
普通白菜高Zn胁迫耐性及Zn积累特性研究
2010-05-21 14:50陈秀灵徐艳如崔秀敏吴小宾
中国蔬菜 2010年14期
陈秀灵 徐艳如 崔秀敏 吴小宾
(山东农业大学资源与环境学院,山东 泰安 271018)
Zn是兼具营养与毒害的重金属之一,作为营养元素,它是植物体内碳酸酐酶、碱性磷酸酶等多种酶的必需组分,促进植物的蛋白质代谢,参与光合作用,提高植物抗逆性(陆景陵,2003)。植物体内正常Zn含量为20~100mg·kg-1,一般植物体内Zn含量达到100mg·kg-1时就表现过量,含量为400mg·kg-1时则会造成严重毒害(汪洪和金继运,2009),大量Zn在根部积累,使根系伸长受到抑制,造成生长不良,甚至死亡(Godbold et al.,1983;保琼莉 等,2007)。Zn的毒害表现为抑制光合作用、减少CO2固定、影响韧皮部运输、改变细胞膜渗透性,导致生长减缓、受阻和失绿(保琼莉 等,2007)。但是,也有相关报道某些植物可在严重Zn污染地区生长,其体内Zn含量可达1000mg·kg-1,具有超富集Zn的能力,它们大多是十字花科类植物,遏蓝菜(Thlaspi caerulescens)中Zn含量可达51600mg·kg-1,是普通草本植物的100倍以上(韩文轩 等,2009)。
锌矿开采、冶炼、镀锌加工等含锌工业三废的不合理排放,使土壤中 Zn超常积累,污染土壤中的 Zn易被植物吸收而造成毒害,并可通过食物链污染农产品,威胁人类健康。因此,近年来有关如何消除重金属对蔬菜等危害的研究颇受关注(王晓芳和罗立强,2009)。普通白菜〔Brassica campestris L.ssp.chinensis(L.)Makino var.communis Tsen et Lee〕又称青菜、小白菜,属十字花科芸薹属类蔬菜,原产中国,是南北方普遍种植的叶菜,具有奢侈吸收 Zn的特性。本试验将对其富集锌的机制及食用安全性作初步探讨。
