
三蕊柳的组织培养及快速繁殖的研究*
2011-01-06 08:21张云慧姜长阳
哈尔滨师范大学自然科学学报 2011年3期
张云慧,姜长阳
(1.东北师范大学;2.辽宁师范大学)
三蕊柳的组织培养及快速繁殖的研究*
张云慧1,姜长阳2
(1.东北师范大学;2.辽宁师范大学)
以三蕊柳的休眠芽为材料,进行了萌动芽的生长,不定芽的分化,试管苗的继代培养与留茬培养,生根苗扦插和移植的研究,成功地建立起三蕊柳快速繁殖技术.研究结果证明:MS+BA0.1 mg·L-1+GA 1.0 mg·L-1+IAA 0.2 mg·L-1是休眠芽生长培养的理想培养基;1/2MS+BA 0.6 mg·L-1+IAA 0.2 mg·L-1+NAA 0.1mg·L-1是生长芽分化培养的理想培养基;1/2MS+BA 0.4mg·L-1+AgNO30.8 mg·L-1+GA30.5 mg·L-1+IAA 0.3 mg·L-1+NAA 0.1 mg·L-1是不定芽分化继代培养和留茬继代培养的理想培养基;移植到河滩上的试管苗生长旺盛,呈丛生状.
三蕊柳;组织培养;快速繁殖
0 引言
三蕊柳(Salix triandra var.triandra)又称白浆柳,属于杨柳科柳属的灌木或小乔木.分布于我国的辽宁、黑龙江、吉林、内蒙古、河北、山东等地[1-2],生长在海拔500 m 以下的河岸湿地,丛生于砂质土壤上.由于三蕊柳萌枝不仅柔软,而且木质部洁白,适于编织筐、篮等家庭用具和手工艺品,在国际、国内市场上倍受青睐.因此,近年来辽宁南部地区有很多人工栽培的三蕊柳.因三蕊柳的繁殖生物学特点,目前栽培的三蕊柳都是通过枝条扦插的方法进行繁殖的.但由于三蕊柳每年萌生枝条刚刚成熟就被人们全部采收编织筐篮了,种苗问题成为辽宁南部地区三蕊柳栽培的制约瓶颈.为此,对三蕊柳进行了快速繁殖的研究,以期获得大量种苗,满足人们的栽培需要.虽然现在已多有柳树组织培养及无性系建立研究的报道[3-4],但迄今未见三蕊柳组织培养及快速繁殖的研究的报告.该研究以三蕊柳的萌动芽为材料,成功地进行了三蕊柳的组织培养,以很快的繁殖速度繁殖三蕊柳的试管苗,建立起三蕊柳的快速繁殖体系.
1 材料与方法
1.1 材料及来源
以采自瓦房店市老帽山山下的三蕊柳休眠芽为材料.
1.2 材料处理
4月中旬,采集三蕊柳一年生枝条,剪取长4 cm左右有休眠芽的茎段,放入250 mL的广口瓶中,用自来水冲洗20 min左右后,用0.05%安利洗涤剂振荡洗涤4次,再用蒸馏水振荡洗涤3次,接着,转移到超净工作台上,加75%的酒精灭菌20 s左右,迅速用无菌水洗涤2次,再用20%的NaClO溶液振荡灭菌16 min,最后用无菌水振荡洗涤6次.即获得无菌材料.
1.3 培养条件
芽的生长与分化培养基加蔗糖30 g·L-1,生根培养基加蔗糖20 g·L-1,培养基胨力强度为 180 g·cm2-1[5],培养基 pH 为 6.0,培养温度为26℃,休眠芽生长分别进行暗培养,其他培养进行光照培养,光培养其强度为2500 Lx,其时间为 10 h·d-1.
