
两种棕榈藤光合日变化及其与环境因子的关系研究
2010-08-21 02:52官凤英范少辉刘亚迪彭颖邓旺华
世界竹藤通讯 2010年4期
官凤英 范少辉 刘亚迪 彭颖 邓旺华
(国际竹藤网络中心 竹藤科学与技术重点实验室生物资源利用科学研究院 北京 100102)
两种棕榈藤光合日变化及其与环境因子的关系研究
官凤英 范少辉*刘亚迪 彭颖 邓旺华
(国际竹藤网络中心 竹藤科学与技术重点实验室生物资源利用科学研究院 北京 100102)
利用LI-6400光合仪测定了盈江省藤和小省藤的光合特征参数,探讨了两种棕榈藤光合日变化规律,并结合环境因子分析影响净光合速率(Pn)的主要因素,研究结果表明:(1) 盈江省藤净光合速率Pn和蒸腾速率Tr日变化曲线均呈“双峰”型,存在“午休”现象;小省藤Pn值低,波动小,日变化呈“双峰”曲线趋势,Tr日变化曲线呈“单峰”型。 (2)小省藤光合“午休”时间是12:00,盈江省藤的光合“午休”时间在下午14:00,比小省藤推迟了2h。(3) 盈江省藤Pn日变化与RH呈显著正相关(r=0.820*,P<0.05),RH和Gs下降是导致盈江省藤发生光合“午休”的决定因素。小省藤的光合能力较盈江省藤弱,Pn日变化与各环境因子无显著相关性。
棕榈藤;净光合速率;环境因子;日变化;光合“午休”
棕榈藤(rattan)是棕榈科(Palmae)省藤亚科(Calamoideae)省藤族(Calameae)中13个属600余种植物的统称(李荣生,2004),是热带和亚热带森林中多用途植物资源,主要分布于亚洲地区,华南地区是棕榈藤分布中心的北缘(江泽慧等,2002)。近年来,不少学者在棕榈藤的繁殖(Umail-Garcia, 1985;Yin & Xu,1992)、种子储藏条件(尹光天等,1992)、种植造林技术(许煌灿等,1994)、藤材特性以及原藤加工利用技术(李旸和腰希申,2002;Wahab et al, 2004;江泽慧等,2007b)、藤苗生长条件等方面开展了较深入研究,但关于棕榈藤光合特性及其与环境因子之间关系的研究还不够系统,仅有少量关于光的形态建成(Aminuddin, 1985)、部分棕榈藤水分利用效率和抗旱能力(冯昌林等,1999;李荣生,2003)等方面的报道。作为决定植物生产力高低的主要因素(Buchanan et al, 2000),植物的光合性能与其自身的遗传性(邓仲篪,1994)和所处的环境条件密切相关(郑国生和邹琦,1993)。因此,本文通过对盈江省藤(Calamus nambariensis var. yingjiangensis)和小省藤(Calamus graeilisRoxb.) 苗期光合作用及环境因子关系进行研究,揭示棕榈藤苗光合生理特性的日变化规律,探索不同棕榈藤对环境因子的适应性,为科学培育棕榈藤苗木提供依据。
1 材料与方法
1.1 供试材料
盈江省藤(Calamus nambariensisvar.yingjiangensis)和小省藤(Calamus graeilisRoxb.) 均采于云南省盈江县铜壁关,是优良的材用藤种。2007年2月初,置于瓦缸内催芽,4月初将芽苗移栽到营养钵,放到日光温室内培养,苗木上方用50%的遮阳网遮阴,9月初进行光合测定。