1 材料与方法
1.1 供试材料与方法
试验于2009年4~5月在山东农业大学温室内进行。供试普通白菜品种为苏州青、黄心乌、瓢儿菜、抗热 605、南农矮脚黄,由南京农业大学普通白菜课题组提供。采用砂培法,盆钵上口径19 cm,下口径12 cm,盆高17 cm。1 mol·L-1HCl浸泡石英砂24 h,蒸馏水冲洗数次后晾干装盆,每盆2 kg。Zn(以ZnSO4·7H2O为Zn源)设5个水平:0.43(CK)、100、200、400和800mg·L-1,3次重复。浇灌霍格兰-阿农营养液。
种子先用50g·L-1次氯酸钠消毒20min,蒸馏水反复冲洗,浸种12 h,置于培养箱中25 ℃下催芽,露白后播于盆钵中。营养液用蒸馏水配制,pH用1 mol·L-1KOH或HCl调至6.0±0.1。出苗后浇灌1/4浓度的营养液,7 d后改为1/2营养液,4 d后换为全营养液。4月3日定株,每盆10株,14 d后幼苗3~4片真叶时,每两天浇灌1次150mL Zn处理液,共浇灌10次,最后各处理浇灌的Zn用量分别为0.615、75.29、150.29、300.29、600.29 mg·kg-1,处理20d后收获。
1.2 项目测定
植株收获后按处理将单株混合后测定。将其分为根系、叶柄和叶片3部分。
植株生长量的测定:先用直尺测量根系长度,然后将根系、叶柄和叶片分装于信封中,105 ℃下杀青30min,并在80℃下烘干至恒质量,称其干质量,测定生物量,计算根系耐性指数(严明理 等,2009)
Zn含量测定:干样磨碎后过筛,HNO3-HClO4消煮后采用原子吸收分光光度法测定Zn含量(鲁如坤,2000)。
1.3 数据处理
