
幽门螺杆菌cag4蛋白的原核表达、纯化及鉴定①
2011-01-23 12:02王文凯谢立苹邵世和江苏大学附属人民医院检验科镇江212002
中国免疫学杂志 2011年4期
王文凯 钟 桥 谢立苹 邵世和 (江苏大学附属人民医院检验科,镇江212002)
溶菌糖基转移酶(Lytic transglycosylase,LT)广泛分布于细菌,能够水解N-乙酰胞壁酸酯(N-acetylmuramic acid,MurNAc)和 N-乙酰葡糖胺(N-acetylglucosamine,G lcNAc)之间的β-1,4糖苷键。与溶菌酶不同的是其不是发生水解作用,而是发挥糖基转移酶的活性,即在N-乙酰胞壁酸酯MurNAc内部形成了1,6糖苷键。由于该酶广泛的参与细菌的分裂繁殖、生物被膜形成以及细菌毒性蛋白的转运,因此与细菌的致病性密切相关[1]。其中,广泛分布于细菌的Ⅲ、Ⅳ型分泌系统中的溶菌糖基转移酶,可以水解肽聚糖层,使得胞内蛋白进入周浆间隙,促进分泌转运装置的装配形成,从而使得细菌发挥毒性作用[2]。而在幽门螺杆菌(Helicobacter pylori,简称 H.pylori)Cag致病岛中,也编码一个Ⅳ型分泌样装置,负责转运毒性蛋白CagA进入宿主细胞,发挥毒性作用[3]。近年来,虽然Cag致病岛Ⅳ型分泌样装置的研究取得了一定的进展,但该分泌装置的装配形成机制尚未明确[4]。其中,幽门螺杆菌cag4蛋白是 H.pylori Cag致病岛编码的溶菌糖基转移酶,可以在局部水解细菌的肽聚糖层,使得胞内蛋白释放进入周浆间隙,促进分泌系统的装配形成。本文利用原核表达技术成功地获得具有良好的水解活性的cag4重组蛋白,并对其酶活性和理化特性做了初步鉴定;旨在进一步阐明幽门螺杆菌致病机制,为新型抗生素(或是抗菌药物)的研发提供作用靶位。
1 材料与方法
1.1材料
1.1.1菌株 H.pyloriNCTC11637由中国疾病控制中心传染病预防研究所张建中教授馈赠,大肠埃希菌DH5α及BL21(DE3)、溶壁微球菌为江苏大学基础医学与医学技术学院中心实验室保存。
1.1.2试剂和仪器 哥伦比亚培养基、厌氧袋购自OXOID公司;Ex TaqDNA聚合酶、dNTP、限制性核酸内切酶 EcoR Ⅰ及 Xho Ⅰ、T4-DNA连接酶、IPTG、DL2000 DNA Marker及蛋白质分子量标准(低)购自TaKaRa公司;DL1Kb DNA marker购自金思特生物有限公司;Ni2+-NTA购自Qiagen公司;pGEM-T载体购自Promega公司;pET-28a载体由江苏大学医学技术学院中心实验室保存;超速离心机购自Beckman公司;其他常规试剂按照《分子克隆实验指南》要求配制。
1.2幽门螺杆菌cag4蛋白基因的克隆 根据幽门螺杆菌cag4蛋白基因的全长序列,设计上、下游引物:P1:5′-CGGG AATTCTTGTTTGGG AAATGG AT-3′;P2 :5′-ATTCTCG AGCTACTCGTTATATCGCACTT-3′,在其5′端分别引入 EcoR Ⅰ、XhoⅠ酶切位点,PCR产物长度为510 bp。引物由上海生工生物技术有限公司合成。以 H.pyloriNCTC11637菌株基因组DNA为模板,用 Ex Taq聚合酶进行 PCR扩增(25μl反应体系)。扩增参数为:94℃5分钟,94℃30秒、52℃30秒、72℃1分钟,30个循环,72℃10分钟。PCR产物经1.0%琼脂糖凝胶电泳,EB染色,GeNius凝胶电泳图像分析系统分析鉴定并用胶回收试剂盒回收PCR产物。
