
番茄早疫病菌抗嘧菌酯突变体培养特性研究
2010-09-11 04:29周杰敏任璐韩巨才刘慧平王文忠
山西农业大学学报(自然科学版) 2010年5期
周杰敏,任璐,韩巨才,刘慧平,王文忠
(1.山西农业大学大学 农学院,山西太谷 030801;2.山西省平顺县北社乡人民政府,山西平顺 047400)
番茄早疫病Alternaria solani是由茄链格孢引起的番茄重要病害之一,在我国多省都有发生,对番茄产量影响很大。由于番茄早疫病具有潜育期短、再侵染频繁、流行速率高的特点,长期使用单一药剂防治容易形成抗药性群体[1]。嘧菌酯azoxystrobin是甲氧基丙烯酸酯类杀菌剂,其杀菌活性高、杀菌谱广、内吸性强、对非耙标作物和从未使用过嘧菌酯等甲氧基丙烯酸酯类杀菌剂的作物安全且与环境有良好的相容性[2~4]。本文对番茄早疫病菌的抗、感菌株进行了营养条件、pH及温度的一系列生物学性状比较,以了解抗、感菌株之间的生物学特性差异。
1 材料与方法
1.1 供试菌株
番茄早疫病菌亲本敏感菌株WZ42采集于山西省五寨地区,经筛选获得。
番茄早疫病菌抗药突变体WZ42-8,是将亲本敏感菌株经过紫外光诱导及药剂驯化而获得。
1.2 供试药剂及培养基
药剂:25%嘧菌酯(azoxystrobin)悬浮剂(先正达作物保护有限公司)。
培养基:PDA培养基:马铃薯200 g、葡萄糖20 g、琼脂20 g、蒸馏水1000 mL。理查德培养基:硫酸镁(MgSO4·7H 2O)2.50 g;氯化铁(FeCl3)0.02 g;硝酸钾10.00 g;磷酸二氢钾5.00 g;蔗糖50.00 g;蒸馏水 1000 mL。
1.3 不同营养条件对抗性突变菌株菌丝生长的影响
供试碳源:葡萄糖、蔗糖、可溶性淀粉、麦芽糖、乳糖;
供试氮源:蛋白胨、牛肉膏、尿素、缺氮处理。
基本培养基为理查德(Richard)培养基[5],不同碳源相当于理查德培养基中50 g蔗糖含碳量置换其中的蔗糖,不同氮源相当于理查德培养基中10 g KNO3含氮量置换其中的KNO3,将含有不同碳源、氮源的培养基分别制成平板,每处理3皿。将抗、感菌株的4 mm菌饼接种于各处理上,在25℃恒温箱中培养,分别在48 h后测量菌落直径[6],计算菌丝生长速度。
1.4 酸碱度对菌丝生长的影响
用0.1 mol·L-1的 HCl和0.1 mol·L-1的NaOH无菌操作,供试PDA培养基分别调整酸碱度为 p H 3、4、5、6、7、8、9、10、11 、12,每一酸度重复3次,每皿放一片4 mm的菌碟,25℃恒温培养,每隔24 h测一次菌落直径,计算菌丝生长速度[7]。
1.5 温度对菌丝生长的影响
用4 mm打孔器切取菌碟,无菌操作下移入直径为9 cm的PDA平板中,每皿一碟,每处理重复3次,分别置于6个不同处理温度的恒温箱、冰箱中培养,每隔24 h观察一次菌落直径,计算菌丝生长速度[8]。
2 结果与分析
2.1 不同营养条件对抗性突变菌株菌丝生长的影响
