
扎龙湿地游禽类数量变动及原因分析1)
2011-03-29 08:06:44沙剑斌李枫苏立英逄士良高中艳
东北林业大学学报 2011年8期
沙剑斌 李枫 苏立英 逄士良 高中艳
(东北林业大学,哈尔滨,150040)(国际鹤类基金会)(扎龙国家级自然保护区)
鸟类群落对直接的和间接的环境变化异常敏感[1],它的变化可以直接反映出环境的变化,可作为监测评价环境质量的因子。湿地的水环境变化能直接或间接地影响在此栖息的水鸟群落动态[2]。而在湿地中繁殖、觅食的游禽类对湿地中水位的变化最为敏感[3]。研究湿地游禽类鸟类群落结构的变化,并分析引起其变化的原因,有助于对湿地环境的变动及其质量进行客观的评价,为有效地保护湿地生态系统的完整性提供科学依据[4]。
20 世纪80年代及90年代初期对扎龙鸟类的区系、群落特征等有过一些研究报道,共记录265种,其中游禽类有䴙䴘目鸟类5种,雁形目鸟类30种,秧鸡科鸟类2种[5-6],[7]19,[8]。近20 a来,人为活动(筑坝挖渠,捕鱼等)的不断增加,导致了湿地内原始景观发生了变化,湿地面积减少,湿地破碎化严重,人类活动加剧[9-10]。21世纪初李枫、邹红菲等人研究了扎龙湿地荒火、人工补水等对丹顶鹤营巢、生境选择的影响[11-14]。但是针对扎龙湿地环境变化对完全依赖湿地水环境觅食、筑巢、夜宿的游禽类影响的研究尚无相关报告。本研究通过监测扎龙湿地内游禽类的种类组成和数量变动,分析其群落动态,了解扎龙湿地环境变化对游禽类群落的影响,为保护区的湿地恢复提供有力的科学依据。
1 研究区概况
扎龙湿地位于东北平原北部的乌裕尔河和双阳河流域下游,嫩江平原西部,位于东经123°51.5'~124°37.5',北纬46°48'~47°31.5',总面积21万hm2。该区属于中温带大陆性季风气候,年平均气温3.5℃,年均风速3.5 m/s,多年平均降水量419 mm,多年平均蒸发量729.6 mm,年均降水量远远小于蒸发量。湿地内地势低洼平坦,海拔140~146 m,地面平均比降1/6 000。湿地内分布众多泡沼,地下水位高,排水不畅,土壤盐渍化比较普遍[15]。发源于小兴安岭西侧的乌裕尔河,源头海拔418 m,全长576 km,流域面积1 508 km2,年平均径流量为6.18亿m3。河流经长途流至本区后比降大减,冲击作用增强,漫溢于闭流区洼地,形成该区面积广阔的芦苇沼泽湿地,河道则隐没于湖沼芦苇沼泽中。湿地内共有高等植物525多种,鸟类265种,哺乳动物20种,两栖类4种,鱼类53种,其中国家重点保护鸟类35种,尤其以鹤类资源保护著称[16]。
2 研究方法
由于雁形目、䴙䴘目及鹤形目秧鸡科的骨顶鸡和黑水鸡这些鸟类的生态型相似,本研究特将雁形目、䴙䴘目及骨顶鸡和黑水鸡等这些鸟类一并称为游禽类。
2.1 调查范围