数据采用DPS软件进行统计分析,采用Microsoft Office Excel软件作图。
2 结果与分析
2.1 供Zn水平对普通白菜生长的影响
从表1可以看出,5个普通白菜品种地上部均没有明显的Zn中毒症状,各供Zn浓度对叶柄和叶片生物量的影响均较小。根系生物量均随供Zn浓度的提高先升后降,抗热605在供Zn浓度为100mg·L-1时最高,南农矮脚黄和苏州青在200mg·L-1时最高,而瓢儿菜和黄心乌在400mg·L-1时才达最大值。黄心乌、瓢儿菜、南农矮脚黄根系生物量在供Zn浓度为100~800mg·L-1之间增幅范围分别为25.00%~42.18 %、12.77 %~53.19 %、0~46.51 %。与对照相比,供Zn浓度为800mg·L-1时,苏州青和抗热605根系生物量降幅分别为11.21 %、29.17 %,抗热605降幅较大;黄心乌和瓢儿菜增幅分别为26.56 %、12.77 %,黄心乌增幅较大;而对南农矮脚黄无影响。同一供Zn水平下瓢儿菜的根系生物量最低。
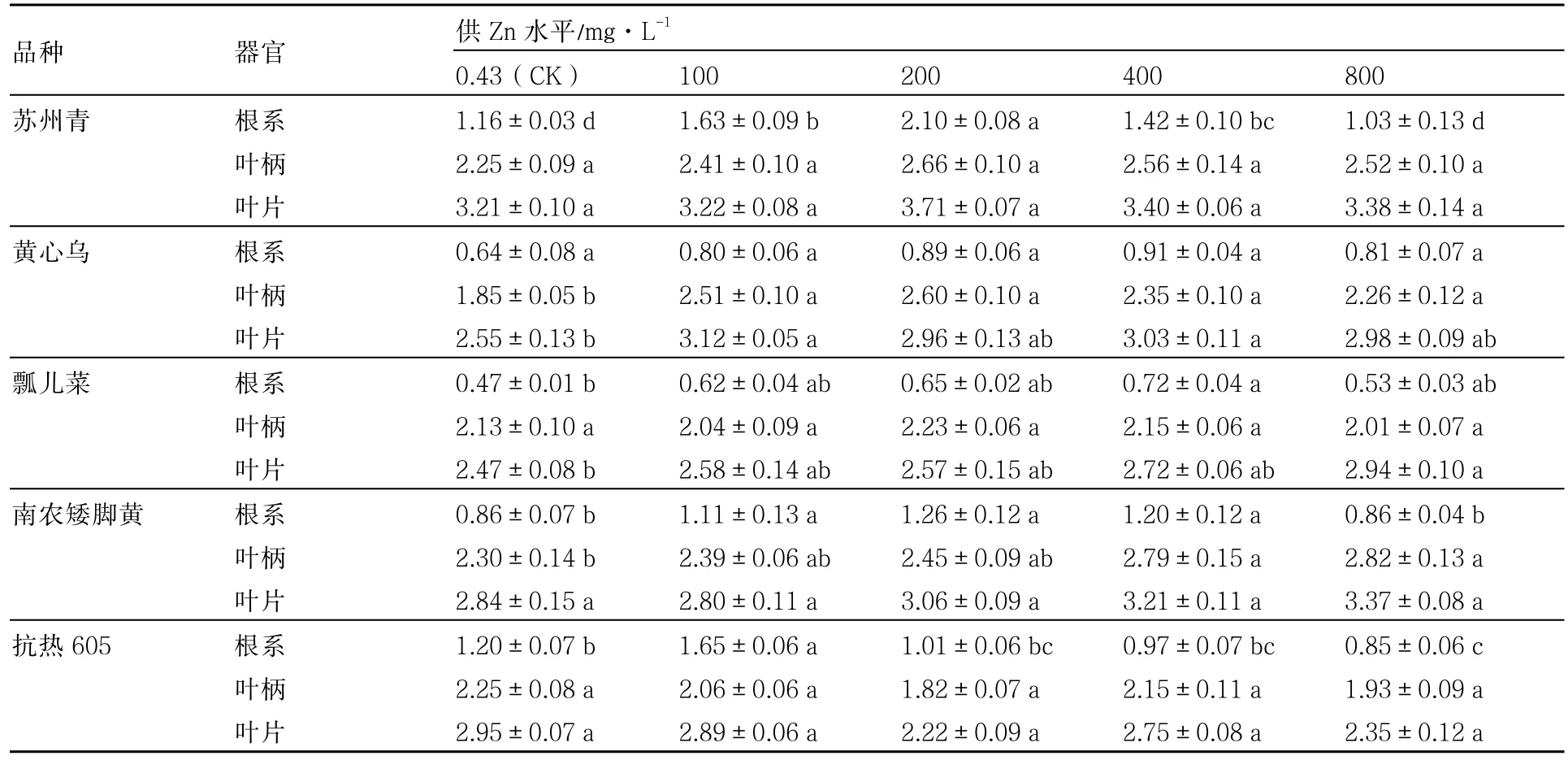
表1 不同供Zn水平下普通白菜各器官的生物量 g·盆-1
2.2 供Zn水平对普通白菜根系耐性指数的影响
根系耐性指数(RTI)是指重金属处理的根长与对照根长的比值(严明理 等,2009)。植物与重金属作用时,根系细胞壁中的大量交换位点能将重金属离子吸收或固定,从而促进或阻止重金属离子进一步向地上部转运。因此,根系耐性指数能够反映植物对重金属的耐性(Monni et al.,2001)。从表2可知,除抗热605外,不同供Zn水平下4个普通白菜品种的根系耐性指数均先升高后降低,且均大于 0.5。