1.3T-A克隆和原核表达载体pET-28a-cag4的构建胶回收的 PCR产物与pGEM-T载体,加1μl T4-DNA连接酶,4°C连接过夜。连接产物转化 E.coli DH5α感受态细胞。转化后的细菌涂布于含50μg/ml氨苄青霉素的LB平板上,37℃培养16小时。挑取菌落转种于含氨苄青霉素50μg/ml的LB液体培养基,37℃振摇12小时,用碱裂解法进行质粒提取,并进行EcoRⅠ、XhoⅠ双酶切鉴定。
胶回收酶切目的片段,构建表达pET-28a-cag4,转化感受态细菌 E.coliBL21(DE3),转化好的细菌涂布含100μg/ml卡那霉素的LB琼脂培养基培养12~16小时,挑选单菌落接种于含卡那霉素的LB液体培养基中,置37℃摇床振荡培养12小时,抽提质粒,酶切鉴定。
1.4蛋白诱导表达和鉴定 经酶切鉴定构建成功的重组工程菌,接种于含100μg/ml卡那霉素的LB液体培养基中,在37℃振荡至菌液OD600达到0.6,加入 IPTG诱导后,收集菌液进行SDS-PAGE鉴定。同时,设立阴性对照。
取诱导后的菌液进行SDS-PAGE电泳后,然后4℃电转移至PVDF膜上,用含5%脱脂奶粉的封闭液室温封闭3小时,换新鲜的封闭液,加入鼠抗His单克隆抗体作为一抗,37℃摇床反应1小时。1×PBST洗涤3~5次,每次5分钟,于1×PBST中加入辣根过氧化物酶(HRP)标记的羊抗兔 IgG作为二抗,37℃摇床反应1小时,最后DAB显色至目的条带清晰时将膜转移至双蒸水终止反应。
1.5蛋白的纯化和复性 通过比较不同诱导时间和IPTG浓度后,确定了最佳诱导条件,即1 mmol/L IPTG,诱导4小时。细菌活化后,按照1∶200稀释后加入500 ml LB中,进行大量诱导。收集菌体后,重悬于 lysis buffer(50 mmol/L NaH2PO4·2H2O,300 mmol/L NaCl,5 mmol/L iminazole和8 mmol/L urea[pH8.0])中,冰上超声破碎(200 W)。离心后,收集上清和镍柱Ni2+-NTA(Qiagen)室温结合1小时后纯化。洗涤去除蛋白后,利用 Elute buffer(50 mmol/L NaH2PO4·2H2O,100 mmol/L NaCl,200 mmol/L imidazole,8 mmol/L urea,和 5 mmol/L DTT[pH8.0])进行洗脱,收集蛋白后,SDS-PAGE鉴定。
蛋白透析之前将透析袋放入含2%NaHCO3和1 mmol/L EDTA的蒸馏水中煮沸10分钟,用蒸馏水反复冲洗,再放入含1 mmol/L EDTA的蒸馏水煮沸10分钟,自然冷却,加入待透析的蛋白,分别用含6、4、3、2和1 mol/L尿素 PBS在4℃条件下梯度透析,各4小时,最后用不含尿素的PBS平衡过夜。收集透析液,SDS-PAGE鉴定。
1.6细菌肽聚糖的提取与酶谱分析 参照Zeiger[5]报道的方法并加以改良,具体按以下步骤进行。
