
抗番茄花叶病毒RNAi载体的构建及烟草遗传转化
2011-01-08 08:49赵峰玉郑银英张瑜乔亚红崔百明向本春
石河子大学学报(自然科学版) 2011年2期
赵峰玉,郑银英,张瑜,乔亚红,崔百明,向本春
(石河子大学农业生物技术重点实验室/石河子大学生命科学院,石河子832003)
抗番茄花叶病毒RNAi载体的构建及烟草遗传转化
赵峰玉,郑银英,张瑜,乔亚红,崔百明,向本春
(石河子大学农业生物技术重点实验室/石河子大学生命科学院,石河子832003)
为了获得抗番茄花叶病毒的转基因烟草,试验采用RT-PCR技术扩增新疆加工番茄花叶病毒基因组,然后与已知株系序列比对获得相应的保守序列。根据保守区选择干扰序列并设计引物进行 PCR扩增,构建了pBi35STo5,pBi35SToBK,pBi35ToMV3 3个RNAi表达载体;利用农杆菌介导法分别将表达载体侵染普通烟,转化植株经分子鉴定,结果显示已成功获得了转基因阳性植株,为进一步的攻毒试验奠定了基础。
加工番茄;RNAi;番茄花叶病毒
番茄花叶病毒(Tomato mosaic virus,ToMV)是一类世界性分布的植物病毒,其寄主范围很广,能侵染茄科、十字花科等许多植物,其中番茄是其主要寄主[1]。在新疆许多加工番茄种植区,番茄病毒病大面积的普遍发生,症状十分复杂,有的地块发病率高达100%,造成产量下降,甚至绝产[2]。近年来,该病害在新疆有逐年加重的趋势,严重影响了加工番茄的产量和品质。
番茄花叶病毒(ToMV)属烟草花叶病毒属,病毒粒子为杆状,其基因组为单链正义RNA,共编码4个蛋白:130/180 kDa复制酶蛋白、30 kDa移动蛋白(MP)和17 kDa外壳蛋白(CP)。ToMV的复制酶蛋白是由进入寄主细胞的基因组RNA直接翻译而成,而CP和MP是在病毒复制过程中产生的亚基因组翻译生成[3-4]。其复制酶蛋白在病毒的复制中起主要作用,MP主要参与病毒在细胞间的运输,而CP在辅助病毒衣壳化、识别宿主、病毒的长距离运输中起到重要作用。
目前,已有报道将病毒外壳蛋白和复制酶基因导入烟草可在一定程度上增强对病毒的抗性,但其抗性主要在病毒侵染初期,可以推迟发病,但难以有效持久地控制病毒的危害[5-6]。随着RNAi的机制的逐步被阐明,其特异性和有效性逐渐被人们所认识,是近年来迅速发展起来的一项基因阻隔技术[7-8],正越来越多的应用于植物抗病毒的研究。Waterhouse等[9]对马铃薯 Y病毒(PV Y)蛋白酶基因片段构建 IRS载体进行烟草转化,获得抗性植株;朱俊华等[10]将马铃薯Y病毒外壳蛋白基因部分序列反向重复转化入烟草,获得对PV Y免疫烟草植株。颜培强等[11]利用 TMV、CMV CP部分序列的融合基因片段构建干扰载体,转化烟草后,获得双抗的转基因植株。