1.2 光合作用日变化测定
每个藤种选生长条件一致的待测苗木3株做3次重复,选择苗木上部功能完全的叶片作为测定对象,每株测3片叶子,结果取多次测定的平均值。选择晴好天气从8:00-18:00每隔2h用LI-6400便携式光合作用分析仪(Li-cor, USA)测定叶片的净光合速率Pn,(μmol CO2/m2•s1)、蒸腾速率Tr(mmol H2O/m2•s1)、气孔导度Gs(cm/s1) 、胞间CO2浓度Ci,(μmol/mol1)等因子以及大气CO2浓度Ca,(μmol/mol1)、大气相对湿度RH(%)、大气温度Ta(℃)、光合有效辐射PAR(μmol/m2•s1)、饱和蒸汽压差VPD等环境因子。测定时,叶室条件与温室环境保持一致,不作任何设定或调节。
1.3 数据处理
试验数据采用Excel进行统计处理,均采用各测量指标的平均值。数据采用SPSS 13.0软件系统进行相关性分析和方差分析。
2 结果与分析
2.1 环境条件日变化
测定结果显示(图1),在8:00-18:00期间,2种棕榈藤苗各环境因子的日变化趋势基本一致,只有图1-A中所示的小省藤所处大气CO2浓度(Ca)在16:00以后有上升的趋势,而盈江省藤保持稳定。图1-A所示二者的Ca的日变化均呈明显的“双峰”曲线,分别在10:00和14:00达到最高峰,在12:00达到最低值,盈江省藤和小省藤环境CO2浓度分别为405.954μmol/mol1、409.963μmol/mol1;而光合有效辐射(PAR)呈先升高后降低的“单峰”曲线,在12:00时达到最高峰。图1-B中所示的大气相对湿度(RH)和大气温度(Ta)则呈相反的变化趋势,前者先下降又上升,后者先上升后下降,均在14:00达到最低值和最高值,这符合环境因子的变化规律。饱和蒸汽压差(VPD)的变化趋势(图1-C)是先上升后下降,在14:00达到高峰值,盈江省藤和小省藤的VPD分别为4.697Kpa、4.178Kpa。

2.2 净光合速率的日变化
2种棕榈藤苗叶片的净光合速率日变化结果显示(图2),日变化基本上呈不规则的“双峰”曲线,二者均在10:00达到第1个峰值,盈江省藤和小省藤的净光合率分别为2.492μmol/m2•s1、1.545μmol/m2•s1。对于小省藤来说,在14:00达到第2个峰值1.346μmol/m2•s1,2h后峰值基本上保持平稳,到18:00时Pn才开始有下降的趋势,前后2个峰值相差不大。对于盈江省藤,在14:00达到最低峰0.725μmol/m2•s1,出现“午休”现象,之后开始上升,到18:00时仍没有下降的趋势,第1个峰值高于第2个峰值,这是由于高光照强度、高温的条件提高了叶片的光呼吸强度,增加了呼吸消耗,导致净光合速率下降(万素梅等,2009)。对于小省藤,在正午12:00时的PAR达到当天的最大值(图1-A),超过了藤类的光饱和强度,叶片周围空气之间的VPD(图1-C)上升,空气湿度下降(图1-B),再加上此时Ca达到全天的最低值(图1-A),在一定程度上促进呼吸,光合作用的底物减少,从而降低植物的光合作用。而在光照强度日变化一致的情况下,盈江省藤的光合午休比小省藤推迟了2h,直到VPD达到最高值时才表现出“午休”现象, 说明盈江省藤对光照强度不敏感。经过分析(图1所示), 2种藤类的环境因子之间没有显着差异,但是二者的Pn的日变化有显着差异。此外,如图1-A 所示,小省藤的Ca日变化在18:00时上升,而其Pn 并没有随着反应底物的变化而变化,反而下降,可以说明此时的变化与PAR有关。