溶壁微球菌接种LB培养基后,37℃培养过夜。收集细菌后,重悬于20 ml冰预冷的25 mmol/L磷酸盐缓冲液中。将菌液逐滴加入煮沸的8%SDS溶液中,煮沸1小时,1 000 r/min 15℃离心30分钟收集后沉淀,再重复煮沸一次。用磷酸盐缓冲液洗涤沉淀重复5次后,再利用超速离心机4 000 r/min 25℃离心30分钟后,收集后乙醚脱脂后,干燥获得肽聚糖。
酶谱分析,即将收集的肽聚糖作为底物,掺入12%的分离胶中,进行SDS-PAGE电泳[6]。电泳后,采用复性缓冲液(20 mmol/L sodium phosphate buffer[pH7.0],0.1%Triton X-100和10 mmol/L MgCl2),37℃孵育48小时。采用1%的亚甲基蓝染色后,双蒸水脱色。实验过程,设立溶菌酶和牛白蛋白分别为阳性和阴性对照。
1.7浊度实验 将热致死后的溶壁微球菌,重悬于不同pH值的磷酸盐缓冲液(pH5.0~7.0)中,调整到其浊度在OD600为1.0。分别加入5μg/ml的重组蛋白后,37℃孵育,监测其浊度值的变化速率,计算ΔA·min-1mg-1(protein)值代表酶活性的大小[7,8]。
1.8统计学处理 每组样本测定重复3次。所有检测值均以±s表示。组间差异的比较采用两样本间 t检验,P<0.05为差异具有显著性。
2 结果
2.1幽门螺杆菌cag4蛋白与其他溶菌转移酶的氨基酸序列同源性分析 从NCBI数据库,检索获得不同细菌分泌系统中溶菌糖基转移酶的基因序列。再根据PFAM数据库,搜索分析这些溶菌糖基转移酶的催化保守区域,再采用clustalX 1.8将这些催化区域进行多重序列比较分析后,利用GenDoc软件显示结果。结果显示,序列之间具有一定的保守性。其中,幽门螺杆菌 cag4蛋白中第 56位氨基酸(G LU56),被推测认为是发挥溶菌糖基转移酶的催化活性的活性中心。
2.2原核表达载体pET-28a-cag4的构建 PCR结果显示在510 bp左右有一条带,与预计大小一致,无非特异性条带。将PCR产物与pGEM-T载体连接产物转化至DH5α,随机挑取菌落,抽提质粒,双酶切产物电泳后,阳性克隆出现的两条带分别位于约3 003 bp和510 bp处(见图1)。
质粒pET-28a和TA克隆鉴定阳性的质粒分别经EcoRⅠ和XhoⅠ双酶切后,回收酶切片段,16℃连接过夜,连接产物转化至 E.coliBL21,随机挑取菌落,抽提质粒,双酶切产物电泳后,阳性克隆出现的两条带分别位于5.3 kb和510 bp处(见图2)。
2.3幽门螺杆菌cag4蛋白重组蛋白的表达与纯化

图1 pGEM-T-cag4质粒双酶切鉴定图Fig.1 Double restriction enzyme digestion map of the recombinant plasmid pGEM-T-cag4

图2 pET-28a-cag4质粒酶切鉴定图Fig.2 Double restriction enzyme digestion map of the recombinant plasmid pET-28a-cag4
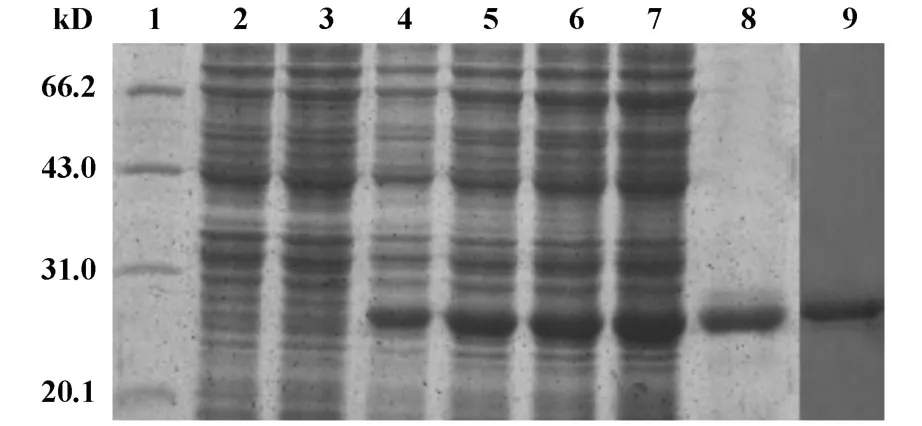