本实验在成功克隆 ToMV序列的基础上,设计合成ToMV复制酶蛋白和CP特异性RNAi干扰片段,利用pBi35SG12载体构建 ToMV的 siRNA的表达载体,载体通过农杆菌介导法转化烟草,以期为下一步通过攻毒试验筛选高效表达载体,为最终获得抗ToMV转基因加工番茄奠定基础。
1 材料与方法
1.1 材料
1.1.1 菌株和质粒
大肠杆菌DH5α、农杆菌 GV3101和质粒载体pBi35SG12,由本实验室保存。
1.1.2 试剂
逆转录酶 (AMV)、限制性内切酶、T4DNA连接酶和 PCR试剂盒购自 Fermentas公司;Wizard DNA clean-up kit凝胶回收试剂盒购自Promega公司;Plant Genomic DNA Kit植物基因组DNA提取试剂盒购自 TIANGEN公司;MS培养基各成分购自上海生工;Trizol试剂购自 Invitrogen公司。普通烟(N icotiana occidentalis)由本实验室提供。
1.2 方法
1.2.1 番茄花叶病毒的鉴定
自新疆石河子周边田间采集带有病毒病症状的加工番茄叶片,用 Trizol法提取总 RNA,然后用逆转录酶(AMV)合成cDNA,具体操作参照产品说明书。
根据已报道的 ToMV保守序列设计检测的引物:
ToMVIDSP,5′-GTGGACATGCCGGTGCTAGA-3′;
TToVASP,5′-A GATTCGACACCGCA GCAGA-3′。
以合成的cDNA为模板进行降落 PCR扩增,反应条件为:94℃预变性5 min,94℃变性30 s,65℃退火30 s,72℃延伸30 s,每循环降1℃,共15个循环;最后94℃30 s;50℃30 s;72℃30 s,共20个循环;72℃延伸7 min。
PCR产物经1%琼脂糖凝胶电泳后确定片段大小,鉴定样品中是否含有番茄花叶病毒[1,12]。
1.2.2 番茄花叶病毒的分离、纯化
取检测带有番茄花叶病毒的加工番茄病叶,利用摩擦接种(缓冲液为0.02 mol/L PBS缓冲液,p H 7.0)至心叶烟(N icotiana glutinosa)上,通过连续5次单斑分离,得到纯化的 ToMV分离物CJ-2。
1.2.3 ToMV基因组测序
取纯化后的番茄花叶病毒烟草病叶,提取病毒总RNA。用逆转录酶合成cDNA第一链。
