
基于涡度相关法的中国草地生态系统碳通量研究进展
2010-03-31 06:12王云龙胡正华薛红喜
草业科学 2010年12期
李 琪,王云龙,胡正华,薛红喜,李 洁
(1.南京信息工程大学,江苏南京 210044;2.中国气象局气象探测中心,北京 100081)
陆地生态系统的碳氮循环是驱动生态系统变化的关键过程[1],由于下垫面的复杂性和人类活动的干扰,陆地生态系统碳循环存在着最大的不确定性,已成为全球变化研究的焦点。草地作为地球上分布最广的植被类型之一,对全球气候变化具有一定的影响和响应[2-4]。草地生态系统作为陆地生态系统的重要组成部分,在陆地生态系统碳循环过程中起着重要作用[5-6]。对草地生态系统碳循环过程及控制机理的研究是全球碳循环研究的热点[7]。
草地生态系统不同碳库碳通量的观测方法主要有4类:化学法、箱式法、微气象法和土壤浓度廓线法,目前常用的是箱式法和微气象法。箱式法对表面状态的要求不高,但测量结果外推的准确度受到怀疑,而且不能用于大型植物构成的植被;微气象法避免了箱式法的局限性,但它对地表的均匀性和大气状态的要求很高[8]。涡度相关技术是一种非破坏性的微气象观测技术,近年来涡度相关技术的成熟使得长期、连续的观测成为可能,其在生态系统尺度上研究碳收支的时间格局、变率及其对环境因素的响应机理具有一定的优势。目前,涡度相关技术已经广泛应用于陆地生态系统碳循环的测定中,并逐渐成为生态系统水平直接测量CO2通量的标准方法[9-11]。本研究就近年来利用涡度相关技术对中国草地生态系统碳通量进行研究的结果作以概述,分析草地生态系统碳通量的不同时间尺度的变化特征以及环境控制机理,以期能够为中国的草地生态系统碳管理提供理论支撑,为中国实现减排目标、参与国际合作研究、履行相关国际公约提供技术支撑。
1 碳通量在不同时间尺度上的变化
草地生态系统碳通量在不同时间尺度上的变化规律是正确认识草地生态系统碳源/汇性质的基础,中国学者从日、季节、年际等不同时间尺度对典型草地类型进行了研究。
1.1 日变化在非生长季,草地净生态系统CO2交换(net ecosystem CO2exchange,NEE)的日变化不明显,波动多且昼夜变化幅度小,表现为CO2的释放。在生长季,随着气温的升高和降水量的增加,草地生态系统日间CO2吸收量和夜间CO2排放量都开始增大,出现了明显的CO2吸收和释放峰值。在通常情况下,草地生态系统NEE在凌晨由正值(释放)转换为负值(吸收),碳吸收能力随着光合有效辐射的增加而增加,大概在正午前后吸收量达到最大,随后碳吸收逐渐减弱,傍晚NEE由负值转换为正值[12-16]。草地生态系统NEE的这种变化进程是典型的单峰型,而且会出现上午碳吸收强、下午吸收弱的不对称现象[17]。
观测显示,草地生态系统NEE的日进程除了单峰型外,还有双峰型[18-20]。如薛红喜等[20]发现在生长季的大部分时间里,克氏针茅(Stipa krylovii)草地生态系统的NEE都表现出单峰型特征,双峰型主要发生在生长季后期,即7:30以后克氏针茅草地开始吸收CO2,9:30左右达到第1个吸收高峰后,很快转变为释放CO2,13:00以后又快速转变为吸收CO2,并在13:30左右达到第2个吸收高峰,之后碳通量开始下降,大约在17:00左右生态系统开始向大气排放CO2。从中国典型草地生态系统已观测到的CO2吸收速率的极值可以看出(表1),中国草地生态系统NEE的最大吸收速率因草地类型的不同而有很大差异。