2.3 蒸腾速率的日变化
2种藤类的蒸腾速率(Tr)日变化趋势如图3所示,盈江省藤呈现出“双峰”趋势,存在“午休”现象,在12:00时达到最高峰0.862mmol/m2•s1,在16:00时达到次高峰0.592 mmol/m2•s1;小省藤蒸腾速率日变化呈现“单峰”曲线,在14:00时达到最高值0.774mmol/m2•s1。从蒸腾速率日变化来看,盈江省藤在12:00时的高蒸腾速率值可以缓和此时的较高的光照强度对它的影响,因此,Pn没有在高光强时出现“午休”现象,而Pn和Tr均在14:00时发生“午休”现象,这有可能是由于此时盈江省藤的气孔关闭所致。而对于小省藤来说,此时的Tr最高,也有可能是持续高温迫使水分失衡,增加蒸发散。
2.4 气孔导度的日变化

气孔导度(Gs)反映了气孔开度的大小。光合作用所需要的水分和CO2首先必须经过叶片的气孔才能进入叶肉细胞。因此,气孔的关闭,即气孔导度的大小对植物的光合作用起着重要的调节作用(王会肖和刘昌明,2003)。图4显示,对于不同的藤类,Gs的日变化表现出不同的变化趋势。对于盈江省藤来说,在早上8:00时气孔导度最大,但由于PAR较低,Pn的值也不高,随着时间的推移,Gs降低Pn却上升(图2),Ca也上升(图1-A),此时有可能是与光合作用有关的酶等非气孔因素在起作用(冯建灿和张玉洁,2002),并使Pn值10:00时达到一个高峰值,随后Pn值下降,在14:00时表现出光合“午休”现象,此时Gs也达到最低值,而小省藤整体的Gs值均变化比较平缓,对Pn的影响相对较小,但在12:00时达到相对最低值,与光合“午休”时间一致。可见气孔因素是光合作用的限制因素,在日变化的不同阶段起着不同作用,在光合“午休”时均起决定作用。
2.5 胞间CO2浓度与大气CO2浓度之比的日变化
大气CO2浓度(Ca)的变化会直接影响到胞间CO2浓度(Ci)的变化。以Ci/Ca代表叶片内CO2的动态变化,叶片气孔下腔细胞间CO2供应浓度变化将反映气孔导度问题。有研究显示,考察Pn和Ci/Ca的变化可以判断Pn的变化是否由气孔因素调节,当Ci/Ca与Pn的变化方向相同时,可以认为Pn的下降是由于气孔导度降低引起的;如果在Pn下降时Ci/Ca保持稳定或上升,则可判断Pn的下降是非气孔因素引起的(黄丛林和张大鹏,1996;薛仿正等,2006)。由图5可见,2种藤类的Ci/Ca的日变化趋势趋于平缓,并不随时间的推移呈明显的高低起伏的变化,只在10:00时有降低的趋势,达到一个相对较低值,并且2种藤类之间并不存在明显差异,而图2中显示的Pn的变化方向与Ci/Ca不同,这说明Pn的日变化并非全由气孔因素所致。
2.6 不同棕榈藤苗各光合特征参数的相关性分析

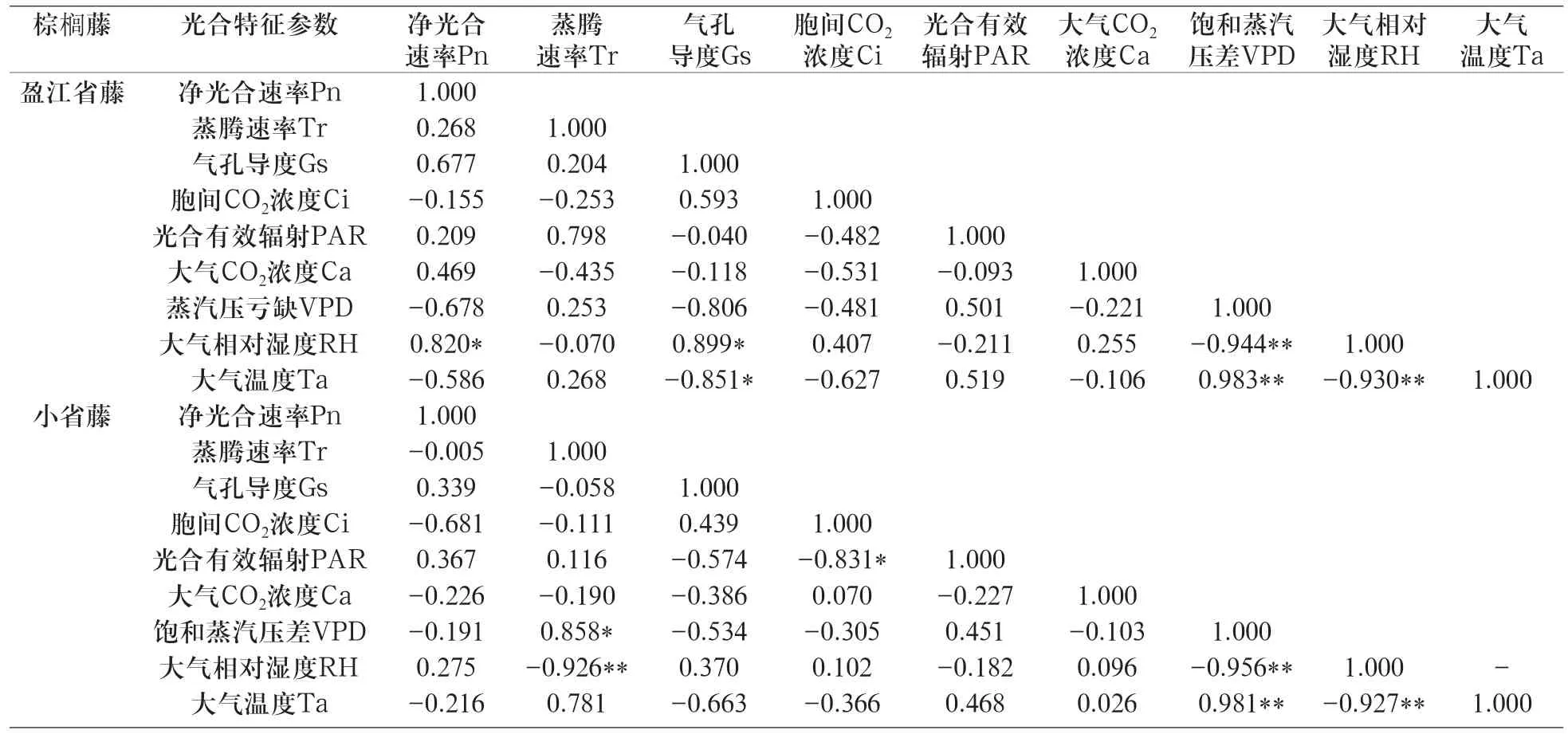