在扎龙湿地的西部设立调查区域(图1b和图1c)。该区域位于扎龙湿地西侧,这个区域涵盖2个较大的明水泡沼,大面积的芦苇沼泽,部分草甸,草甸草原,一个较大的(2 000多人口)和一个中型(500多人口)的村庄和少部分农田。根据湿地类型特点设置了3条调查样线,分布于保护区局址附近的仙鹤湖、龙湖及中部引嫩(中引)八支干沟渠上。调查区域面积约为1 668.5 hm2,约占扎龙湿地面积的0.8%,调查范围与1983年的夏秋季调查区域完全重叠。

图1 扎龙湿地地理位置
样线Ⅰ:由扎龙保护区旅游区放鹤地出发,向北经其饲养场,沿仙鹤湖东侧环湖高约1.0~1.5 m的坝棱子向北,经仙鹤湖北边天然小浅水湖沼(水深5~10 cm)、向西至中引八支干交界处。仙鹤湖是扎龙湿地内较大的湖沼之一,总面积约为180 hm2。在调查样线上向西在坝和明水湖面间有50~150 m宽的芦苇和蒲草为优势的挺水植物组成的芦苇沼泽带,另一侧生境是典型的芦苇沼泽生境。调查样线长约4.2 km,覆盖芦苇沼泽面积821.8 hm2,明水面面积478.2 hm2。
样线Ⅱ:吐木克村出发经小片农田,沿龙湖堤坝围绕龙湖一周。龙湖为扎龙湿地内较大的湖沼之一,总面积约为403 hm2。样线内侧与龙湖明水面间有200~350 m宽的芦苇沼泽带,样线外侧为大面积芦苇沼泽区。调查样线长约8.1 km,其覆盖了两种生境类型:农田、芦苇沼泽、明水,覆盖芦苇沼泽面积60 hm2,农田面积约13.6 hm2,明水面面积为403 hm2。
样线Ⅲ:自中引八支干与扎龙乡至齐市公路交界处沿八支干水渠至与翁海排干交叉处,堤坝西北侧生境为草甸和草甸草原,东南侧生境主要为芦苇沼泽;两坝之间为引水渠、坝二侧为护坝的灌丛林、防护林。调查样线长约4 km,覆盖草甸和草甸草原面积600 hm2,引水渠面积为7 hm2,灌木丛、防护林面积为3.5 hm2。
2.2 调查频率及方法
2.2.1 野外调查时间及频率
调查时间自2006年4月份开始至2009年8月份结束。将每年的4—5月份为春季迁徙季节调查期,一般情况1次/周;进入迁徙高峰期,监测频率相应增加至2次/周。6—7月份为夏季繁殖季节调查期,6月1—10日1次/周;6月11日—7月30日2周1次。8—11月为秋季迁徙季节调查期,8月1日—10月15日1次/周,10月16日—11月15日2周1次。12月至次年3月底为冬季调查期,1次/月。
2.2.2 调查方法
在设定的样线上,在晴朗、3级风以下的天气进行调查,每天调查时间在清晨或傍晚,步行速度控制在2~3 km/h。借助8~10倍双筒望远镜和20~60倍单筒望远镜直接记录位于样线前方及两侧的游禽类,其中包括飞过样地的个体。繁殖期调查时听到或看到一只雄鸟记做一对,在没有见到雄鸟的情况下,见到一只雌鸟,一窝卵或雏也视为一对[17]。
对调查区域内一些较大的明水面,如:仙鹤湖及其北边水泡、龙湖等游禽类集中分布、数量大的区域进行定点观察,借助8~10倍双筒望远镜和20~60倍单筒望远镜采用直数法和目测估算法,直接记录鸟类种类、数量。当鸟类数量较少时,直接计数;若鸟类数量较多时,则采用目测估算法[18]。
