前,该叶甲在美国、墨西哥、加拿大、日本、韩国等已被广泛研究和应用 (Palmer and Goeden,1991;Sohn et al.,2002;Tamura et al.,2004;Kiss,2007)。在加拿大,广聚萤叶甲已成功用于控制蔬菜地发生的豚草 (Teshler et al.,2004)。研究表明,该叶甲在室内适宜的温湿度下种群数量将迅速增长,表现出巨大的繁殖力(Zhou et al.,2010a;Zhou et al.,2010b)。本文通过在半自然的状态下考察不同初始密度对广聚萤叶甲种群发展的影响,以期为其大棚扩大饲养提供依据,并明确其在野外适宜气候条件下种群扩增潜力。
1 材料与方法
1.1 供试昆虫
广聚萤叶甲来自湖南省农业科学院植物保护研究所豚草天敌饲养室。采集初羽化成虫,置于透明塑料养虫盒 (19 cm ×12 cm ×6 cm)内备用。
1.2 试验设计和方法
试验地内设20个小区,每小区面积6.0 m×1.5 m,各小区内于2008年4月20日种植15 cm高的豚草幼苗30株 (2行×15列),行距×株距为60 cm×40 cm。四周立6根2 m高的竹竿,然后围40目的筛网。按接入广聚萤叶甲成虫数量的不同设置4个处理 (即3、5、10和20对/小区),每处理5个重复。试验于2008年7月19日按上述密度释放叶甲于各试验小区。释放后第13 d开始进行第1次调查,之后每隔12 d调查1次,每次均随机抽取各小区的10株豚草作为调查对象,调查、记录每株豚草上叶甲卵、幼虫、蛹、成虫的数量。
1.4 数据统计
将实验数据进行正态和方差性检验,必要时则将数据进行反正弦平方根或对数转换。不同处理之间的实验数据经单因素多重比较进行差异显著性分析 (P≤0.05) (SAS Institute,2004),通过最小差异法 (LSD法)比较处理间平均数的差异程度。
2 结果与分析
2.1 广聚萤叶甲卵的数量动态
广聚萤叶甲卵的数量在各处理间存在显著性差异 (8月1日:F3,16=105.90,P<0.0001;8月12日:F3,16=449.45,P<0.0001;8月 23日:F3,16=60.17,P<0.0001;图1)。在试验初期,各处理的卵量较低,均在50粒/株以下,其中以释放20对成虫的卵量最多,为45.45粒/株。随着时间推移,除高密度处理 (20对)外,其余处理的卵量随着时间而不断上升。相对于其它处理,高密度处理的卵数量高峰期出现较早,之后迅速降低,至8月23日高密度处理的卵量明显低于其它处理。

图1 广聚萤叶甲卵数量的动态变化Fig.1 Dynamics of O.communa eggs
2.2 广聚萤叶甲幼虫数量动态
各处理间广聚萤叶甲幼虫数量差异较大 (8月1 日:F3,16=96.85,P<0.0001;8 月 12 日:F3,16=121.34,P<0.0001;8 月 23 日:F3,16=10.43,P=0.0005;图2)。高密度处理 (20对)的幼虫种群上升速度快,8月1日调查,其密度高达47.0头/株,之后种群逐渐减少。释放10对成虫的小区,出现了明显的起伏,即第1次调查发现幼虫种群数量仅次于高密度处理区,随后急剧下降至最低,而最后1次调查,其种群数量上升至最高。两种低初始密度的小区幼虫种群数量一直维持在较低的水平。

图2 广聚萤叶甲幼虫数量的动态变化Fig.2 Dynamics of O.communa larvae
2.3 广聚萤叶甲蛹的数量动态
各时期处理间蛹的数量差异显著 (8月1日:F3,16=11.10, P=0.0003;8 月 12 日:F3,16=21.31,P<0.0001;8 月 23 日:F3,16=36.90,P<0.0001;图3)。在实验初期,各处理的蛹数量都较少,均在2头/株以下。随时间的推移各处理蛹的数量逐渐增多,其中以高密度处理 (20对)蛹的种群数量最多,在8月12日和8月23日的调查中蛹量分别可达8.08头/株和17.00头/株。在8月1日和8月12日的调查中,低密度处理 (3对)蛹的数量显著低于其它处理,随后种群迅速增长,至8月23日其种群数量仅次于高密度处理,为6.80头/株。