根据已报道的 ToMV序列设计阅读框引物(表1),以cDNA为模板进行 PCR扩增,反应条件:94℃预变性5 min,94℃变性30 s,65℃退火30 s,72℃延伸30 s,每循环降1 ℃,共15个循环;最后94℃30 s;50℃30 s;72℃30 s,共20个循环;72℃延伸7 min。
PCR产物经1%琼脂糖电泳后用Wizard DNA Clean-up kit试剂盒回收,并将回收的目的片段用T4DNA连接酶连到 T-Easy Vector上,转化大肠杆菌DH5α,经 EcoRI酶切鉴定后,将阳性克隆送到上海生工公司测序。然后用测序结果拼接 ToMV基因组。
将得到的 ToMV全长序列输入NCBI进行不同株系序列比对,找出保守序列区域,确定RNAi的靶序列。

表1 扩增ToMV序列所用引物Tab.1 The primers of ToMV sequences amplification
1.2.4 RNAi载体的构建
以引物 TToVSP1和 TToVASP1 PCR扩增序列为模板,设计其引物 SP/5′LP和 ASP/5′P,通过PCR扩增其RNAi的靶序列,将干扰序列的Bam-HI/KpnI双酶切片段和SacI/SpeI双酶切片段分别插入到 pBi35SG12相应位点,构建 pBi35STo5 RNAi载体(表 2)。
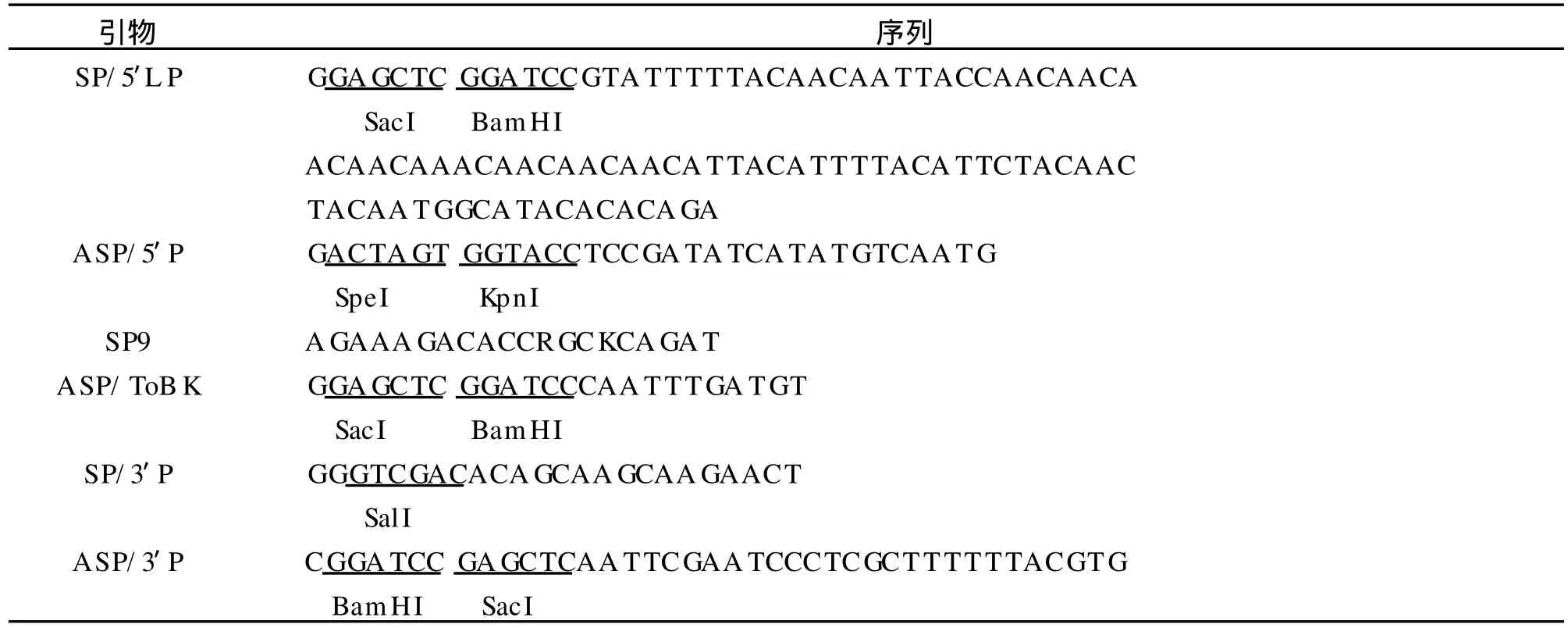
表2 扩增干扰序列所用引物Tab.2 The primers of RNAi sequences amplification
以引物 TToVSP9和 TToVASP8 PCR扩增序列为模板,设计其引物 SP9和 ASP/ToBK,通过PCR扩增其RNAi的靶序列,将干扰序列的Bam-HI/KpnI双酶切片段和XbaI/SacI双酶切片段分别插入到 pBi35SG12相应位点,构建pBi35SToBK RNAi载体(表 2)。
以引物 TToVSP12和 TToVASP11 PCR扩增序列为模板,设计其引物 SP/3′P和 ASP/3′P,通过PCR扩增其 RNAi的靶序列,将干扰序列的 SacI/SpeI双酶切片段和BamHI/SalI双酶切片段分别插入到pBi35SG12相应位点,构建pBi35SToMV3 RNAi载体(表 2)。
1.2.5 烟草转化及鉴定
提取表达载体质粒并纯化,用电击转化方法导人农杆菌 GV3101中,并用菌落PCR方法鉴定出阳性克隆。采用叶盘法通过农杆菌介导将表达载体整合到普通烟基因组中,受侵染的普通烟暗培养2 d后,转移至具有一定选择压的分化培养基,出芽后转到生根培养基中,培养条件:28℃、光照16 h/d。