2.3 数据分析
为了便于比较每个时期的调查结果,我们用遇见率和频度作为鸟类数量指数,以减少由于在调查时间上的差异所造成的误差和随机误差。每次将一条样线调查作为一个样本。
遇见率:Ri,j=Ni/Hj。
式中,R(i,j)为j时间段内i种游禽鸟类的遇见率(只·h-1);Ni为第i种记录到的个体数量(i=1,2,3……24);Hj为j时间段总调查时间[7]17-18。
频度:Oi,j=si/Sj。
式中,O(i,j)为j时间段内第i种游禽鸟类的出现频(%)度;si为i种在j时间段调查的出现次数;Sj为j时间段总调查次数。
将本调查的同期(夏秋季)数据与1983年的(夏秋季)调查数据进行比较,以分析扎龙游禽类鸟类群落的变化。仍采用1983年扎龙鸟类资源调查中的数量级评定方法,将物种的优势度划分为6个数量等级[7]17-18。
Ⅰ级:数量极多的种,平均每小时可遇见10只以上的鸟类;
Ⅱ级:数量最多的种,平均每小时遇见1~9.999只的鸟类;Ⅲ级:优势种,平均每小时遇见0.142 9~0.999的鸟类;Ⅳ级;普通种,平均每小时遇见0.016 7~0.142 8只的鸟类;
Ⅴ级:数量很少的种,平均每小时遇见0.01~0.016 6只的鸟类;
Ⅵ级:数量极少的种,平均每小时遇见数量低于0.01只的鸟类。
3 结果与分析
4 a间总计野外调查时间为670 h。2006年调查时间为89 h、2007年为263 h、2008年为267 h、2009年为47 h。由于人力、天气等缘故,夏冬季节实际监测频率低于设计值。2006年和2009年的数据有些月份有间断。
3.1 2006—2009年间扎龙湿地内游禽类种类组成
在2006年4月至2009年8月调查中所记录所有鸟类种类为14目42科171种,其中游禽类种类为3目3科24种。游禽类占整个扎龙湿地鸟类群落的14.04%。以优势度来看,4 a所见游禽类中无I级、Ⅴ级和VI级的物种;II级为5种,III级共14种,Ⅳ级5种(表1)。

表1 扎龙保护区2006—2009年游禽组成及组成变化
与扎龙保护区建区早期的调查记录的37种相比[17]19-21,其中有13种游禽在此次调查中未见,即减少了35%的物种。
3.2 2006年—2009年扎龙湿地游禽类的数量变动
3.2.1 扎龙湿地游禽类的数量和出现的频率
遇见率说明物种的种群数量,而出现的频率则表明物种在时间上的分布状态。总结了每年各种类遇见率和频度的月平均值,结果发现整个调查期间只有3个物种小䴙䴘、风头䴙䴘、和绿头鸭4 a的月平均遇见率>1只·h-1;另有5个物种在个别年份的月平均遇见率>1只·h-1;大多数物种的月平均遇见率非常低。由此可见扎龙的游禽类的种群数量低,而且不同年份存在较大波动(表2)。扎龙游禽类常见的鸟类的出现频度大多仅为0.135~0.330;而其他鸟类的频度就更低,大多低于0.05。

表2 扎龙保护区2006—2009年各种游禽的月平均遇见率和出现频度*