结果(表1)表明各抗性突变体与其亲本敏感菌株对碳源和氮源的利用情况有所不同:亲本菌株WZ42以在可溶性淀粉为碳源的培养基中菌丝生长最快,生长速度为0.4375 mm·h-1,而抗性菌株WZ42-8在以乳糖为碳源的培养基中生长最快(其速率为0.3073 mm·h-1);亲本菌株WZ42在以蛋白胨为氮源的培养基中菌丝生长最快(0.4583 mm·h-1),抗性菌株WZ42-8在以牛肉膏为氮源的培养基中菌丝生长最快(0.2951 mm·h-1)。抗性菌株在不同营养源下生长速率均低于其敏感亲本。其中在蔗糖、可溶性淀粉及蛋白胨培养基中抗、感菌株之间生长速率在5%显著水平及1%极显著水平上有明显差异。
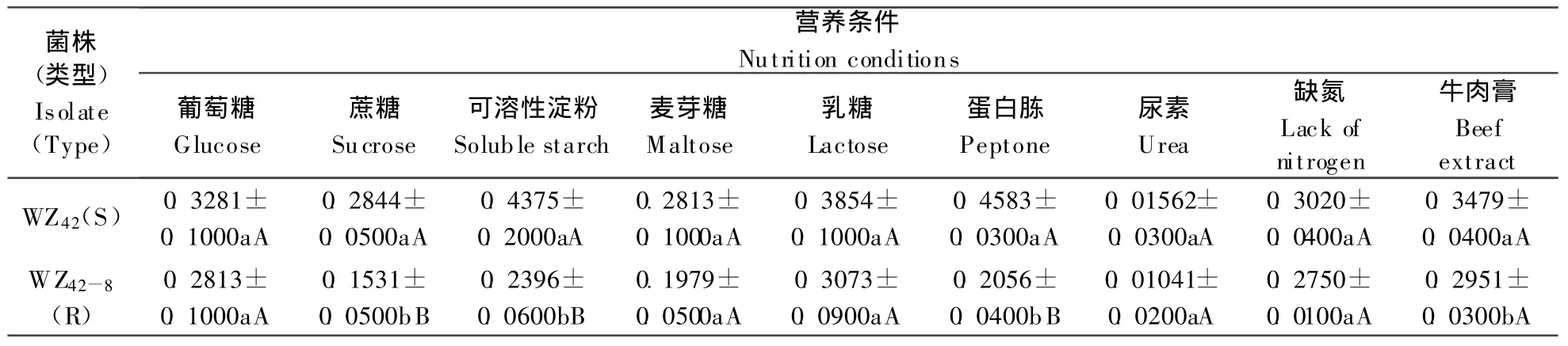
表1 不同营养条件下亲本菌株和抗药菌株菌丝的生长速率/mm·h-1Table1 The growing rate of isolates in different nutrition conditions/mm·h-1
2.2 酸碱度对菌丝生长的影响
研究结果(表2)表明,抗、感菌株在 p H 4~10范围内能生长,p H 5~7最适合菌丝生长,过酸或过碱的环境均不利于菌丝生长,而在p H>8的条件下亲本菌株与抗性突变体随着p H的升高菌株生长的速度迅速减慢,在不同pH值条件下,亲本菌株比突变体生长速度快。由表2还可以看出,在最适p H值范围内,亲本菌株在p H 6时生长最快,其菌丝生长速率为0.4467~0.6167 mm·h-1,而突变体在 p H=7时生长最快,菌丝生长速率为0.3041~0.4111 mm·h-1,可见二者生长最适pH值有所不同。在 48 h内,抗、感菌株在p H 5、6、7、8、9差异显著。

表2 不同p H值下亲本菌株和抗药菌株菌丝的生长速率/mm·h-1Table2 The growing rate of isolates in different p H value conditions/mm·h-1
2.3 温度对菌丝生长的影响
结果见表3,从生长速率来看,抗、感菌株的菌丝的生长速率均较稳定,从温度处理来看,4℃时菌丝基本不能生长,以20~25℃生长最快。在4~25℃之间随温度升高,两类菌丝生长速度逐渐加快,在25~35℃范围内随温度升高,菌丝生长速率逐渐下降,35℃几乎不生长。在25℃下,随时间的增加抗、感菌株之间的方差分析差异逐渐减少,在120 h时差异不显著。

表3 不同温度下亲本菌株和抗药菌株菌丝的生长速率(mm·h-1)Table3 The growing rate of isolates in different temperature conditions(mm·h-1)
3 结论与讨论
通过研究番茄早疫病菌对嘧菌酯抗、感菌株生物学特性,可以得出:敏感亲本在以可溶性淀粉为碳源和以蛋白胨为氮源的培养基中菌丝生长最快,而抗性突变体在以葡萄糖为碳源和以牛肉膏为氮源的培养基中生长最快;抗性突变体在p H值为7时生长最快而亲本菌株在pH值为6时生长最快;虽然不同温度下抗、感菌株生长情况基本相似,但敏感菌株生长速率均快于抗性突变体。不同的营养条件、p H值、温度对突变体菌丝生长均有影响,且不同营养条件及p H下,抗、感菌株之间变化明显,说明突变体的生理特性出现了变异,生长发育的内源节律发生了变化。