用Plant Genomic DNA Kit植物基因组DNA提取试剂盒提取转化株叶片基因组DNA,具体操作参照产品说明书,以其基因组DNA为模板进行PCR鉴定。
2 结果与分析
2.1 番茄花叶病毒的鉴定
利用RT-PCR技术,以健康的番茄为阴性对照,从11份疑似病毒病的加工番茄样品,均扩增出约557bp的片段,说明加工番茄样品均被 ToMV侵染(图 1)。
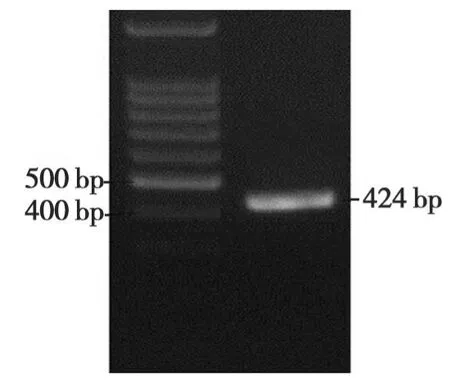
图2 干扰片段的PCR扩增Fig.2 PCR product of RNAi fragment
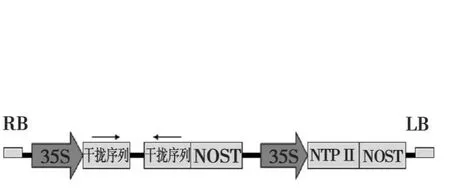
图3 RNAi载体构建Fig.3 The construction of RNAi vector
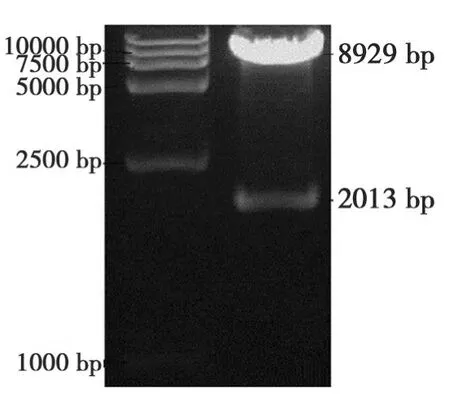
图4 pBi35STo5表达载体酶切鉴定Fig.4 Restriction identification of pBi35STo5
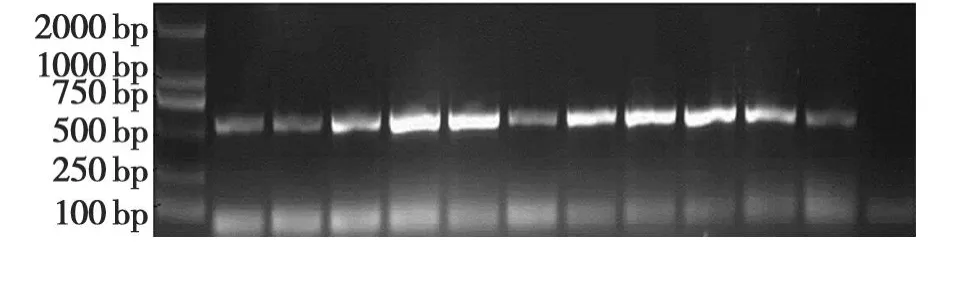
图1 ToMV RT-PCR检测Fig.1 RT-PCR identification of ToMV
2.2 RNAi靶序列的确定
ToMV经分离、纯化后,根据设计的12对引物,利用 RT-PCR从 ToMV cDNA中扩增出CJ2SP1、CJ2SP2、CJ2SP3、CJ2SP4、CJ2SP5、CJ2SP6、CJ2SP7、CJ2SP8、CJ2SP9、 CJ2SP10、CJ2SP11、CJ2SP12共12个片段,回收目的条带并将其连接到 T-Easy Vector中,用 EcoRI单酶切鉴定正确后将样品测序,然后利用测序结果拼接ToMV基因组。
将得到的T oMV序列输入NCBI进行不同株系序列比对,找出保守序列区域,确定RNAi的靶序列。
2.3 pBi35STo5 RNAi载体的构建与鉴定
以CJ2SP1序列为模板,扩增出424 bp大小的干扰片段(图 2)。将干扰片段大小为400 bp的BamHI/KpnI双酶切片段和大小为420 bp的SacI/SpeI双酶切片段分别正向和反向插入到表达载体pBi35SG12相应位点,构建出pBi35STo5 RNAi载体(图3)。pBi35STo5表达载体由BglⅡ单切可得2013 bp和8929 bp条带(图4)。
2.4 pBi35SToBK RNAi载体的构建与鉴定