在4 a间,对各种鸟类来说不论是遇见率还是频度年度间都有一定的差异,多个种类呈下降趋势。
3.2.2 扎龙湿地游禽类群落的季节性变动
对每年不同月份游禽类数量变动进行分析,可以发现4 a来扎龙游禽类数量的年变动趋势较为相似(表3)。每年均在春季、秋季有两个高峰期,但春季的峰值高于秋季峰值。夏季种类最少,仅5种左右。
同样,游禽类种数的变动趋势也存在春秋季有两个高峰期,春季的峰值高于秋季峰值的现象(表3)。但是秋季的高峰出现的具体月份各年并不相同。由于2006和2009年的数据不连贯,不能确定这一峰值的分散性是由于年度间的差异造成。

表3 2006—2009年扎龙湿地游禽季节性变动
夏季记录到的游禽类种类数量均较少,这表明在扎龙湿地繁殖的游禽鸟类种类少,繁殖种群数量较低。目前常见的种类是小䴙䴘、风头䴙䴘、绿头鸭、斑嘴鸭、红头潜鸭。而过去常见的繁殖鸟骨顶鸡、罗纹鸭等鸟类现在仅见于迁徙季节。
同时,记录到的游禽种类、数量与个体数量在年度内呈现正比关系,说明扎龙游禽类群落动态主要受迁徙鸟类影响。整个冬季无游禽鸟类停留。4 a的结果显示同样的趋势。
3.3 本研究结果与历史同期游禽类数量的比较
由于无法获得1983年鸟类调查中游禽类的原始数据,故无法得知1983年夏秋调查中每种游禽类的确切遇见率。在此仅对不同种的优势等级进行比较。
1983 年夏秋季(6—9月份)调查中所见游禽类有23种,其中I级种1种;II级4种;III级3种;IV级12种;V级1种;VI级2种。本研究夏季(6—9月份)调查所见游禽鸟类有19种,其中II级5种;III级2种;IV级10种;VI级2种;无I级和V级物种(表4)。
1983 年同期的调查数据中,有7种在近4 a的调查中未见,另有4种游禽在1983年未见到而近年调查中有记录(表4)。值得指出的是在7种本次调查未见到的物种中赤颈鸭、花脸鸭、罗纹鸭、普通秋沙鸭、翘鼻麻鸭和针尾鸭在1983年调查中曾是普通种,尤其在秋季迁徙季节里常常可见大群。从优势度来看有13种雁鸭类鸟类数量呈现明显的下降趋势;6种鸟类数量基本稳定;7种鸟类数量呈现上升趋势。数量下降最为明显的游禽鸟类包括罗纹鸭、青头潜鸭和骨顶鸡3种,数量上升最明显的种类有红头潜鸭和小䴙䴘。
分别在扎龙游禽种类选取过去非常常见现在不常见的种、过去不常见的种现在常见的种和种群数量相对稳定3个类群中的几种游禽分析其种群动态。
结果显示:近20 a来数量下降的物种:骨顶鸡、青头潜鸭和罗纹鸭繁殖季节未见,在迁徙季节时的数量最大;绿头鸭虽有繁殖,但是数量不大。其中,骨顶鸡的数量变动最大,从原来数量极多的物种变为普通种(表4—表6)。如果从调查的频度来看,骨顶鸡4 a中的月平均频度低于0.1(平均值0.028~0.094,标准差0.056~0.120)。由此可见,骨顶鸡不但是数量低,而且出现的随机性也很强。而罗纹鸭的数量由原来的II级(数量很多的物种)降为夏秋无记录。罗纹鸭在本研究中仅在2007、2008年的4—5月份见到几次,遇见率和月平均频度都非常低。青头潜鸭原来在夏秋季也是数量很多的种,在本研究中虽每年都有1、2次记录,但仅仅出现在迁徙季节,数量低。20 a来游禽鸟类数量的下降是近些年在湿地内繁殖的鸟类种群数量下降或无繁殖鸟所导致的。