1.4 培养方法
1.4.1 不同生长素对休眠芽生长的影响
在超净工作台上,用解剖刀和解剖针将无菌休眠芽外面的包裹物剥掉后,将0.2 cm左右的休眠芽接种于以 MS+BA 0.1 mg·L-1+GA 1.0 mg·L-1为基本培养基,附加浓度分别为0 mg·L-1、0.2 mg · L-1、0.4 mg · L-1、0.6 mg·L-1和 0.8 mg·L-1的 IBA、IAA、NAA 和2,4-D共17种培养基上,进行休眠芽的生长培养.每种培养基接种100个休眠芽,试验重复了2次.
1.4.2 不同生长素组合对生长芽分化的影响
将上述培养的浅黄色生长芽从培养瓶中取到无菌培养皿中,用解剖刀在距叶柄约0.1 cm处(实际上是在距生长点约0.1 cm处)切下,使其形成具有一个生长点的茎段后,接种到以1/2MS+BA 0.6 mg·L-1为基本培养基,附加不同生长素、不同浓度组合的培养基中,进行生长芽的分化培养.每种处理接种100个生长芽,试验重复了2次.
1.4.3 不同培养基对分化芽继代培养的影响
将分化培养的不定芽从培养瓶中取到无菌培养皿中,用解剖刀在距叶柄约0.1~0.2 cm处切下,使其形成具有2个生长点、长0.5 cm左右的茎段后,分别接种到 1/2MS+BA 0.4 mg·L-1+AgNO30.8 mg·L-1+IAA 0.3 mg·L-1+NAA 0.1 mg·L-1、1/2MS+BA 0.4 mg·L-1+GA 30.5 mg·L-1+IAA 0.3 mg·L-1+NAA 0.1 mg·L-1和1/2MS+BA 0.4 mg·L-1+AgNO30.8 mg·L-1+GA 30.5 mg·L-1+IAA 0.3 mg·L-1+NAA 0.1 mg·L-13种培养基上,进行生长芽的分化继代培养.分化芽继代试验重复了3次,每种处理接种200个材料,每次试验继代培养4代.
1.4.4 丛生生根试管苗的留茬继代培养试验
将分化芽继代培养的丛生生根试管苗上部剪下,继续进行分化芽的继代培养,基部保留两个生长点,使其进行留茬继代培养.丛生生根试管苗的留茬继代培养试验重复了3次.
1.4.5 丛生生根试管苗的扦插试验
把留茬培养的丛生生根试管苗取出,剪成长2 cm左右、至少具有2个生长点的茎段后,剪掉下部的1个叶片,把茎的下部切口放到60 mg·L-1的IAA溶液中处理3~4 min,扦插到上面铺着一层约6 cm厚河沙、下面为肥沃园土的温室苗床上.扦插完后用弥雾的方法使插孔淤闭.扦插后前14 d要保持湿度为95%左右、温度20℃以上、无直射光照的环境条件.14d后按照温室条件进行正常管理.扦插试验重复了3次,每次扦插200个材料.
1.4.6 试管苗的移植
于5月中旬,把扦插成活的试管苗移植到辽宁南部碧流河支流的河滩上.移植试验重复了2次,移植的数量分别为500株和650株.
2 结果及分析
2.1 不同生长素对休眠芽生长的影响
培养15d时可见,在附加不同浓度的NAA和2,4-D培养基上会不同程度地生长出愈伤组织;在附加不同浓度的IBA培养基上培养材料褐化或基本没有变化;附加IAA的培养基上,有的可见培养芽开始生长.培养到50 d时统计证明:在附加浓度为 0.2 mg·L-1的 IAA培养基上,91%的培养芽能生长为长1.6~3.1cm的浅黄色生长芽.这个结果说明:MS+BA 0.1 mg·L-1+GA 1.0 mg·L-1+IAA 0.2 mg·L-1这一培养基是三蕊柳休眠芽生长培养的理想培养基.