1.2 季节和年际变化草地类型及所处环境不同,导致植被的光合作用、土壤呼吸以及各种环境因子季节变化的差异,进而会影响到草地生态系统NEE的季节动态特征。通常而言,5-9月为中国草地生态系统的碳吸收期,以7、8月的碳吸收最强,同时碳释放最大值也往往出现在这一时期。这说明对草地生态系统而言,7、8月的水热条件适宜,植物的光合作用和呼吸作用都比较旺盛,土壤呼吸也达到比较高的水平,因而NEE的日最低和最高值都出现在这一时期[1,5,16,20-22]。中国典型草地生态系统CO2日吸收极值也有明显的差异(表1)。
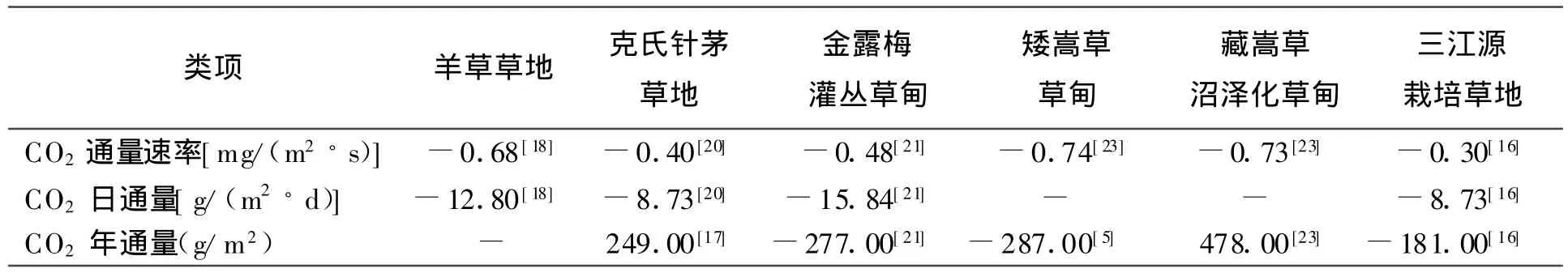
表1 中国典型草地生态系统CO2通量极值
赵亮等[23]的研究表明,青藏高原的3种草地生态系统的碳通量变化有明显差异,如5月嵩草草甸NEE有正、负变化过程,而灌丛草甸和沼泽草甸不存在正负变化过程。对同一生态系统而言,也存在明显的季节变化特征,如郝彦宾等[19]发现内蒙古羊草草地2003年5月中旬开始吸收CO2,而2004年直到6月初才开始吸收CO2,比2003年推迟将近20 d,吸收高峰也有所推迟。同样,Zhao等[24]研究显示嵩草草甸生态系统5月即进入生长期,但石培礼等[25]、Shi等[26]研究显示,嵩草草甸生态系统只有7-9月为碳吸收,其余月份均为碳排放。
除了生长期的时间不同外,同一生态系统的碳通量格局也会因环境因素的不同而产生差异。如杨娟等[15]发现克氏针茅草地生态系统在2004年呈现出2个碳吸收高峰,而薛红喜等[20]则发现2008年出现3个碳吸收高峰,并认为降雨量的不同是造成这种差异的原因。
草地生态系统NEE的年际变化情况可以反映某一草地类型的碳源/汇特性。中国草地生态系统CO2全年交换量因植被类型的不同而不同,因此相对应的生态系统的碳源/汇功能也不同。从表1可以看出,青藏高原的金露梅灌丛草甸和矮嵩草草甸、三江源栽培草地都表现出了碳汇的特征[21-22,27],而青藏高原藏嵩草沼泽化草甸、克氏针茅草地则表现出了碳源的特征[17,23-24]。对同一植被类型而言,环境条件的不同也会影响草地生态系统的碳源/汇特性。如当雄草原化嵩草草甸2004年表现为弱小的碳汇,而2005年则表现为较小的碳源,草地生态系统碳源/汇特征的转换主要与年降水量和降水分配有较大关系[28]。
2 影响碳通量的环境因子
2.1 温度2.1.1空气温度 空气温度是影响草地生态系统碳通量的重要因子,但不同的草地类型对温度的响应是不一样的。如青藏高原矮嵩草草甸生态系统每日的NEE与大气温度有显著的相关性[29]。而青藏高原高寒灌丛草甸在暖季夜间温度与NEE有显著正相关关系,白昼NEE与温度无显著关系;在冷季不论夜间还是白昼,NEE变化均与温度密切相关[30-31]。
不同的温度条件尤其是低温和高温都会对草地生态系统的固碳能力产生影响。如青藏高原高山嵩草草甸生态系统的叶面积指数较高,有较强的固碳潜力,但青藏高原较低的温度限制了其对碳的吸收[27]。同样,高温也会降低草地生态系统的生产力[32-34]。
另外在特殊的环境条件下,草地生态系统对温度的响应也可能是不敏感的,如在干旱胁迫下,羊草生态系统的NEE和温度在小时、日、月尺度上都没有显著的相关关系[35]。
昼夜温差也会影响草地生态系统的碳通量。研究显示,金露梅灌丛和矮嵩草草甸生态系统NEE随日温差的增加而增大,但二者之间的相关性不甚显著[36]。但Shi等[26]和Zhao等[37]认为,昼夜温差对碳固定影响不显著,甚至具有负效应。赵亮等[16]研究也认为生长季昼夜温差大并不利于三江源栽培草地生态系统的碳固定。