表4 扎龙湿地游禽鸟类变化趋势
数量上升的游禽类中,凤头䴙䴘、小䴙䴘和红头潜鸭常年可见,数量稳定在II级,频度在0.3左右。小䴙䴘数量在秋季迁徙时达到最大值。红头潜鸭数量在春季迁徙时数量大。豆雁是1983年调查中未记录到的种,现仅在迁徙季节里可见到数量2~50只不等的小群,秋季数量略大于春季,但观察到的随机性强,月平均频度低于1.6(平均值0~0.151,标准差0~0.345),在2009年的调查中未记录到此种(表5、表6)。
比较20 a来数量级相对稳定,在扎龙较常见的物种斑嘴鸭尽管种群数量不大(平均值0.728~2.834,标准差0.241~0.425),出现的频率稳定(平均值0.241~0.425,标准差0.110~0.253),全年(除冬季外)可见。其种群数量秋季高于其他季节(表5、表6)。绿翅鸭,尽管20 a的数量级相对稳定,但仅在迁徙季节见到小群,夏季只在2007年7月份记录到2只个体。因此,其调查频度很低(平均值0~0.069,标准差0~0.075)(表5、表6)。

表5 扎龙湿地几种游禽类不同年份逐月数量变动
4 讨论
4.1 扎龙湿地游禽类群落特征的改变
将调查结果与1983年研究结果比较,可以看出,扎龙湿地内游禽类群落中过去占优势的繁殖种群转变为迁徙鸟类种群。原来记录到37种游禽中有15种繁殖鸟,而本研究在夏季最多记录到7种游禽(表3)。这些缺失的物种多是鸭科鸟类,如罗纹鸭、青头潜鸭、赤颈鸭、琵嘴鸭、白眉鸭和普通秋沙鸭。尽管不能说这些物种在扎龙湿地的居留型发生了改变,但可以肯定地说这些物种即使有在扎龙湿地繁殖也仅为零星个体。骨顶鸡曾是扎龙湿地中数量极多的物种之一[7]19,在开阔的水域内随处可见,但现在只偶尔见到几个个体。除了那些这次调查中未记录到的游禽外,大部分物种只能在迁徙的季节里见到。
扎龙湿地即是多数水鸟的繁殖地,又位于东北亚迁徙水鸟通道的中部是迁徙水鸟的重要停息地。尽管本研究的结果显示在迁徙季节游禽的种群数量高于繁殖季节,然而无论是数量还是频度都大大低于之前的研究[7]19-21。20世纪90年代迁徙季节里赤颈鸭、花脸鸭、针尾鸭常常可见到几百只的大群,而本调查中最大群2007年4月19日记录到赤颈鸭40只;2008年4月24日花脸鸭68只和针尾鸭118只。由此可见,扎龙湿地游禽类种群数量的减少不仅是繁殖鸟,迁徙的种群的数量也处于下降的趋势。
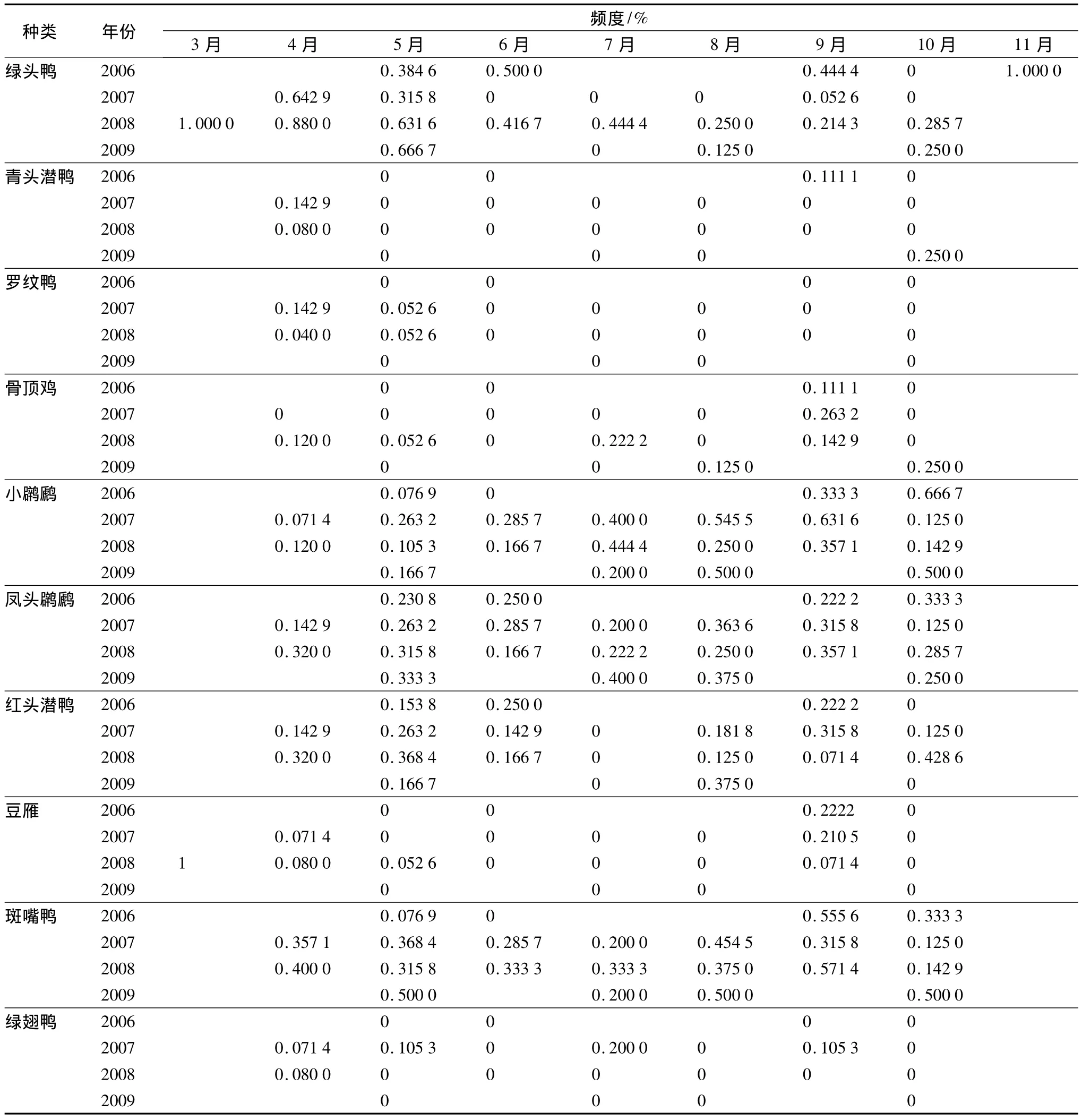
表6 扎龙湿地几种游禽类不同年份逐月频度变动
4.2 扎龙湿地水文和景观的变化对游禽类的影响