2.2 不同生长素组合对生长芽分化的影响
培养45 d观察统计的结果见表1.由表1可以看出,附加不同浓度IBA与不同浓度NAA配合使用的培养基上,不能诱导生长芽分化,而在附加不同浓度IAA与不同浓度NAA配合使用的培养基上,能不同程度地诱导生长芽分化.其中在 IAA 的浓度为 0.2 mg·L-1与浓度为 0.1 mg·L-1的NAA配合使用的培养基上,不仅生长芽的分化率为93%、平均每个培养生长芽分化的不定芽数为5.3,而且分化的不定芽长势好.观察还表明,接种培养10 d后,接种在IAA的浓度为0.2 mg·L-1、NAA 浓度为0.1 mg·L-1的培养基上生长芽基部可见开始分化出不定芽.接着,随着培养时间的延长和分化芽不断地生长,在先分化的不定芽基部,又会陆续分化出多个不定芽.培养到45 d时,每个培养生长芽就会分化形成茎粗0.15 ~0.2 cm、株高 1.0 cm 以上、具有 5~9个叶片,由4~8个不定芽组成的丛生不定芽.这个结果说明:1/2MS+BA 0.6 mg·L-1+IAA 0.2 mg·L-1+NAA 0.1 mg·L-1这种培养基是三蕊柳不定芽分化培养的理想培养基.
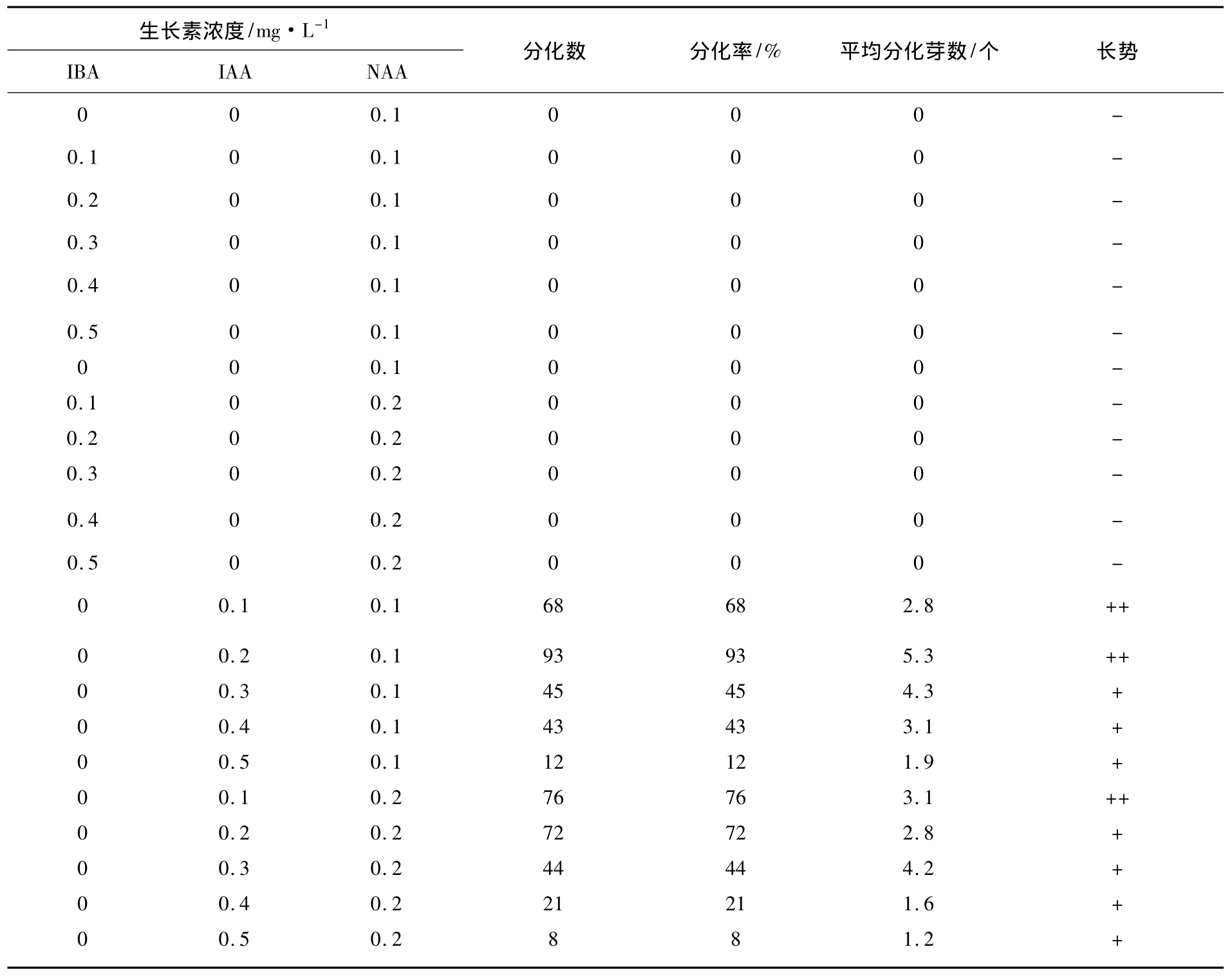
表1 不同浓度生长素对愈伤组织诱导的影响
2.3 不同培养基对分化芽继代培养的影响
培养40 d的统计结果证明,1/2MS+BA 0.4 mg·L-1+AgNO30.8 mg · L-1+GA 30.5 mg·L-1+ IAA 0.3 mg · L-1+NAA 0.1 mg·L-1这种培养基是三蕊柳不定芽分化继代培养的理想培养基.观察统计表明,在上述这种培养基上培养20 d左右,在近培养基表面的茎和叶片上,就会形成数量不等的气生根.气生根长长接触到培养基后,就会成为具有吸收功能的根系.根系一旦形成,不定芽的分化速度和植株的生长速度就会明显加快.培养到40 d时,就会培养成为平均茎粗位0.17 cm、具有6~10个叶片、平均株高3.1 cm、绿色叶片伸展的、由4~10个芽组成的丛生生根试管苗.平均每个培养不定芽能繁殖出6.2个丛生生根试管苗.
2.4 丛生生根试管苗的留茬继代培养试验