2.1.2土壤温度 土壤温度也是影响草地生态系统的重要因素,研究显示青藏高原高寒灌丛草甸生态系统非生长季NEE与5 cm土壤温度变化呈显著正相关,土壤温度是影响非生长季高寒灌丛草甸NEE变化的主导气候因子[38-39]。徐玲玲等[13]的研究进一步表明青藏高原高寒草甸夜间的NEE与5 cm地温有很好的指数函数关系。
土壤温度在作用于草地生态系统的碳通量时,还会受到土壤水分的影响。如研究表明,土壤水分充足时温度变化能够很好地解释克氏针茅草地生态系统呼吸的变异,但在干旱胁迫的条件下,生态系统呼吸与土壤温度的拟合关系较好,而与温度的拟合效果较差[17]。在矮嵩草草甸区,当土壤水分小于0.40 m3/m3或大于0.50 m3/m3时,CO2通量随着土壤温度升高而增大;土壤水分在0.40~0.50 m3/m3时,CO2通量不随土壤温度的增大而改变,基本保持稳定。在金露梅灌丛草甸区,不同土壤水分条件下的CO2通量与土壤温度关系与矮嵩草草甸区截然相反,当土壤水分小于0.25 m3/m3或大于0.30 m3/m3时,CO2通量随土壤温度升高而降低;土壤水分在0.25~0.30 m3/m3条件下,CO2通量随土壤温度的升高而增加[36]。
2.2 水分2.2.1降雨 降雨是影响草地生态系统碳通量的重要因素,一方面降雨量和降雨格局(时间)会直接影响草地植被的生长,另一方面降雨还会通过影响土壤水分间接影响草地植被的生长[40-41]。研究显示,生长季的降水量及其季节分配模式的不同对草地生态系统的生物物候有明显的影响,从而影响羊草草地生态系统碳源/汇的转换时间。如在2003年 6月初羊草生态系统开始净吸收CO2、7月初出现最大CO2吸收量,随后发生的干旱和高温胁迫使其在 8月就提早进入休眠期;2004年春季的严重干旱导致生态系统在7月初才开始净吸收CO2,在降雨丰沛气温适宜的8月出现最大CO2吸收量并使得CO2吸收持续到9月中旬[33-34]。
Zhao等[21]的研究也表明,降雨格局可以影响灌丛的生长季长短以及生态系统的呼吸,因此降雨时间对高山灌丛草甸生态系统碳通量的影响大于降雨总量。石培礼等[25]、张法伟等[42]的研究表明,降雨过程可以提高青藏高原高寒草甸生态系统的碳通量值,从而降低了生态系统的碳汇功能。降雨削弱了光合有效辐射对白天生态系统NEE的影响,而增加了地温对夜间呼吸的控制;降雨强度对白天NEE几乎没有影响,但能降低夜间呼吸。Shi等[26]也通过研究认为降雨格局是影响青藏高原草原化嵩草草甸生态系统碳收支的重要因素。
2.2.2土壤水分 土壤湿度对土壤呼吸的影响也很重要,尤其在干旱或半干旱地区当土壤水分成为胁迫因子时可能取代温度而成为土壤呼吸的主要控制因子,从而影响整个生态系统的碳通量。在干旱的年份中,土壤水分是影响羊草草地生态系统碳源/汇转换的重要因素[33]。黄祥忠等[43]的研究显示,锡林河流域内的羊草草地生态系统生长季内的白天NEE与土壤含水量的变化呈现显著的负相关关系。王云龙[17]研究认为降雨的季节分布通过土壤含水量的季节变异间接影响克氏针茅草地NEE的季节变化。
2.3 光合有效辐射光照是植物生长的重要因子,当生长季温度适宜时,生态系统的初级生产强烈依赖于光照条件[10]。Zhao等[24]的研究表明,青藏高原地区的3种草地生态系统在生长季NEE日变化主要由光合有效辐射控制,其他学者通过研究也得到了相似的结论[12-13,25-26,30,39]。Zhao等[37]的研究则进一步表明在低光照水平下,当光合有效辐射增大时,高寒灌丛草甸生态系统吸收碳的能力迅速增加。另外羊草草地、高寒灌丛草甸、栽培草地生态系统的碳通量有光饱和现象,即光合有效辐射达到一定值时,随着光照强度的增加,草地生态系统的碳吸收能力反而下降[16,19,32]。但是郝彦宾[35]的研究显示,在水分胁迫下,羊草生态系统NEE与光合有效辐射之间没有显著的相关关系。