黄心乌、瓢儿菜和南农矮脚黄均>1,说明浇灌 100~800mg·L-1的 Zn处理液对根系无明显的抑制作用;苏州青仅在高浓度(800mg·L-1)Zn处理下略小于1,抗热605在200mg·L-1Zn处理下小于1,且与100mg·L-1Zn处理差异显著,由此可见,抗热605根系对高浓度的Zn较敏感。
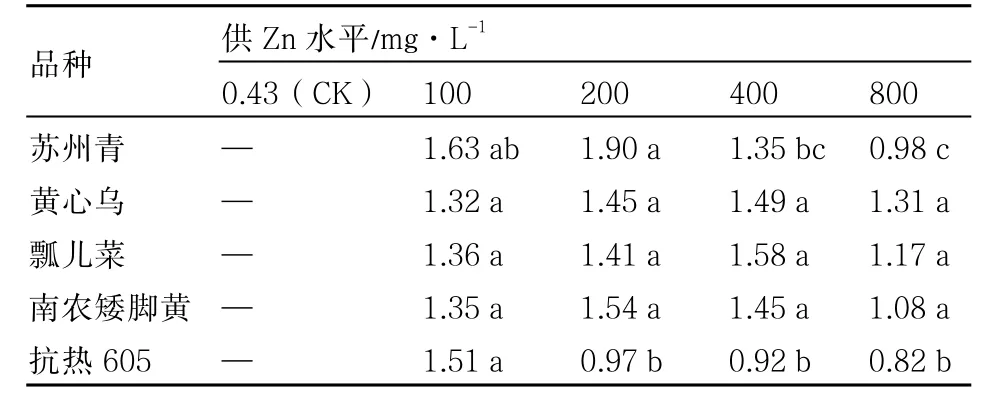
表2 不同供Zn水平下普通白菜的根系耐性指数
2.3 供Zn水平对普通白菜Zn吸收及积累的影响
从表3可以看出,5个普通白菜品种根系、叶柄、叶片中的Zn含量均随着供Zn浓度的升高而大幅增加。除南农矮脚黄外,其他品种根系中Zn含量均可达1000mg·kg-1左右。100mg·L-1Zn处理下根系Zn含量与对照相比,最大极差为瓢儿菜(393.28 mg·kg-1),最小极差为南农矮脚黄(2.97 mg·kg-1)。
从表3还可以看出,南农矮脚黄各器官Zn含量明显低于其他品种。不同器官中,同一供Zn水平下南农矮脚黄叶柄Zn含量最低,在较低供Zn水平(0.43~200mg·L-1)下南农矮脚黄根系Zn含量最高,而在较高供Zn水平(400~800mg·L-1)下则是叶片Zn含量最高。其他品种同一供Zn水平下不同器官中Zn含量均是根系最高,叶柄和叶片相差不大。
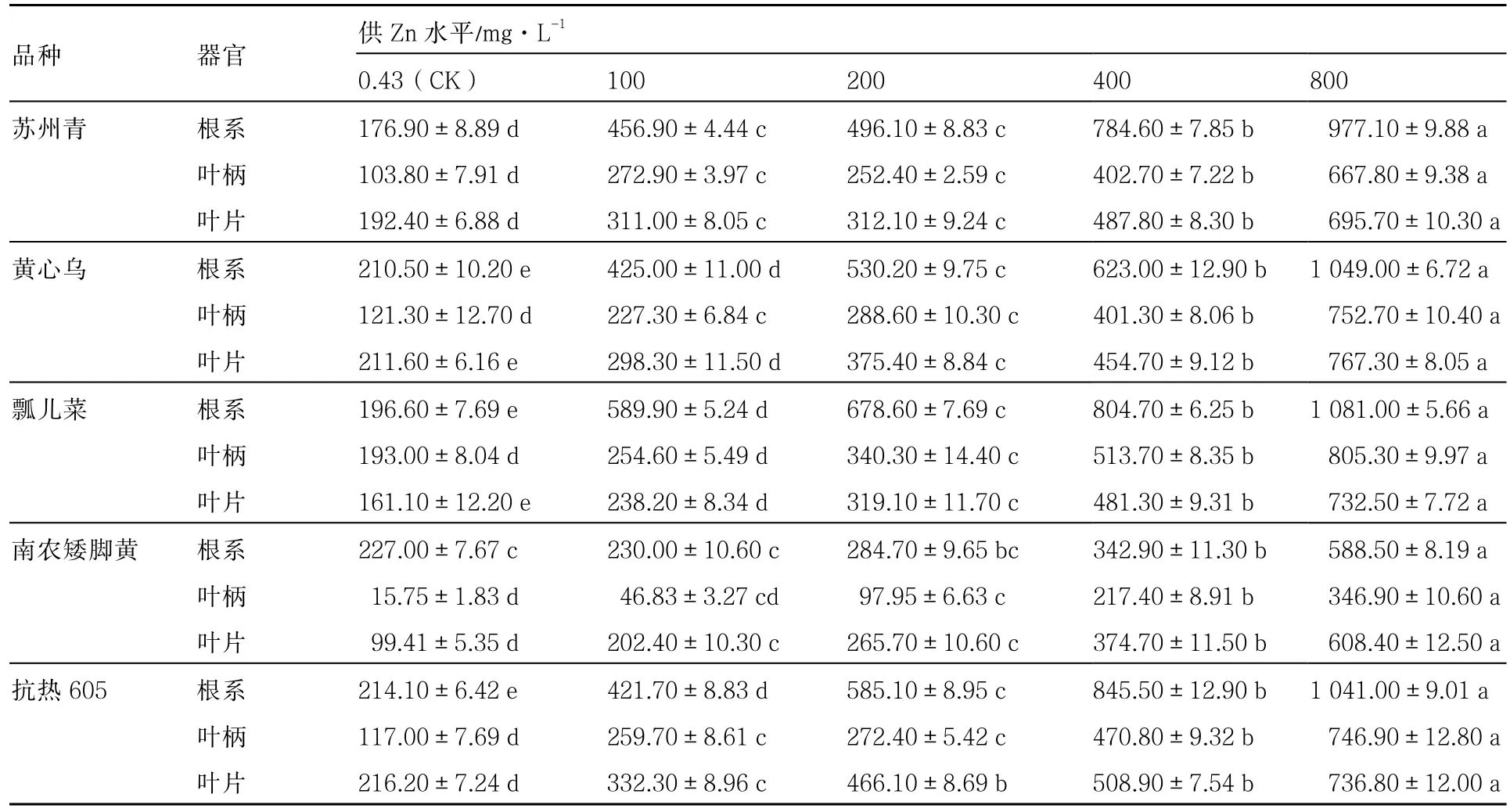
表3 不同供Zn水平下普通白菜根系、叶柄、叶片中Zn含量 mg·kg-1
从表4可以看出,普通白菜各器官Zn积累量与供Zn浓度的变化趋势相同,处理之间差异较大。800mg·L-1Zn处理下,南农矮脚黄根系Zn积累量最低,苏州青最高,其次为抗热605、黄心乌、瓢儿菜,可见不同品种之间存在一定的差异。普通白菜地上部Zn积累量在32~410μg·株-1之间,显著高于根系Zn积累量。