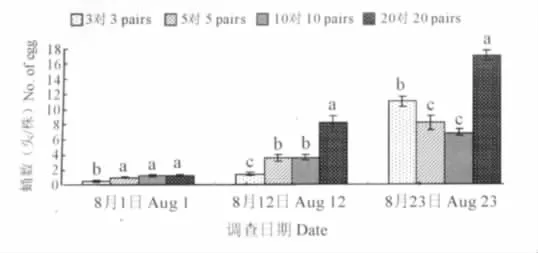
图3 广聚萤叶甲蛹数量的动态变化Fig.3 Dynamics of O.communa pupae
2.4 广聚萤叶甲成虫的数量动态
各处理的成虫数量存在着显著性差异 (8月1日:F3,16=20.68,P<0.0001;8 月12 日:F3,16=130.89,P<0.0001;8 月 23 日:F3,16=41.47,P<0.0001;图4)。在实验初期,各处理的成虫数量都较低,均在2头/株以下。随实验的持续进行各处理成虫的数量逐渐增多,其中以高密度处理(20对)的成虫种群数量增长最为迅速,至8月12日其成虫数量便可达11.4头/株。而低密度处理 (3对)的成虫种群数量在8月12日的调查中还处于较低水平,仅为1.68头/株,而后种群数量有较快增长,8月23日成虫种群数量超过释放5对的处理。从总体上看,除低密度处理,其余的各处理在8月12日之后成虫种群数量变化不明显。

图4 广聚萤叶甲成虫数量的动态变化Fig.4 Dynamics of O.communa adults
3 结论与讨论
昆虫在一定时期内,其种群迅速增长需要一定的起始虫量 (即虫口基数)。本试验结果表明,不同成虫初始密度对广聚萤叶甲的种群扩增具有显著的影响,在高初始密度下其种群短期内即迅速增长,成虫释放20天后,广聚萤叶甲各虫态数量均明显增加。在高初始密度小区中,后期叶甲卵和幼虫数量显著降低,尤其卵明显低于其它处理,卵和幼虫是种群扩张的基础,由此推断8月23日之后,高密度处理 (20对)小区的广聚萤叶甲种群数量会急速降低。然而,最后一次调查发现,高密度处理小区内的豚草已经被取食殆尽,因此食源不足可能是导致种群下降的原因之一。广聚萤叶甲成虫寿命长,具有多次交配、持续产卵的习性 (孟玲和李保平,2006),其世代重叠显著,在野外笼罩的情况下,在8月份卵到成虫的历期为19 d(孟玲等,2007)。由于世代重叠,导致了高密处理蛹和成虫数量在试验的中后期一直保持较大密度。
目前广聚萤叶甲主要分布在我国南方的一些省的豚草发生区,据推测其也可在华北和东北一带定居 (孟玲等,2007;hiyakeand Moriya,2005),具有广泛的应用前景 (Zhou et al.,2010b;Emura,1999)。在我国南方一些省份,广聚萤叶甲已被作为一种豚草的生物防治制剂进行推广和应用,取得了良好的控制效果 (周忠实等,2009)。经研究,认为大棚规模饲养是解决该叶甲的工厂化生产之一,根据我们的试验结果,推荐0.4~0.7头/株成虫作为大棚饲养的起始虫数为宜,如此可确保30 d内取得较大的种群数量。
References)
Dernovici SA,Teshler MP,Watson AK,2006.Is sunflower(Helianthus annuus)at risk to damage from Ophraella communa,a natural enemy of common ragweed(Ambrosia artemisiifolia)?Biocontrol Science and Technology,16(7):669-686.
Emura K,1999.The ragweed beetle Ophraella communa LeSage(Coleoptera:Chrysomelidae)which injures harmful exotic plants.