以CJ2SP9序列为模板,扩增出579 bp大小的干扰片段(图 5)。将干扰片段大小为324 bp的BamHI/KpnI双酶切片段和大小为 530 bp的XbaI/SacI双酶切片段分别反向和正向插入到表达载体pBi35SG12相应位点,构建出 pBi35SToB K RNAi载体(图 3)。
pBi35SToB K表达载体由BamHI/SphI双酶切可得 339、861、1145、1873 和 6766 bp 条带(图 6)。
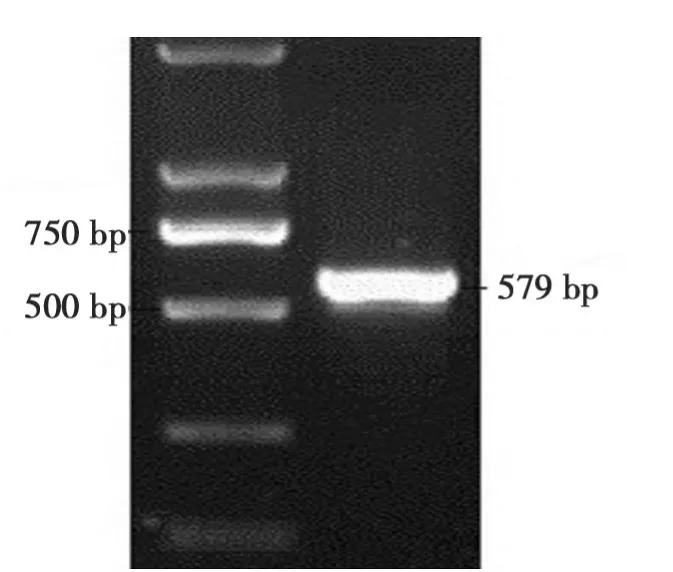
图5 干扰片段的PCR扩增Fig.5 PCR product of RNAi fragment
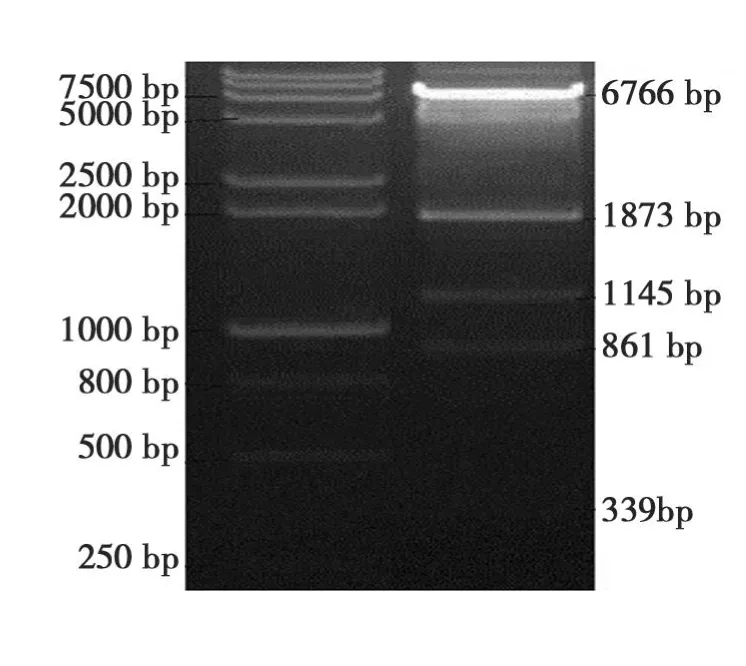
图6 pBi35STo5表达载体酶切鉴定Fig.6 Restriction identification of pBi35SToBK
2.5 pBi35SToMV3 RNAi载体的构建与鉴定
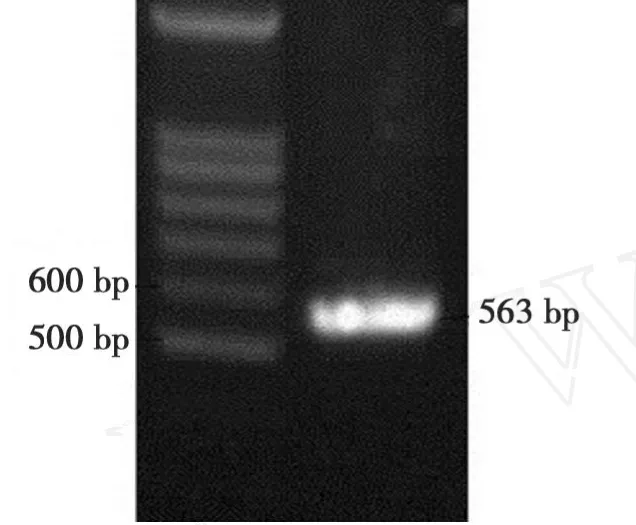
图7 干扰片段的PCR扩增Fig.7 PCR product of RNAi fragment
以CJ2SP12序列为模板,扩增出563 bp大小的片段(图7)。将干扰片段大小为295 bp的 SacI/SpeI双酶切片段和大小为554 bp的BamHI/SalI双酶切片段分别正向和反向插入到表达载体pBi35SG12相应位点,构建出pBi35SToMV3 RNAi载体(图3)。pBi35SToMV3表达载体由 EcoRV单切可得2096、2408和6147 bp片段(图8)。
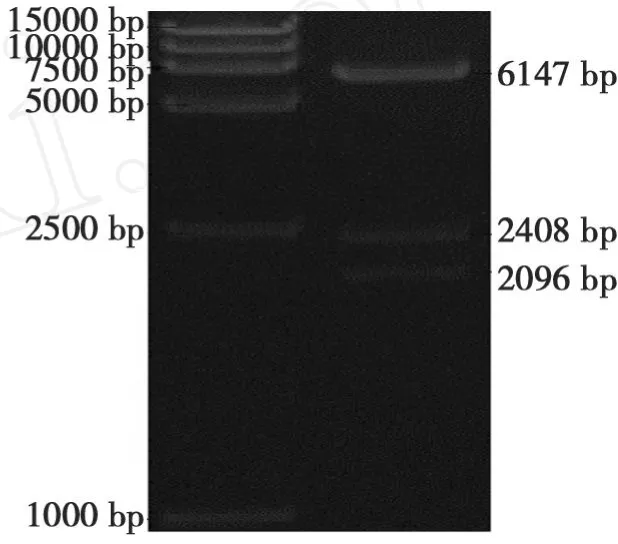
图8 pBi35STo5表达载体酶切鉴定Fig.8 Restriction identification of pBi35SToMV3
2.6 转基因烟草的PCR鉴定
通过电击转化法将RNAi表达载体质粒导入农杆菌 GV3101,菌落 PCR鉴定出阳性克隆,然后利用叶盘法将质粒导人烟草基因组中,得到烟草再生苗。分别提取生根烟草植株叶片基因组DNA进行PCR检测,以构建的RNAi表达载体质粒做阳性对照,以健康普通烟做阴性对照。
转PBi35STo5载体烟草共获得了8个转基因烟草株系(图9),转PBi35SToBK载体烟草共获得了9个转基因烟草株系(图10),转 PBi35SToMV3载体烟草共获得了6个转基因烟草株系(图11),这表明各个干扰基因片段已经成功地整合到烟草基因组中。
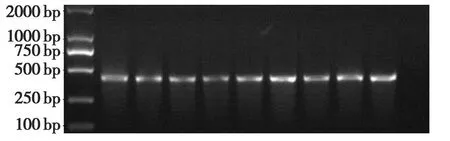