2.4 积雪研究表明,在冬季有积雪时,CO2通量与5 cm土温相关不显著,积雪显著地影响着青藏高原北部地区CO2通量,但这种影响因生态系统类型的不同而异,高寒草甸CO2交换量随着积雪时间的延长呈线性降低,而高寒灌丛和高寒沼泽生态系统则不随积雪时间的延长而降低,沼泽和草甸在有积雪时CO2通量显著高于无雪时,而灌丛在这2个条件下CO2通量值差异不显著[44]。因此在冬季,积雪是影响青藏高原草地生态系统碳通量的重要因子。
2.5 植被对碳通量的影响对羊草草地生态系统2个生长季的观测发现,当年生物量积累和叶面积指数与CO2通量之间的关系存在年际间差异。如2个生长季最大地上生物现存量都出现在8月,但2003年碳吸收高峰期在6月中旬-7月中旬,CO2通量和地上生物量之间没有显著的关系;而2004年CO2通量出现的高峰期和最大地上生物量与叶面积指数相一致,具有较好的相关性。发生这种现象的原因很可能是与CO2通量有关的非生物因素部分,和生物量累积并不相关[18-19]。
3 结语与展望
综上所述,中国学者利用涡度相关技术对温带草原、高寒草地、栽培草地等典型生态系统都开展了定位观测,在草地生态系统碳通量及其驱动机制研究方面已经有了较大进展。但受观测时间较短、技术和方法还不成熟等诸多因素的限制,中国草地生态系统碳通量研究还需要在以下几方面做深入的工作:
1)目前中国草地碳通量的观测主要集中在典型草地和高寒草甸上,但已有的研究结果表明中国草地生态系统NEE测定值之间存在较大的空间变异性,这说明现有的站点还不具备精确评估我国草地碳储量的能力。为了准确评估中国草地生态系统碳源/汇功能以及空间分布格局,必须增加草地碳通量站点的密度,并在原有类型的基础上加大对半湿润的草甸草地和干旱区的荒漠化草地研究力度[45]。另外中国草地生态系统碳通量的连续观测时间相对较短,利用短期观测数据进行生态系统碳源/汇的评价会造成很大的不确定性,还需要长时间尺度的观测数据来分析中国草地生态系统的源/汇功能。
2)尽管涡度相关技术是目前碳通量研究的标准方法,但仪器本身的物理限制、二维和三维的气流运动、数据处理的方法和夜间通量的低估等不确定性因素仍然是今后通量技术观测和研究中需要优先考虑的问题[46-47]。完善的数据质量控制与分析以及误差评价是深入研究草地生态系统碳通量及其驱动机制的前提。另外,应结合其他方法进行草地生态系统的碳通量研究,为准确评估中国草地生态系统的碳积蓄和碳循环提供全方位的技术支撑。
3)未来气候变化背景下草地的适应性研究是今后中国草地生态系统管理以及畜牧业可持续发展的前提,而草地生态系统碳通量的驱动机制研究则是正确认识草地生态系统对气候变化的适应性的基础,同时也是发展具有自主知识产权的草地生态系统碳循环模型的前提。中国学者的研究主要集中在自然因子以及单因子驱动机制上,对草地生态系统碳循环过程的描述和参数化还不够[8],在草地生态系统碳循环模型研究方面几乎还是空白。今后需要加强对人为因子以及多因子之间的协同影响的研究[48],深化对草地生态系统碳循环的认识与理解,为提高以CO2减排与增汇为目的的草地生态系统管理水平以及畜牧业应对气候变化的适应能力提供科学依据。