2.4 供Zn水平对普通白菜Zn分配的影响
从表5看出,随供Zn浓度的提高,苏州青、黄心乌、瓢儿菜、抗热605的根系Zn分配比例先升高后降低;南农矮脚黄根系 Zn分配比例急剧下降,叶柄则相反,叶片中变化不明显。各品种叶片Zn分配比例明显高于叶柄,均在40%以上。Zn在普通白菜地上部的分布存在品种间差异,南农矮脚黄、抗热 605、苏州青、黄心乌叶柄 Zn分配比例逐渐增加,最高增幅比例分别是20.65 %、12.50%、11.74 %、10.21 %。瓢儿菜叶柄Zn分配比例随供Zn浓度提高则呈下降的趋势,叶片的分配比例有所增加。说明南农矮脚黄、苏州青、黄心乌、抗热 605地上部富集的 Zn主要贮存在叶柄中,而瓢儿菜富集的 Zn更多贮存在叶片中;南农矮脚黄地上部较其他品种更易富集大量的Zn。供试的5个普通白菜中,南农矮脚黄转运Zn的能力最强。
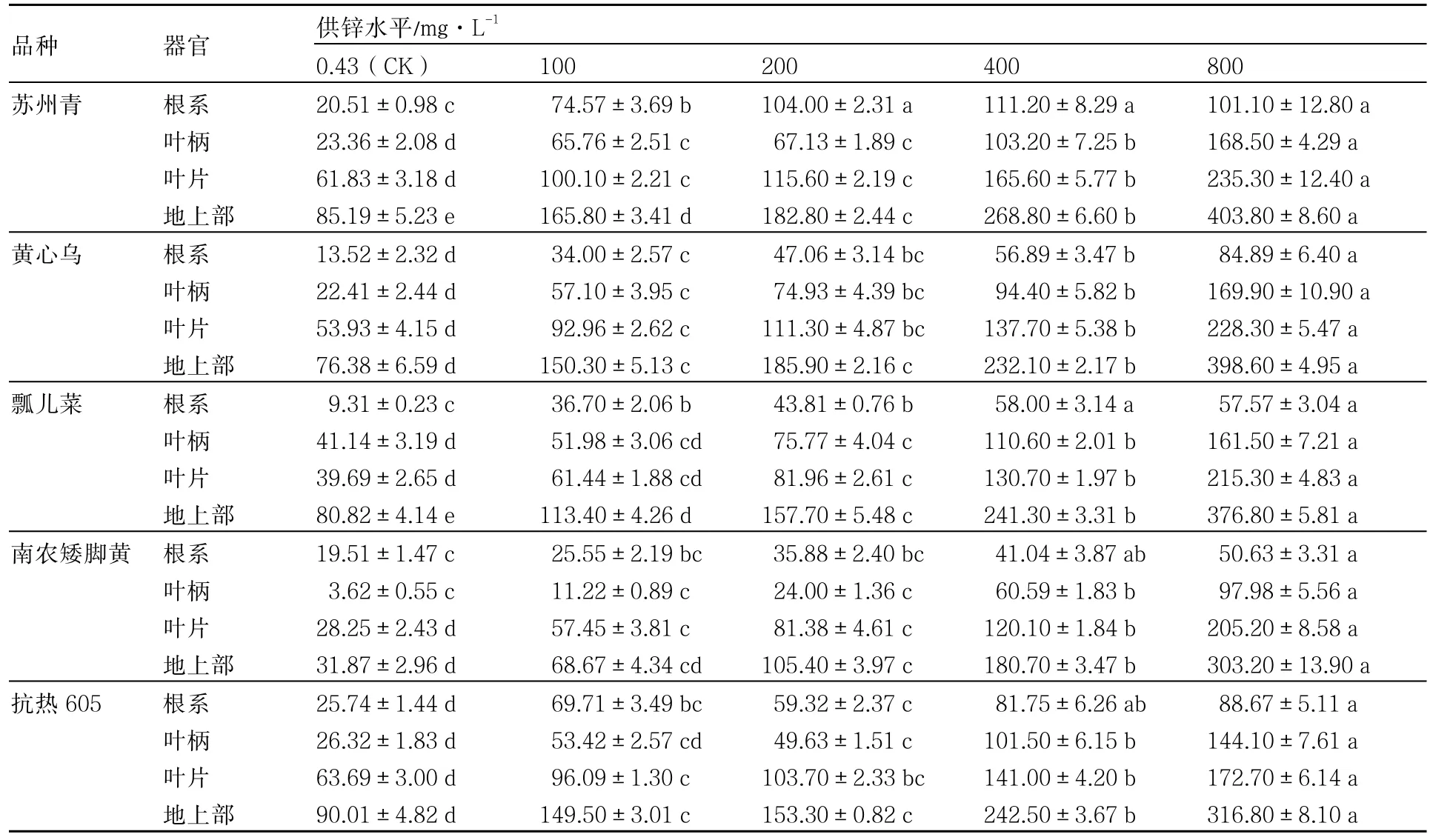
表4 不同供Zn水平下普通白菜的Zn积累量 μg·株-1
苏州青和黄心乌根系 Zn分配比例在供Zn浓度为200mg·L-1时达最大值,瓢儿菜和抗热 605在供 Zn浓度为 100mg·L-1时就达到最大值,这可能与根系对 Zn的容纳存在某个饱和点有关(汤叶涛等,2009),且饱和点的高低与基因型有关。除南农矮脚黄外,其他品种地上部的 Zn分配比例变化趋势与根系相反,呈U型变化。地上部Zn分配比例在60%以上,根系Zn分配比例在40%以下。可见,在较高浓度(400~800mg·L-1)Zn处理下,当根系吸收的Zn超过饱和点后,无法再承受大量吸收的Zn,则通过将 Zn不断向地上部转运来避免对根系的伤害。
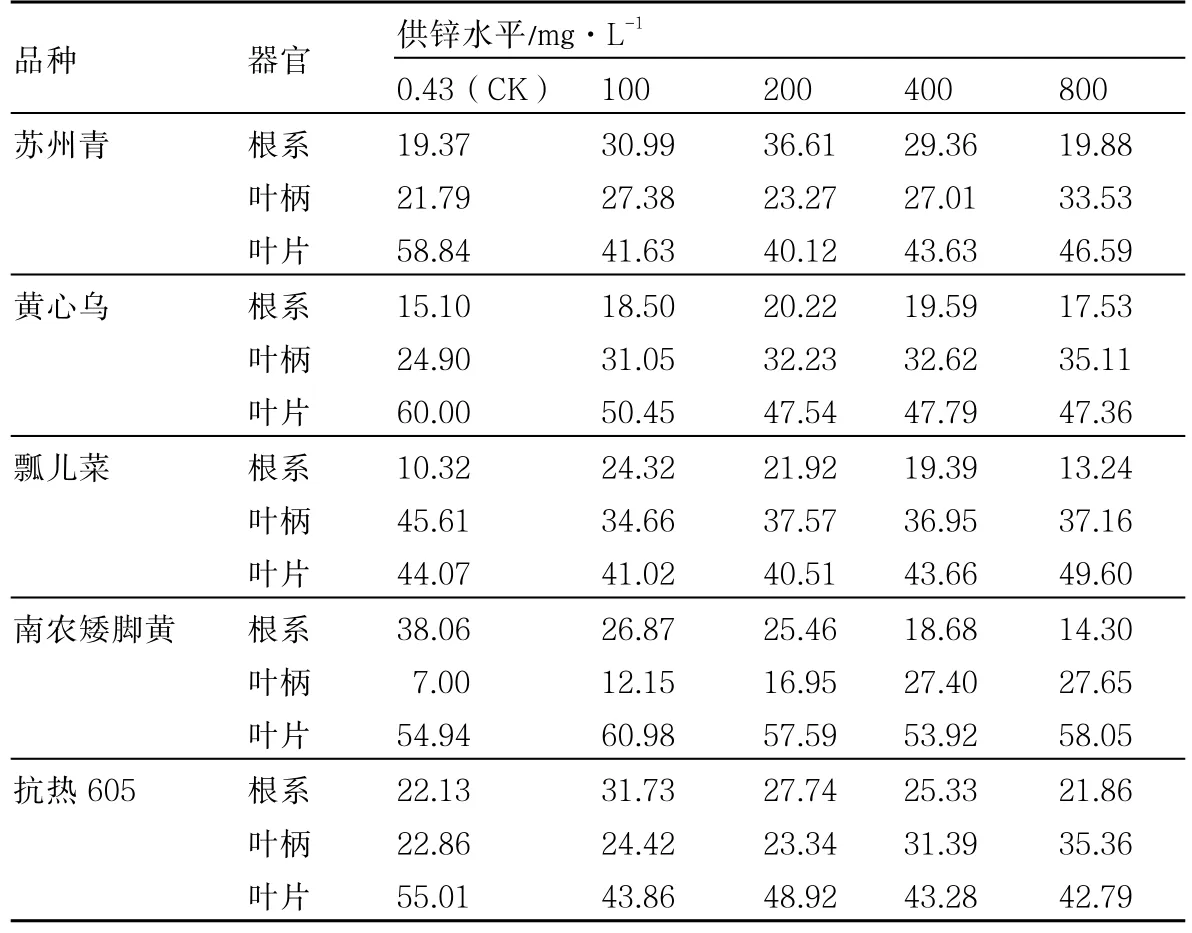