图9 转PBi35STo5载体烟草的PCR鉴定Fig.9 PCR identification of PBi35STo5 in transgenic tobacco plants
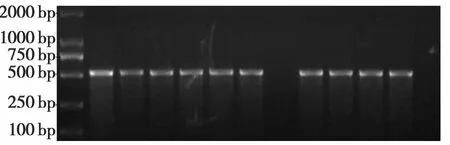
图10 转PBi35SToBK载体烟草的PCR鉴定Fig.10 PCR identification of PBi35SToBKin transgenic tobacco plants

图11 转PBi35SToMV3载体烟草的PCR鉴定Fig.11 PCR identification of PBi35SToMV3 in transgenic tobacco plants
3 讨论
RNAi是由dsRNA介导的、在转录后水平关闭相应序列基因表达的过程,即序列特异性的转录后基因沉默(post-transcriptional gene silencing,PTGS)。dsRNA介导的干扰机制是 Waterhouse等[9]和 Fire等[8]分别在植物和线虫中发现,利用该机制可以将任何目的基因序列插入RNA干涉载体,并获得基因沉默效应。当dsRNA进入细胞后,被一种 RNaseⅢ类的核酸酶Dicer酶[13]切割成21~25 nt的干扰性小RNA(short interfering RNA,siRNA),siRNA进一步与其他多种蛋白成分结合形成诱导的沉默复合体(RNA-induced silencing complex,RISC),最后由RISC介导siRNA反义链与靶mRNA分子互补结合并引起同源性靶mRNA分子的切割效应。由于番茄花叶病毒为单链RNA(single strand RNA,ssRNA)病毒,主要依靠病毒自身编码的RNA依赖的RNA聚合酶进行复制扩增,因此,在复制过程中都有病毒基因的dsRNA形式产生,这就为本实验利用 RNAi技术T oMV研究提供了条件。
在本实验中,pBi35ST o5、pBi35ST oBK、pBi35ST oMV3 RNAi表达载体的干扰序列分别选自ToMV 130/180 kDa的复制酶蛋白、外壳蛋白编码区,其在植物中表达后,沉默外源的 ToMV的同源基因使病毒不能进行正常的复制和表达,从而达到植物抗病的目的。插入的干扰序列转录过程中会形成hpRNA结构,而这种结构有助于进一步提高基因沉默的效率。3个 RNAi表达载体转化普通烟后,可以通过进一步的攻毒试验来筛选抗番茄花叶病毒的高效表达载体,为获得抗 ToMV加工番茄奠定基础。
[1]韩盛,向本春.植物病毒分子检测方法研究进展[J].石河子大学学报:自然科学版,2006,24(5):550-553.