4)涡度相关技术只是碳循环研究的一个工具,陆地生态系统碳循环机理的复杂性要求在研究中需要运用地理学、生态学、植物生理学、气象学等多学科的理论知识,将传统的生态学研究方法与地面通量观测、区域遥感观测和碳循环模型相结合,形成跨区域、多尺度、多途径地综合性观测研究体系,并运用数据模型融合和尺度扩展的方法,更准确地量化中国草地生态系统的碳收支状况[17,49],从而深入探讨中国草地生态系统碳收支及其分布格局以及草地功能对未来全球变化的响应与适应性机制。
[1]赵亮,古松,徐世晓,等.青藏高原高寒草甸生态系统碳通量特征及其控制因子[J].西北植物学报,2007,27(5):1054-1060.
[2]Scurlock J M O,Hall D O.The global carbon sink:a grassland perspective[J].Global Change Biology,1998,4:229-233.
[3]戴雅婷,那日苏,吴洪新,等.我国北方温带草原碳循环研究进展[J].草业科学,2009,26(9):43-48.
[4]周萍,刘国彬,薛萐.草地生态系统土壤呼吸及其影响因素研究进展[J].草业学报,2009,18(2):184-193.
[5]Zhang W L,Chen S P,Chen J,et al.Biophysical regulations of carbon fluxes of a steppe and a cultivated cropland in semiarid Inner Mongolia[J].Agricultural and Forest Meteorology,2007,146:216-229.
[6]钟华平,樊江文,于贵瑞,等.草地生态系统碳蓄积的研究进展[J].草业科学,2005,22(1):4-8.
[7]Ojima D S,Dirk B O M,Gleovn E P,et al.Assessment of C budget for grasslands and drylands of the world[J].Water Air and Soil Pollution,1993,70:95-109.
[8]钟华平,樊江文,于贵瑞,等.草地生态系统碳循环研究进展[J].草地学报,2005,13:67-73.
[9]Ciais P,Reichstein M,Niovy N,et al.Europe-wide reduction in primary productivity caused by the heat and drought in 2003[J].Nature,2005,437:529-533.
[10]Falge E,Tenhunen J,Baldocchi D,et al.Phase and amplitude of ecosystem carbon release and uptake potentials as derived from FLUXNET measurements[J].Agricultural and Forest Meteorology,2002,113:75-95.
[11]Gilmanov T G,Soussana J F,Aires L,et al.Partitioning European grassland net ecosystem CO2exchange into gross primary productivity and ecosystem respiration using light response function analysis[J].Agriculture,Ecosystems and Environment,2007,121:93-120.