试验结果表明,进行1次分化芽的继代培养,可连续进行2代丛生生根试管苗的留茬继代培养.留茬培养的生根试管苗植株粗壮、平均株高为4.3 cm、生长旺盛.留茬培养丛生生根试管苗,平均每代的培养周期为30 d,每代繁殖系数为 4.2.
2.5 丛生生根试管苗的扦插试验
观察表明,扦插半个月左右可见成活并开始生长.3次扦插试验的平均成活率为92.5%.每株试管苗平均可繁殖出2.3个扦插苗.
2.6 试管苗的移植
移植的试管苗成活率为99%.与同期的实生苗相比,移植的试管苗具有植株较大、生长速度快、株型整齐、根系发达等特点.移植成活的试管苗6月中旬长到高30 cm左右时,将上部剪掉,当年秋天就会生长成为长110 cm左右、3~5个可用作编织材料的萌枝.将当年形成的萌枝剪掉,翌年秋天生长出15~25个可用作编制材料的萌枝.所有植株都成丛生状.
3 讨论
在该研究中,采用生长芽分化培养的方法,1个生长芽经过45 d的培养,能繁殖出5.3个不定芽.按照这个速度,每年可繁殖出 5.38.1个后代;采用不定芽分化继代培养的方法,1个不定芽经过40 d的培养,能繁殖出6.2个丛生生根试管苗.按照这个速度,每年可繁殖出6.29.1个后代;采用丛生生根试管留茬继代培养的方法,经过30 d的培养,1株留茬苗能繁殖出4.2个丛生生根试管苗.按照这个速度,每年可繁殖出4.212.2个后代.上述分析说明,在该研究中,不论采用哪种方法进行繁殖,每年都会获得大量的三蕊柳后代,完全可以满足人们栽培对种苗的大量需求.这也说明该研究所获得的技术达到了快速繁殖的目的.但是,由于采用生长芽分化培养的方法所获得的不定芽长势较弱,无效芽的比率较高.因此,生产上应采用不定芽分化继代培养与丛生生根试管留茬继代培养相结合的方法对三蕊柳进行繁殖.用这两种结合的方法繁殖的丛生生根试管苗是采取微型扦插的方法进入苗床的.采取微型扦插方法每株试管苗可繁殖出2.3个扦插苗,这又使繁殖速度增加了1倍多.虽然国内已有微型扦插的报道[6-7],但研究都是在培养瓶中进行的,迄今未见木本植物微型扦插直接进入苗床的报道.本研究使三蕊柳微型扦插直接进入苗床的成功说明,微型扦插手段也可作为试管苗在生产上应用的技术.