图3 SDS-PAGE和Western blot鉴定结果图Fig 3 The result of SDS-PAGEand Western blot
原核表达载体pET-28a-cag4转化至 E.coliBL21中获得重组工程菌,接种LB液体培养基,于37℃培养至OD600达到0.6时,加入终浓度为1 mmol/L IPTG诱导4小时,收集菌体进行SDS-PAGE检测,可见在约Mr23kD处出现1条新的蛋白带(见图3),与理论预测值基本相符。空载体转化菌pET-28a/BL21诱导后和未诱导重组工程菌在同一位置未出现相应的蛋白带。将诱导后的全菌进行SDS-PAGE分离后电转移至NC膜上,依次加入一抗体和二抗,DAB显色后在相对分子量为23 000的新生蛋白带处,有一特异性条带,同时以空载体菌为对照,未发现条带,说明该蛋白带为目的蛋白,见图3。育处理。结果如图4可见,加入溶菌酶和重组蛋白幽门螺杆菌cag4蛋白的泳道2、3,都有明显的溶解痕迹,而蛋白的泳道1未见。其中,溶菌酶的分子量大小约为14.4 kD,而幽门螺杆菌cag4蛋白约为23 kD。结果说明幽门螺杆菌NTCT11637 cag4蛋白具有溶菌糖基转移酶活性。
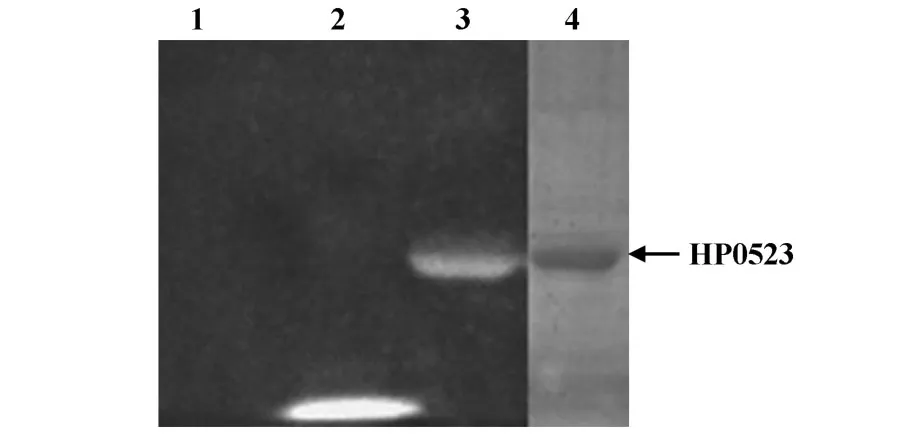
图4 酶谱分析结果图Fig.4 The zymogram analysis of cag pathogenicity island protein 4

图5 重组cag4在不同pH条件下的酶活性大小Fig.5 The enzyme activity of recombinant cag pathogenicity island protein 4 at different pHvalues
2.5不同pH条件下的酶活性变化 将溶壁微球菌重悬于不同pH值的磷酸盐缓冲液(5.0,6.0,7.0)中,调整OD600为1.0,分别加入浓度为5μg/ml的重组 CAG4蛋白,37℃作用,间隔 15分钟测定OD600,连续测定2小时,每次重复测定3次。结果如图5所示,发现pH6.0组的OD600下降速率,较其他组呈现显著性差异(P<0.05)。说明幽门螺杆菌NTCT11637 cag4蛋白在偏酸性环境(pH6.0)下,其酶活性较强。采用ΔA·min-1mg-1值计算不同pH组酶活性大小,每次结果重复测定3次,计算均值和标准差,结果以±s表示;组间差异采用两样本t检验,结果表明P<0.05,呈显著性差异,如图所示,pH6.0时的该酶活性较强。
3 讨论
收集菌体溶解于裂解液中,冰上30分钟后,超声破碎,10 000×g离心收集上清和沉淀,沉淀溶解于裂解液中,SDS-PAGE电泳分析目的蛋白的溶解度,发现目的蛋白主要存在于沉淀中,即以包涵体形式存在表达,故采用变性条件下纯化蛋白。纯化后,采用SDS-PAGE电泳鉴定表明,蛋白纯度达98%以上,见图4。纯化后蛋白,进一步采用透析法进行蛋白的复性处理。
2.4重组蛋白酶活性的鉴定 复性处理后的蛋白,加入掺入溶壁微球菌为底物的聚丙烯酰氨凝胶中,100 V 2小时电泳分离后,进行复性处理和37°C孵