Shokubutsu boeki(Plant protection),53(4):138-141.
Kiss L,2007.Why is biocontrol of common ragweed,the most allergenic weed in Eastern Europe,still only a hope?In:Vincent C,Goettel MS,Lazarovits G,eds.Biological Control:A Global Perspective.CAB International,Oxfordshire,UK,80 -91.
Meng L,Li BP,2006.Mating and oviposition behaviors of exotic leafbeetle.Chinese Bulletin of Entomology,43(6):806-809.[孟玲,李保平,2006.豚草条纹萤叶甲的交配和产卵行为.昆虫知识,43(6):806-809]
Meng L,Xu J,Li HB,2007.Dispersal and bionomics of the Alien Ophraella communa in China Mainland.Chinese Journal of Biological Control,23(1):5-10.[孟玲,徐军,李海波,2007.外来广聚萤叶甲在我国的扩散及生活史特征.中国生物防治,23(1):5 -10]
Palmer WA,Goeden RD,1991.The host range of Ophraella communa Lesage(Coleoptera:Chrysomelidae).Coleopterists Bulletin,45(2):115-120.
SAS Institute,2004.SAS User's?Guide:Statistics,SAS Institute,Cary,NC.
Shiyake S,Moriya S,2005.Expansion of Ophraella communa Lesage in East Asia.Insect and Nature,40(4):11 -131.
Sohn JC,An SL,Li JE,Park KT,2002.Notes on exotic species,Ophraella communa LeSage(Coleoptera:Chrysomelidae)in Korea.Korean Journal of Applied Entomology,41(2):145-150.
Tamura Y,Hattori M,Konno K,Kono Y,Honda H,Ono H,YoshidaM,2004.Triterpenoid and caffeic acid derivatives in the leaves of ragweed,Ambrosia artemisiifolia L.(Asterales:Asteraceae),as feeding stimulants of Ophraella communa LeSage(Coleoptera:Chrysomelidae).Chemoecology,14(2):113-118.
Teshler MP,Demovici SA,DiTommaso A,Coderre D,Watson AK,2004.A novel device for the collection,storage,transport,and delivery of beneficial insects,and its application to Ophraella communa(Coleoptera:Chrysomelidae).Biocontrol Science and Technology,14(4):347-357.
Zhou ZS,Guo JY,Chen HS,Wan FH,2010a.Effect of humidity on the development and fecundity of Ophraella communa(Coleoptera:Chrysomelidae).BioControl,55(2):313 -319.
Zhou ZS,Guo JY,Chen HS,Wan FH,2010b.Effects of temperature on survival,development,longevity,and fecundity of Ophraella communa(Coleoptera:Chrysomelidae),a potential biological control agent against Ambrosia artemisiifolia(Asterales:Asteraceae).Environmental Entomology,39(4):1021-1027.
Zhou ZS,Guo JY,Wan FH,2009.Biocontrol of Ambrosia artemisiifolia.In:Wan FH,Guo JY,Zhang F.Research on Biological Invasions in Chian.Beijing:Science Press,246-251.[周忠实,万方浩,郭建英,2009.普通豚草的生物防治.见:万方浩,郭建英,张峰.中国生物入侵研究.北京:科学出版社,246-251]
Zwǒlfer H,Harris P,1971.Host specificity determination of insects for biological control of weeds.Annual Review of Entomology,16:159-178.
猜你喜欢
草食家畜(2022年5期)2023-01-02 02:53:26
资源信息与工程(2021年5期)2022-01-15 05:37:28
环境昆虫学报(2020年3期)2020-07-07 01:35:48
四川地质学报(2020年2期)2020-05-31 06:53:12
现代农村科技(2018年8期)2018-08-23 12:07:50
金色少年(奇趣科普)(2017年6期)2017-07-25 09:21:14
新农业(2016年19期)2016-08-16 12:00:18
中国房地产业(2016年8期)2016-03-01 01:26:17
湖南农业科学(2015年5期)2015-02-27 14:33:51
湖南农业(2014年7期)2014-04-30 08:25:08