表5 不同供Zn水平下普通白菜Zn的分配比例 %
3 结论与讨论
3.1 普通白菜Zn耐性评价
据有关研究报道(Bert et al.,2000;Escarre et al.,2000;Wu et al.,2007),根系和地上部的相对生长状况可以很好地被用来评价芸薹属类植物对过量Zn的耐性。敏感植物在中毒浓度下,根系生长被抑制,植物生长缓慢,生物量减少,而耐性植物则没有影响或影响较小。本试验5个普通白菜品种在供试 Zn水平下均未出现毒害症状,对地上部的生物量影响较少,甚至有一定的促进作用。从耐性指数指标来看,指数越大表示植物对重金属的耐性越大,耐性指数大于0.5时,表明这种植物对此重金属有较强的耐受性,生长得较好。当耐性指数小于0.5时,则说明重金属对这种植物的毒害作用明显,这种植物基本难以或不能生长在这种浓度的重金属环境中(徐卫红等,2005)。本试验中5个供Zn水平处理下,5个普通白菜品种的根系耐性指数在0.82~1.90之间,可见高Zn胁迫对根系伸长的抑制效应较小,这可能与过量Zn对芸薹属植物根系生长的影响更多体现在对侧根密度的抑制上,而不是对最大根长的抑制有关(Ebbs & Kochian,1997)。因此,综合生物量和根系耐性指数推断:普通白菜对Zn存在较高的耐性。
在800mg·L-1供Zn水平下,5个普通白菜品种中抗热605的根系生物量降幅最大,同时根系耐性指数较其他基因型低,因此,抗热605耐性相对较弱。
3.2 普通白菜的Zn积累与分布
重金属超富集植物这一概念最初是由 Brooks等(1977)提出的,当时用以定义地上部富集Ni超过1000mg·kg-1的植物。