近年来,扎龙湿地的水文和景观特征发生巨大的变化[9-10,19-22],这必然影响到对湿地的水生环境的依赖性非常强的游禽类的栖息、生存。自然情况下,扎龙湿地内的水源主要来自于乌裕尔河水、双阳河水、当地区间径流、降水以及嫩江洪泛水等5部分。受上游农业生产和生活用水量增加、水库截留和水利工程阻隔和排泄导致湿地来水量减少[19,23];加之受全球大气候变化的影响,近年以来区域气温呈现上升趋势而降水量呈现下降趋势[22];同时,嫩江防洪堤坝的修建,嫩江干流中小洪水洪泛水量一般不再进入扎龙湿地[24],导致湿地蓄水量显著下降。最终表现为湿地内水量严重不足,扎龙湿地面积严重萎缩,其中湖泊湿地面积减少15.17%,湿地明水面面积下降49.36%,在2000年和2001年,湿地水位比平水年下降约lm,湿地面积约减少一半[19]。另外,近年来扎龙湿地内人工挖掘的沟渠、修筑堤坝和土路时遗留下来的沟渠的密度不断增加。这些沟渠加速了湿地内地表水向地下水转换的进程,增加了湿地内地表水的径流速度,降低了湿地原有的蓄水能力[25]。湿地补水后,水资源顺着沟渠以极快的速度流出湿地,未能在湿地内散漫。降低补水对湿地的恢复的能力。由于湿地水位持续下降,导致适宜于雁鸭等水鸟栖息的沼泽湿地面积减少生境破碎化[9,20,26]。本研究认为是水文系统的改变、湿地破碎化、栖息地条件恶化是本研究中雁鸭类等游禽的种类及数量明显减少的主要原因。
扎龙湿地水环境的变化改变了水生植物群落的组成可能是导致在淡水环境中觅食的游禽类种群数量下降的另一原因。由于湿地缺水,水量减少,不仅湿地面积缩小,同时导致水盐度增加;加上纳污量增加,水质发生变化[27]。水体盐碱度增大,雁鸭类、骨顶鸡等游禽的主要植物性食物之一的眼子菜属的沉水植物的生物量减少。这种食物资源的减少将间接地导致湿地内喜淡水的雁鸭类、骨顶鸡等水鸟数量减少[28]。
而这种水环境的变化对一些能够适应咸水和富氧化环境的物种产生有利影响,如红头潜鸭在淡水环境和咸水环境中其食性也会相应的发生转变[29]。这种食性的转变使红头潜鸭适应湿地内水体盐碱度增加导致的食物资源的变化。这是近年红头潜鸭种群数量增加的主要原因之一(表2、表5、表6)。
由于纳污量的增加,湿地内水资源分布在时间的波动性和空间上的不均匀性,导致湿地水体富营养化严重。这种情况恰恰形成了小䴙䴘这一类水鸟最适宜的觅食环境[30],这点与本研究观察到的小䴙䴘的种群数量增加一致(表5、表6)。
4.3 扎龙湿地补水对游禽鸟类数量变动的影响
针对扎龙湿地严重缺水的现象,黑龙江省政府自2002年开始通过中引工程向湿地补水,在一定时空范围内湿地得以恢复。本研究的调查路线之一,位于翁海排干下游,在这一区域通常见到的游禽多于其他区域。但是,现行的湿地补水时间多为春末夏初湿地干旱之时,此时已有很多雁鸭等水鸟在芦苇沼泽中营巢繁殖,补水后水位的突然提升,淹没鸟巢,降低游禽鸟类的繁殖成功率。放水时水流速度过快过急,对湿地植被冲击大,也破坏水鸟适宜的觅食和繁殖生境。同时,多数雁鸭类等水鸟不能在较快的水流区域内正常觅食,影响其在湿地内的生存。秋季多点补水应该是较好的方法,因为秋季植物生长末期,密度大,耐冲击力大;而鸟类处于非繁殖期,游禽多集中于较大的明水面中,水文变化对它们的影响不大。