番茄早疫病长期以来一直以化学防治为主,使得番茄早疫病菌在一定程度上对几种常用杀菌剂的敏感性降低[9]。尽管目前在山西省大田中尚未检测到抗嘧菌酯的番茄早疫病菌,但已有关于番茄早疫病抗性菌株的报道[10],因此即使是嘧菌酯这类敏感性较高的药剂在生产实践中也应该慎重使用。本研究主要对番茄早疫病菌抗嘧菌酯突变体和敏感菌进行了一些初步的生物活性测定,在以后的研究工作中应进一步比较抗性突变体和敏感菌株在分子生物学和生理生化上的差异,从而对番茄早疫病菌对嘧菌酯抗性风险评估及抗性治理提供解决方案。
[1]陈利锋,徐敬.农业植物病理学(南方本)[M].北京:中国农业出版社,2001:314-317.
[2]张舒亚,周明国.甲氧基丙烯酸酯类杀菌剂的生物学及应用技术研究[M].中国植物病害化学防治研究,第三卷.北京:中国农业科技出版社,2002:1-10.
[3]DALES.Efficacy of'Amistar'against Fruit and Vegetable Diseases in Asia[M].中国植物病害化学防治研究,第三卷.北京:中国农业科技出版社 ,2002:42-49.
[4]Gullnoml,Lerouxp,Smithcm.Usesand Challenges of Novel Compounds for Plant Disease Control[J].Crop Protection,2000,19(2):1-11.
[5]方中达.植物病研究方法[M].中国农业出版社,1996:39-40.
[6]倪守延,杨文翠,张成林,等.油菜菌核病营养生理研究[J].安徽农业大学学报,1995,22(3):246-250.
[7]史建荣,王裕中,沈素文,等.pH值对油菜菌核病菌生长的影响[J].江苏农业科学,1999(3):47-49.
[8]Choulwar,A.B.and Datar,V.V.Physiological studies onAlternar ia solanicausing early blight of tomato[J].Journal of Maharashtra Agricultural Universities,1991,16(2):265-266.
[9]王洪俊.50%异菌脲可湿性粉剂防治番茄早疫病药效试验[J].现代农药.2003(6):40-41.
[10]张洪,刘慧平,韩巨才,等.番茄早疫病对杀菌剂的敏感性研究[J].山西农业大学学报:自然科学版,2006,26(1):36-37.
猜你喜欢
陕西农业科学(2022年2期)2022-03-28
浙江农业学报(2021年5期)2021-05-31
长江蔬菜(2021年12期)2021-04-04
中国果业信息(2019年11期)2019-01-05
生命科学研究(2018年1期)2018-05-29
上海农业学报(2017年4期)2017-04-10
安徽医科大学学报(2016年12期)2017-01-15
广东饲料(2016年5期)2016-12-01
西南农业学报(2016年5期)2016-05-17
天津医科大学学报(2015年2期)2015-12-22