一般认为超富集植物应同时满足以下3个标准(Brooks et al.,1998,Reeves & Baker,2000):① 植物地上部富集的某种元素含量达到生长在同一介质非超富集植物地上部含量的100倍以上,其临界含量标准(以干质量计)是Zn 10000mg·kg-1;② 地上部重金属含量大于根部含量;③ 植物的生长没有受到明显的伤害。理想的超富集植物还应具有生长期短、抗病虫能力强、地上部生物量大、能同时富集两种或两种以上重金属的特点。虽然本试验中5个普通白菜品种在供Zn浓度为800mg·L-1时,根系Zn含量比对照提高近20倍,达1000mg·kg-1,远远超过了植物体内正常Zn含量25~100mg·kg-1的范围(陆景陵,2003)。但Zn含量均未达到超富集植物的临界标准,且地上部小于根系,因此,不具有超富集植物的基本特征。
植物对重金属的吸收对策主要有两种:一种是植物排斥对策,另一种是富集耐受对策。在富集耐受对策中,一种为限制吸收运输对策,即根部富集截留而限制重金属向地上部运输,另一种为富集或超富集对策,即能将大部分重金属运送到地上部(柯文山 等,2004)。本试验5个普通白菜品种地上部的Zn积累分配比例均在60%以上,而根中分配比例不足40%。可见,普通白菜吸收的Zn大部分转移到地上部,叶柄和叶片可被认为是植株体内Zn的临时贮备库,属于富集耐受对策。普通白菜Zn积累及其分布存在品种间差异,南农矮脚黄根系Zn积累量最低,但其转运能力最强,苏州青根系 Zn积累量最高,转运能力也较高。徐卫红等(2006)研究表明,大白菜在Zn胁迫下分泌的有机酸和氨基酸种类和含量不同是造成品种间Zn吸收差异的重要原因。普通白菜对Zn的富集耐受机制可能与根系分泌物、Zn的赋存形态以及Zn在细胞、亚细胞结构上的区室化分布有关,这正在进一步研究中。
保琼莉,田霄鸿,杨习文,李生秀.2007.不同供Zn量对三种小麦基因型幼苗生长和养分吸收的影响.植物营养与肥料学报,13(5):816-823.