湿地内的游禽鸟类对湿地内水文变化引起的生境类型的变化及人为活动的干扰极其敏感,可以用游禽鸟类数量的变动趋势指示生态环境的变化。但是游禽鸟类对湿地环境的变化有一定的滞后性,短期的游禽鸟类资源监测不能完整的指示生态环境的变化。因此,长期的鸟类和植物群落常规监测工作,才能完整真实的反应生态环境变化。鉴于扎龙湿地目前的状况,严格禁止进一步对湿地水文的改变和有效的湿地恢复措施非常重要。
致谢:本研究受到由联合国环境署/全球环境基金(GEF)开展的“为了保护西伯利亚鹤及其它亚洲迁移水禽而形成与发展的湿地样地与鸟类迁移路径网络”项目资助,在课题设计方面受益于项目专家指导组;扎龙保护区对本研究的实施给予支持;参与野外调查工作的有关雪、刘福剀、汪青雄、曲文慧、刘磊等同学,在此一并表示感谢。
[1]Wiens J A.Foundations and patterns:Volume 1[M].Cambridge University Press,1989:539.
[2]Mistry J,Andrea B,Simpson M.Birds as indicators of wetland status and change in the North Rupununi,Guyana[J].Biodiversty and Conservation,2008,17(10):2383-2409.
[3]Kantrud H A,Stewart R E.Ecological distribution and crude density of breeding birds on prairie wetlands[J].The Journal of Wildlife Management,1984,48(2):426-437.
[4]郑光美.我国鸟类生态学的回顾与展望[J].动物学杂志,1981(1):63-68.
[5]郑作新.中国鸟类分布名录[M].北京:科学出版社,1976.
[6]苏立英,马建章,许杰,等.扎龙保护区鸟类区系的初步研究[J].东北林业大学学报,1987,15(2):62-72.
[7]高中信,费殿金,李春源,等.扎龙鸟类[M].北京:中国林业出版社,1989:14-22.
[8]李枫,鲁长虎,杨红军,等.扎龙芦苇沼泽繁殖鸟类群落多样性研究[J].东北林业大学学报,1998,26(5):68-72.
[9]韩敏,孙燕楠,许士国,等.基于RS,GIS技术的扎龙沼泽湿地景观格局变化分析[J].地理科学进展,2005,24(6):42-49.
[10]黄芳,王平,王永洁,等.扎龙湿地生态环境变化及其对白鹤迁徙的影响[J].东北师大学报:自然科学版,2007,39(2):106-111.