[2]常波,向本春,刘升学,等.加工番茄条斑坏死病原黄瓜花叶病毒外壳蛋白基因的克隆和序列分析[J].石河子大学学报:自然科学版,2006,24(4):410-414.
[3]Kubota K,Tsuda S,T amai A,et al.T omato mosaic virus replication protein suppresses virus-targeted posttranscriptional gene silencing[J].J Virol,2003,77(20):11016-11026.
[4]Ishikawa M,Okada Y.Replication of tobamovirus RNA[J].Proc Jpn Acad Ser B,2004,80(5):215-224.
[5]Powell A P.Delay of disease development in transgenic plants that express the tobacco mosaic virus coat protein gene[J].Science,1986,232(4751):738-743.
[6]Malyshenko S I,Kondakova O A,Nazarova J V,et al.Reduction oftobacco mosaicvirus accumulation in transgenic plants producing nonfunctional viral transport proteins[J].J Gen Virol,1993,74(1993):1149-1156.
[7]Bassble.RNA interference.The short answer[J].Nature,2001,411(6836):428-429.
[8]Fire A,Xu S,Montgomery M K,et al.Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans[J].Nature,1998,391(6669):806-811.
[9]Waterhouse P M,Garham M W,Wang M B,et al.Virus resistance and gene silencing in plants can be induced by simultaneous expression of sense and antisense RNA[J].Proc Natl Acad Sci USA,1998,95(23):13959-13964.
[10]朱俊华,朱常香,温孚江,等.正向和反向重复 RNA介导的抗马铃薯 Y病毒基因工程比较研究[J].植物病理学报,2004,34(2):133-140.
[11]颜培强,李丽杰.应用RNAi技术培育抗2种病毒病的转基因烟草[J].中国生物工程杂志,2007,27(11):27-31.
[12]凌岸瑶.侵染新疆辣椒CMV的外壳蛋白基因的克隆和序列分析[J].石河子大学学报:自然科学版,2008,26(4):431-434.
[13]Bass B L.Double-stranded RNA as a temp late for gene silencing[J].Cell,2000,101:235-238.
Construction of RNAi Vector Which ResistTomato mosaic virus and Transformation of Tobacco
ZHAO Fengyu,ZHENG Yinying,ZHANG Yu,QIAO Yahong,CUI Baiming,XIANG Benchun
(Key Laboratory of Agricultrure Biotechnology/College of Life Sciences,Shihezi University,Shihezi 832003,China)
To obtain transgenic tobacoo which resistTomato mosaic virus,genome sequence of ToMV from Xinjiang processing tomato was amplified by reverse transcription polymerase chain reaction(RT-PCR),then the conserved sequence of the virus among known strains were obtained by using sequence alignment;RNAi sequences were chosen according to conserved regions,and amplified by PCR,pBi35STo5,pBi35SToBK,pBi35ToMV3 three RNAi expression vectors were constructed;through introducing vectors intoNicotiana occidentalisbyA grobacteriummediated transformation separately.The results showed that positive transgenic plants were obtained successfully by molecular identification,which also provides a foundation for further experiment of attacking virus.
processing tomato;RNAi;Tomato mosaic virus
S436.412.19
A
1007-7383(2011)02-0163-06
2010-03-31
国际科技合作项目(2008DFA30560),973计划前期研究专项(2008CB117018)
赵峰玉(1982-),男,硕士生,专业方向为植物基因工程;e-mail:zhaofy82@sina.com。
向本春(1958-),男,教授,博士生导师,从事植物病毒学研究;e-mail:xbc@shzu.edu.cn。
猜你喜欢
中国蔬菜(2016年8期)2017-01-15
浙江中西医结合杂志(2017年2期)2017-01-12
电信科学(2016年11期)2016-11-23
当代化工研究(2016年9期)2016-03-20
中国组织化学与细胞化学杂志(2016年3期)2016-02-27
中国糖料(2015年6期)2015-11-25
中国当代医药(2015年17期)2015-03-01
中国烟草学报(2015年3期)2015-01-25
声屏世界(2014年6期)2014-02-28
西南学林(2013年2期)2013-11-12