表1 不同棕榈藤苗各光合特征参数与环境因子的相关性分析Table1 Correlation analysis between photosynthetic characteristic parameters and environmental factors for seedlings of different rattans
光合作用的日变化随着植物种类和环境条件的变化而有所不同。在供试的各种藤类植物中,以其各光合特征参数以及相关环境因子的日变化值进行相关分析(表1)。结果表明,盈江省藤Pn日变化与RH呈显著正相关(r=0.820*,P<0.05),与其它环境因子相关性不显著。RH与Gs呈显著正相关(r=0.899*)、与VPD(r=-0.944**,P<0.01)、Ta(r=-0.930**)呈极显著负相关。Ta与Gs呈显著负相关(r=-0.851*),与RH呈极显著负相关(r=-0.930**),与VPD呈极显著正相关(r=0.983**)。从相关系数的大小来看,对Pn影响较大的因子还有Gs、VPD,对Tr影响较大的因子是PAR。
小省藤Pn日变化与各环境因子相关性不显著,相关系数也不大,由图2也能看出小省藤Pn的日变化幅度相对较小。Tr日变化与RH呈极显著负相关(r=-0.926*),与VPD呈显著正相关(r=0.858*),从相关系数的大小来看,对Tr影响较大的因子还有Ta。VPD与RH呈极显著负相关 (r=-0.956**),与Ta呈极显著正相关(r=0.981**)。可见,环境因子即VPD对小省藤的蒸腾作用有显著作用,而光合速率受环境因子的影响较小。
3 讨论
植物净光合速率在一天中的动态变化是叶片光合能力与环境条件综合作用的结果,在不同的环境条件下,引起光合作用动态变化的原因也不同(蔚荣海等,2009);在相同的环境条件下,不同植物表现出的光合作用的动态变化的原因也不同(孔蓓蓓等,2009)。关于植物光合作用与各环境因子之间的关系,研究者之间针对不同植物持不同的观点。万素梅等(2009)人研究得出苜蓿净光合速率与光合有效辐射、气孔导度呈极显著正相关,与田间CO2浓度呈极显著负相关;王瑞等(2009)认为净光合速率与光照强度、大气CO2浓度呈正相关,与胞间CO2浓度呈负相关。本研究显示,2种棕榈藤的各环境因子之间没有显著性差异。在环境条件一致的情况下,与光合作用有关的各项参数日变化随着棕榈藤种类的不同而存在差异,初步分析表明各环境因子在藤类光合参数日变化的不同阶段起不同作用。
盈江省藤Pn日变化曲线存在“午休”现象;而小省藤Pn日变化波动较小,且值都不高,可以说明小省藤的光合能力较前者低。盈江省藤Pn的最低峰比小省藤推迟了2h,在12:00的光强高峰时并没有出现光合“午休”,而在VPD高峰时才显现出来,并且在18:00时Pn仍没有降低的趋势,而小省藤随着光照强度的降低,Pn值也开始下降,这是否能说明盈江省藤是一种耐强光的藤类仍需进一步分析。