[11]李枫,王强,蔡勇军,等.扎龙春季火烧对丹顶鹤营巢以及芦苇生态的影响[M]//王岐山,李凤山.中国鹤类研究.昆明:云南教育出版社,2007:129-134.
[12]Wang Q,Li F.Nest raising by red-crowned cranes in response to human-mediated flooding at Zhalong Nature Reserve,China[J].The Wilson Journal of Ornithology,2008,120(3):610-613.
[13]邹红菲,吴庆明,马建章.扎龙保护区火烧及湿地注水后丹顶鹤(Grus japonensis)巢址选择[J].东北师大学报:自然科学版,2003,35(1):54-59.
[14]邹红菲,吴庆明,史荣红.扎龙湿地恢复初期丹顶鹤孵化期觅食生境选择[J].东北林业大学学报,2007,35(7):55-58.
[15]刘昌明,夏军,于静洁.东北地区水与生态环境问题及保护对策研究:东北地区有关水土资源配置、生态与环境保护和可持续发展的若干战略问题研究(生态环境卷)[M].北京:科学出版社,2007:135-147.
[16]中华人民共和国林业部.扎龙国家级自然保护区管理计划[M].北京:中国林业出版社,1997:1-8.
[17]许龙,张正旺,丁长青.样线法在鸟类数量调查中的应用[J].生物学杂志,2003,22(5):127-130.
[18]尹达云.水禽生物学研究的基本内容及其研究方法[J].动物学杂志,1960,5:206-211.
[19]佟守正,吕宪国,苏立英,等.扎龙生态系统变化过程及影响因子分析[J].湿地科学,2008,6(2):179-184.
[20]张玉红,李晓峰,张树清.基于MATLAB的扎龙湿地归一化差值指数时间变化分析[J].东北林业大学学报,2008,36(3):39-40.
[21]舒展.火烧与缺水对扎龙湿地植被群落的影响[J].环境科学与管理,2010,35(1):135-139.
[22]沃晓棠,黄智超,孙彦坤,等.扎龙湿地近50年温度和降水年际变化分析[J].东北农业大学学报,2010,41(5):61-65.
[23]姜宝玉,韩玉梅,曹波,等.试论扎龙湿地与水资源优化配置问题与对策[J].黑龙江水利科技,2003(2):5-6.
[24]刘开棘.扎龙湿地水资源利用现状分析及对策[D].北京:北京交通大学经济管理学院,2008.
[25]郗敏,吕宪国,姜明.人工沟渠对流域水文格局的影响研究[J].湿地科学,2005,3(4):310-314.
[26]张玉红.扎龙湿地景观动态监测与分析[D].北京:中国科学院研究生院,2010.
[27]王建成,张洪斌,王丽媛,等.论乌裕尔河水污染对扎龙自然保护区生态环境的影响[J].东北水利水电,1997,9:31-33.
[28]Hamdi N,Charfi F,Moali A.Variation of the waterbird community relying to the Ichkeul National Park,Tunisia[J].European Journal of Wildlife Research,2008,54(3):417-424.
[29]Olney P J S.The food and feeding-habits of the Pochard,Aythya ferina[J].Biological Conservation,1968,1(1):71-76.
[30]Ceccobelli S,Corrado B.On the water depth in diving sampling sites of Tachybaptus ruficollis[J].Rend Fis Acc Lincei,2010,21(4):359-364.
猜你喜欢
青年文学家(2023年4期)2023-05-30 10:48:04
中国动物保健(2021年3期)2021-11-23 05:02:10
伙伴(2020年4期)2020-05-11 06:15:11
家禽科学(2020年2期)2020-04-14 04:57:59
经济技术协作信息(2018年22期)2019-01-19 02:59:58
当代畜禽养殖业(2018年12期)2018-04-12 21:27:52
青年文学家(2017年28期)2017-11-28 17:27:53
妇女之友(2017年3期)2017-04-20 09:20:00
兽医导刊(2017年16期)2017-04-05 11:36:42
中国药物应用与监测(2015年5期)2015-12-11 03:15:55