H.pylori感染在世界范围内流行,现已证实感染H.pylori者,特别是 Ⅰ型H.pylori(含 Cag-PAI),可以导致慢性胃炎、消化性溃疡,甚至胃癌和粘膜相关性淋巴瘤(MALT)的发生[3]。现已证明,CagPAI是编码一个IV型样分泌系统(type IV like secretion system)转运CagA蛋白进入宿主细胞,发挥毒性作用,导致一系列的病理变化[3]。虽然目前对该结构的研究取得了一定的进展,但仍有许多问题尚未阐明,如其装配形成机制,底物蛋白识别机制等等。然而,细菌的肽聚糖层作为天然的屏障,给该多蛋白复合物的组装形成,造成了很大的困难。因此,分泌系统中发挥溶菌功能的组分,如VirB1、ORF169等,在多蛋白复合物装配形成过程中起重要的作用。然而,这些溶菌糖基转移酶在氨基酸序列上存在着很大的差异,可能仅仅是部分序列发挥水解肽聚糖功能,如根瘤农杆菌VirB/D转运系统中VirB1[9]。因此,我们将不同细菌分泌系统中的溶菌糖基转移酶,利用PFAM数据库检索获得其发挥酶活性的保守功能域,进行多重序列之间的比对研究。结果表明cag4基因和其他细菌分泌系统中溶菌糖基转移酶具有高度的同源性。其中,在α-螺旋结构的末端存在一个保守的谷氨酸(G lu),可以形成一个催化的活性中心,发挥催化活性作用。结合Slt35的三维晶体结构研究表明,在溶菌糖基转移酶的氨基酸序列中还存在一个保守的AVG AY基序,可能是与肽聚糖结合作用的功能域[10](图1)。
临床上H.pylori感染常采用根除治疗,但根除之后可以导致体内的菌群比例失衡,易产生新的胃肠道疾病。因此,根除疗法势必不是理想的治疗手段。另一方面,随着对克拉霉素和甲硝唑等抗生素的原发耐药率普遍上升,也对这种治疗方案提出了新的挑战。寻找和研制新型抗生素或是抗菌药物迫在眉睫。溶菌糖基转移酶是细菌分泌毒性蛋白关键酶之一,具有良好的保守性,是新型抗生素良好的候选作用靶位。此次利用原核表达技术,重组获得了大量具有酶活性的幽门螺杆菌cag4蛋白,为其结构与功能的研究和生物抑制剂的研发奠定了坚实基础。
溶菌糖基转移酶,与溶菌酶不同的是能在N-乙酰胞壁酸酯MurNAc内部形成了1,6糖苷键。而国外学者通过对其水解产物进行HPLC-MS质谱分析发现,其主要的水解产物是胞壁肽、四肽(Muropeptide G lcNAc-(1,6 anhydro) MurNAc-tetrapeptide)等[11,12]。而这些水解产物,可以被胞内受体Nod1识别,活化初始免疫应答,导致炎症因子释放[13,14]。因此,我们可以推测:H.pylori在感染过程中,幽门螺杆菌cag4蛋白不仅仅水解肽聚糖,促进分泌系统的装配而发挥致病作用,还可以利用其水解产物,诱导炎症反应。此外,在分泌系统装配过程中,溶菌糖基转移酶,水解肽聚糖释放蛋白质进入周浆间隙,组装形成分泌装置。但这种作用释放势必要得到及时的终止,才能避免多蛋白的聚集,而影响其装配形成效率。那么,细菌又是如何控制其表达,或是分泌其他抑制剂来终止这种反应呢?这些问题都值得进一步深入研究。
1 Scheurwater E,Reid C W,Clarke A J.Lytic transglycosylases:bacterial space-making autolysins[J].Int J Biochem Cell Biol,2008;40(4):586-591.