在进行分化芽继代培养的试验中,出现了在近培养基表面的茎、叶上形成数量不等气生根,气生根长长接触到培养基后,就会成为具有吸收功能的根系,从而出现了促进了培养芽的分化速度和植株的生长速度的现象.产生这种现象与以下两个原因有关:一方面表明三蕊柳具有形成气生根的遗传性;另一方面与培养芽处在湿度大、温度较高的环境有关.气生根长长接触培养基成为具有吸收功能的根系说明:在人工培养的环境中,三蕊柳试管苗气生根的功能可以发生改变.
[1]李书心.辽宁植物志:上册[M].沈阳:辽宁科学技术出版社,1988.
[2]韩全忠,王正兴.大连地区植物志:上册[M].大连:大连理工大学出版社,1993.102-103.
[3]关亚英,陈英,黄敏仁.簸箕柳组织培养初步研究[J].海南师范大学学报,2009,22(2):204-208.
[4]张天宇,燕丽萍,夏阳,等.柳树愈伤组织的诱导研究[J].山东林业科技,2007,(2):17-19.
[5]姜长阳.培养基琼脂用量的商榷[J].上海:植物生理学通讯,1990,26(2):54.
[6]庞发虎.驱蚊草微型扦插快速繁殖的研究[J].安徽农学通报,2009,15(1):68-69.
[7]李秋杰,张吉,陈春燕.月季微型扦插快速繁殖研究[J].安徽农业科学,2007,35(32):10254,10274.
Study of Tissue Culture and Rapid Propagation of Salix Triandra var.Triandra
Zhang Yunhui1,Jiang Changyang2
(1.Northease Normal University;2.Liaoning Normal University)
In this paper,the growing and differentiation of adventitious bud,subculture of test- tube plant,cattaging and transplanting of rooted seedling are studied by using dormant bud of salix triandra var.triandra.The results showed that MS+BA 0.1 mg·L-1+GA 1.0 mg·L-1+IAA 0.2 mg·L-1is suitable for the growth of dormant bud;1/2MS+BA 0.6mg·L-1+IAA 0.2 mg·L-1+NAA 0.1 mg·L-1is suitable for differentiation of the bud;1/2MS+BA 0.4 mg·L-1+AgNO30.8 mg·L-1+GA30.5 mg·L-1+IAA 0.3 mg·L-1+NAA 0.1mg·L-1is suitable for subculture of the adrentitious bud.The seedling grew well and thickly on the flood land by transplanting.
Salix triandra var.triandra;Tissue Culture;Rapid propagation
2011-03-15
*辽宁省教育厅科学技术研究项目资助(2008353)
(责任编辑:季春阳)
猜你喜欢
散文诗(2021年22期)2022-01-12
河北农机(2020年10期)2020-12-14
江河文学(2020年6期)2020-01-04
东坡赤壁诗词(2018年3期)2018-07-16
消费导刊(2018年8期)2018-05-25
好孩子画报(2016年7期)2016-12-12
浙江农业科学(2016年11期)2016-05-04
山东青年(2016年1期)2016-02-28
中小学实验与装备(2014年2期)2014-09-21
读书(2014年8期)2014-01-19