韩文轩,徐一鸣,杜威,唐傲寒,江荣风.2009.ICP-AES分析不同锌浓度培养下锌镉超累积植物遏蓝菜的元素计量变化.光谱学与光谱分析,29(9):2565-2567.
柯文山,陈建军,黄邦全,戴灵鹏,王万贤.2004.十字花科芸薹属 5种植物对 Pb的吸收和富集.湖北大学学报:自然科学版,26(3):236-269.
陆景陵.2003.植物营养学·上册.北京:中国农业大学出版社:95-98.
鲁如坤.2000.土壤农化分析方法.北京:中国农业科技出版社:325-336.
汤叶涛,吴妤都,仇荣亮,曾晓雯,胡鹏杰.2009.滇苦菜(Picris divaricata Vant.)对Zn的吸收和富集特性.生态学报,29(4):1823-1831.
汪洪,金继运.2009.植物对锌吸收运输及积累的生理与分子机制.植物营养与肥料学报,15(1):225-235.
王晓芳,罗立强.2009.铅锌银矿区蔬菜中重金属吸收特征及分布规律.生态环境学报,18(1):143-148.
徐卫红,熊治庭,王宏信,李仰锐,刘吉振,李文一.2005.Zn胁迫对重金属富集植物黑麦草养分吸收和Zn积累的影响.水土保持学报,19(4):32-43.
徐卫红,刘吉振,黄河,熊治庭.2006.高Zn胁迫下不同大普通白菜品种生长、Zn吸收及根系分泌物的研究.中国农学通报,22(8):458-463.
严明理,刘丽莉,王海华,向言词,冯涛.2009.3种植物对红壤中镉的富集特性研究.农业环境科学学报,28(1):72-77.
Bert V,Macnair M R,de Laguerie P,Saumitou-Laprade P,Petit D.2000.Zinc tolerance and accumulation in metallicolous and nonmetallicolous populations of Arabidopsis halleri(Brassicaceae).New Phytol,146:225-233.
Brooks R R,Lee J,Reeves R D,Jaffre T.1977.Detection of nickeliferous rocks by analysis of herbarium specimens of indicator plants.J Geochem Explor,7:49-57.
Brooks R R,Chambers M F,Nicks L J,Robinson B H.1998.Phytomining.T Plant Sci,3(9):359-362.
Ebbs S D,Kochian L V.1997.Toxicity of zinc and copper to Brassica species:implications for phytoremediation.J Environ Qual,26:776-781
Escarre J,Lefebvre C,Gruber W,Leblanc M,Lepart J,Riviere Y,Delay B.2000.Zinc and cadmium hyperaccumulation by Thlaspi caerulescens from metalliferous and nonmetalliferous sites in the Mediterranean area:implications for phytoremediation.New Phytol,145:429-437.
Godbold D L,Horst W J,Marschner H,Collins J C,Thurman D A.1983.Root growth and Zn uptake by two ecotypes of Deschampsia caespitosa as affected by high Zn concentrations.Plant Physiol,112:315-334.
Monni S,Uhlig C,Hansen E,Magel E.2001.Ecophysiological responses of Empetrum nigrumto heavy metal pollution.Environmental Pollution,112:121-129.
Reeves R D,Baker A J M.2000.Metal accumulating plants//Raskin I,Ensley B D.Phytoremediation of toxicmetals:using plant to clean up the environment.New York:John Wiley Press.
Wu J,Schat H,Sun R,Koornneef M,Wang X W,Aarts M G M.2007.Characterization of natural variation for zinc,iron and manganese accumulation and zinc exposure response in Brassica rapa L.Plant Soil,291:167-180.
猜你喜欢
趣味(作文与阅读)(2022年3期)2022-06-10
今日农业(2022年4期)2022-06-01
小读者(2021年6期)2021-11-23
趣味(作文与阅读)(2021年6期)2021-11-06
陶瓷学报(2021年2期)2021-07-21
沈阳工业大学学报(2020年6期)2020-12-29
有色金属加工(2020年5期)2020-10-23
快乐语文(2020年26期)2020-10-16
读者(2015年20期)2015-05-14