2 Zahrl D,Wagner M,Bischof Ket al.Peptidoglycan degradation by specialized lytic transglycosylases associated with type III and type IV secretion systems[J].Microbiology,2005;151(Pt 11):3455-3467.
3 Olbermann P,Josenhans C,Moodley Yet al.A global overviewof the genetic and functional diversity in the Helicobacter pylori cag pathogenicity island[J].PLoS Genet,2010;6(8):e1001069.
4 Zhong Q,Shao S H,Cui L Let al.Type IV secretion systemin Helicobacter pylori:a new insight into pathogenicity[J].Chinese Medical J,2007;120(23):2138-2142.
5 Zeiger A R,Wong W,Chatterjee A Net al.Evidence for the secretion of soluble peptidoglycans by clinical isolates of Staphylococcus aureus[J].Infect Immun,1982;37(3):1112-1118.
6 Blackburn N T,Clarke A J.Assay for lytic transglycosylases:a family of peptidoglycan lyases[J].Analytical Biochemistry,2000;284(2):388-393.
7 Zhao Y P,Zhang H,Yang Y J.Improvement of lysozyme measurement[J].Food Sci Technol,2002;23(3):116-119.
8 Li D H,Chi YJ.Facility detection of lysozyme activity[J].Chin Dairy Ind,2002;30(5):128-129.
9 Zupan J,Hackworth C A,Aguilar Jet al.VirB1*promotes T-pilus formation in the vir-Type IV secretion system of Agrobacterium tumefaciens[J].J Bacteriology,2007;189(18):6551-6563.
10 van Asselt E J,Dijkstra A J,Kalk K Het al.Crystal structure of Escherichia coli lytic transglycosylase Slt35 reveals a lysozyme-like catalytic domain with an EF-hand[J].Structure,1999;7(10):1167-1180.
11 Fukushima T,K itajima T,Yamaguchi Het al.Identification and characterization of novel cell wall hydrolase CwlT:a two-domain autolysin exhibiting n-acetylmuramidase and DL-endopeptidase activities[J].J Bio Chem,2008;283(17):11117-11125.
12 Viollier P H,Shapiro L.A lytic transglycosylase homologue,PleA,is required for the assemblyof pili and the flagellum at the Caulobacter crescentus cell pole[J].Molecular microbiology,2003;49(2):331-345.
13 Inohara N,Ogura Y,Nunez G.Nods:a family of cytosolic proteins that regulate the host response to pathogens[J].Current Opinion in Microbiology,2002;5(1):76-80.
14 Viala J,Chaput C,Boneca I Get al.Nod1 responds to peptidoglycan delivered by the Helicobacter pylori cag pathogenicity island[J].Nature Immunology,2004;5(11):1166-1174.
猜你喜欢
保健与生活(2022年11期)2022-06-09
中国典型病例大全(2022年7期)2022-04-22
中老年保健(2021年5期)2021-12-02
现代临床医学(2021年6期)2021-11-20
现代临床医学(2021年5期)2021-11-02
保健与生活(2021年5期)2021-04-12
中华养生保健(2020年9期)2021-01-18
天然产物研究与开发(2018年8期)2018-09-10
中外医疗(2015年16期)2016-01-04
中外医疗(2015年11期)2016-01-04