盈江省藤发生光合“午休”时的Ca值较高,Tr值较低,而有研究认为大气CO2浓度升高增加光合作用的底物,同时会抑制呼吸,减少蒸腾,从而提高植物光合作用(Ceulenman,1994),这与之本文的结果不符,可以说明盈江省藤的光合“午休”现象与Ca无关;而气温较高会引起空气湿度下降、叶片周围空气的饱和蒸汽压差上升,从而导致叶片失水、气孔导度下降,进而抑制光合作用的正常进行(何文兴等,2004),这与本研究的结果相符,因此可以推测出RH和Gs下降是导致盈江省藤发生光合“午休”的决定因素,这也与日变化整个阶段相关性分析中(表1)的结果一致。而从日变化整个阶段来说,相关性分析表明,Pn的日变化与Ca呈显著相关,可见光合“午休”阶段与日变化的整个阶段,起决定作用的环境因子不同。小省藤Gs值与Pn值的日变化趋势均较平缓,可以推断出Gs对其光合速率的变化起决定作用,同时相关性分析得出,小省藤的Pn日变化与各环境因子均无显著相关性,至于小省藤Pn日变化是否与其叶片气孔自身组织结构有关仍需进一步研究。
1 李荣生.中国棕榈藤植物名录[J].世界竹藤通讯,2004,2(4):8-10.
2 李旸,腰希申.棕榈藤的电镜观察[J].林业科学,2002,38(1):173-174.
3 江泽慧,范少辉,张昌顺,官凤英.棕榈藤研究进展[J].江西农业大学学报,2007,29(6):957-964,1005.
4 江泽慧,吕文华,费本华,任海青,吴玉章,王朝晖.3种华南商用藤材的解剖特性[J].林业科学,2007,43(1):121-126.
5 江泽慧,萧江华,许煌灿.世界竹藤[M].沈阳:沈阳科技出版社,2002,509-612
6 王会肖,刘昌明.作物光合,蒸腾与水分高效利用的试验研究[J].应用生态学报,2003,14(10):1632-1636.
7 Aminuddin M. Performances of some rattan trial plots in Peninsular Malaysia[M]//Wong K M,Manokaran N. (Eds.). Proceedings of the Rattan Seminar, Kuala Lumpur, Malaysia: RIC, 1985:49-56
8 许煌灿,尹光天,曾炳山等.黄藤栽培技术研究[J].林业科学研究,1994,7(2):239-246.
9 冯昌林,张伟良,许煌灿,尹光天,范晋渝,张万幸.白藤苗木生长与水分条件的初步研究[J].广西科学院学报,1999,15(2):94-96.
10邓仲篪.水稻光合日变化与内生节律的关系[J].中国水稻科学,1994,8(1):9-14.
11冯建灿,张玉洁.喜树光合速率日变化及其影响因子的研究[J].林业科学,2002,38(4):34-39.
12何文兴,易津,李洪梅.根茎禾草乳熟期净光合速率日变化的比较研究[J].应用生态学报,2004,15(2):205-209.
1 3黄丛林,张大鹏.葡萄叶片光合速率日间降低内外因调控的研究[J].园艺学报,1996,23(2):128-132.
14孔蓓蓓,刘超,尹伟伦,夏新莉.沙柳、黄柳和杞柳光合作用的日变化[J].河南科技大学学报:自然科学版,2009,30(3):79-83.
15李荣生.华南地区3个棕榈藤种水分利用效率和抗旱能力的研究[D].北京:中国林业科学研究院,2003.
16万素梅,贾志宽,杨宝平.苜蓿光合速率日变化及其与环境因子的关系[J].草地学报,2009,17(1):27-31
17王瑞,陈永忠,王湘南,彭邵锋,杨小胡,王玉娟,陈隆升.油茶优良无性系光合作用的日变化.中国农学通报[J].2009,25(24):236-239.
18蔚荣海,赵颖君,徐克章,张美善,张治安,陈展宇.不同生境条件下人参、西洋参光合作用的日变化[J].华南农业大学学报,2009,30(4):7-11.
19薛仿正,温鹏飞,柴凯,王鹏飞,杜俊杰.设施栽培欧李光合特性的研究[J].安徽农业科学,2006,34(23):6100-6101.
20郑国生,邹琦.不同天气条件下田间大豆光合作用日变化的研究[J].中国农业科学,1993,26(1):44-50.
21Buchanan B B, Gruissen W, Jones R L.Biochemistry and molecular biology of plants[J]. Rockville: The American Society of Plant Physiologist, 2000, 260-310.
22Ceulenman S R, Mousseau M. Effects of elevated atmospheric CO2on woody plants [J].New Phytologist, 1994, 127: 425-446.
23Umail-Garcia M. Tissue culture of some rattan species[C]//Proceedings of the rattan seminar,Singapore: IPGPI, 1985: 23-31.
24Yin G T, Xu H C. Effect of fertilizers on seedlings ofDaemonorops margaritae[J]. Rattan Information Center Bulletin,1992,11(1): 9-12.
A Study of Diurnal Variations in Photosynthetic Activity and Its Relationship with Environmental Factors for Two Species of Rattans
Guan Fengying, Fan Shaohui*, Liu Yadi, Peng ying, Deng Wanghua
(International Centre of Bamboo and Rattan, The Key Laboratory for Bamboo and Rattan, Academy of Bioresources Utilization, Beijing 100102)
In the study, the diurnal dynamics of photosynthetic activity inCalamus nambariensisvar.yingjiangensis andCalamus graeilisRoxb. were monitored using portable LI-6400 photosynthesis system in combination with the analysis of the main environmental factors inf l uencing the net photosynthetic rate. The results were summarized as follows: 1)Calamus nambariensisvar. yingjiangensis had an obvious bimodal curve of diurnal variation both in Pn and Tr with obvious midday depression of photosynthesis whileCalamus graeilisRoxb.had small fl uctuations in the bimodal curve of diurnal variation for Pn and unimodal curve for Tr. 2) The midday depression of photosynthesis inCalamus nambariensisvar. yingjiangensis was later for two hours than that ofCalamus graeilisRoxb.. 3) There was a signif i cantly positive correlation between the diurnal variation in Pn and RH (r=0.820*,P<0.05). The decrease in RH and Gs was the determinant resulting in the midday depression forCalamus nambariensis var. yingjiangensis. There was no signif i cant correlation between the diurnal variation in Pn and environmental impact factors forCalamus graeilisRoxb. which had lower photosynthetic capacity thanCalamus nambariensis var. yingjiangensis.
rattan, net photosynthetic rate, environmental factor, diurnal variation, midday depression of photosynthesis
* 资助项目:林业科学技术推广项目“棕榈藤种质资源培育及利用技术推广与示范”([2005]86);国际竹藤网络中心科研专项“优质棕榈藤苗期生长及光合作用研究”(06/07-B18)。
官凤英(1974-),女,吉林人,博士。主要从事竹藤资源培育、经营与管理技术的研究。
* 通讯作者:博士,研究员,首席专家。
猜你喜欢
云南画报(2021年6期)2021-07-28
小天使·一年级语数英综合(2020年11期)2020-12-16
当代工人(2020年11期)2020-07-06
Asian Herpetological Research(2019年1期)2019-03-27
文理导航·科普童话(2017年4期)2018-02-10
散文诗(2017年17期)2018-01-31
风景园林(2017年3期)2017-06-22
作文大王·中高年级(2016年11期)2016-12-09
中国糖料(2016年1期)2016-12-01
学生天地(2016年24期)2016-05-17