[12]Xu L L,Zhang X Z,Shi P L,etal.Establishment of apparent quantum yield and maximum ecosystem assimilation on Tibetan Plateau alpine meadow ecosystem[J].Science in China Series D:Earth Sciences,2005,48(Supp.Ⅰ):141-147.
[13]徐玲玲,张宪洲,石培礼,等.青藏高原高寒草甸生态系统净二氧化碳交换量特征[J].生态学报,2005,25(8):1948-1952.
[14]徐世晓,赵新全,李英年,等.青藏高原高寒灌丛CO2通量日和月变化特征[J].科学通报,2005,50(5):481-484.
[15]杨娟,周广胜,王云龙,等.内蒙古克氏针茅草地生态系统-大气通量交换特征[J].应用生态学报,2008,19(3):533-538.
[16]赵亮,古松,周华坤,等.青海省三江源区人工草地生态系统CO2通量[J].植物生态学报,2008,32(3):544-554.
[17]王云龙.克氏针茅草地的碳通量与碳收支[D].北京:中国科学院研究生院,2008.
[18]Hao Y B,Wang Y F,Sun X M,et al.Seasonal variation in carbon exchange and its ecological analysis overLeymus chinensissteppe in Inner Mongolia[J].Science in China Series D:Earth Sciences,2006,49(Supp.Ⅱ):186-195.
[19]郝彦斌,王艳芬,孙晓敏,等.内蒙古羊草草原碳交换季节变异及其生态学解析[J].中国科学D辑:地球科学,2006,36(增刊Ⅰ):174-182.
[20]薛红喜,李琪,王云龙,等.克氏针茅草地生态系统生长季碳通量变化特征[J].农业科学环境学报,2009,28(8):1742-1747.
[21]Zhao L,Li Y N,Xu S X,et al.Diurnal,seasonal and annual variation in net ecosystem CO2exchange of an alpine shrubland on Qinghai-Tibetan plateau[J].Global ChangeBiology,2006,12:1940-1953.
[22]李英年,孙晓敏,赵新全,等.青藏高原金露梅灌丛草甸净生态系统CO2交换量的季节变异及其环境控制机制[J].中国科学D辑:地球科学,2006,36(增刊Ⅰ):163-173.
[23]赵亮,李英年,赵新全,等.青藏高原 3种植被类型净生态系统 CO2交换量的比较[J].科学通报,2005,50(9):926-932.
[24]Zhao L,Li Y N,Zhao X Q,et al.Comparative study of the net exchange of CO2in 3 type of vegetation ecosystems on the Qinghai-Tibetan plateau[J].Chinese Science Bulletin,2005,50:1767-1774.
[25]石培礼,孙晓敏,徐玲玲,等.西藏高原草原化嵩草草甸生态系统CO 净交换及其影响因子[J].中国科学D辑:地球科学,2006,36(增刊Ⅰ):194-203.
[26]Shi P L,Sun X M,Xu L L,et al.Net ecosystem CO2exchange and controlling factors in a steppe——Kobresiameadow on the Tibetan Plateau[J].Science in China Series D:Earth Sciences,2006,49(Supp.Ⅱ):207-218.
[27]Kato T,Tang Y H,Gu S,et al.Carbon dioxide exchange between the atmosphere and an alpine meadow ecosystem on the Qinghai-Tibetan Plateau,China[J].Agricultural and Forest M eteorology,2004,124:121-134.
[28]于贵瑞,孙晓敏.中国陆地生态系统碳通量观测技术及时空变化特征[M].北京:科学出版社,2008.
[29]Zhang F W,Li H Q,Li Y N,et al.“ Turning point air temperature”for alpine meadow ecosystem CO2exchange on the Qinghai-Tibetan Plateau[J].Pratacultural Science,2007,24(9):20-28.
[30]徐世晓,赵亮,李英年,等.温度对青藏高原高寒灌丛CO2通量日变化的影响[J].冰川冻土,2007,29(5):717-721.
[31]徐世晓,赵亮,李英年,等.青藏高原高寒灌丛暖季CO2地-气交换特征[J].中国环境科学,2007,27(4):433-436.
[32]Fu Y L,Yu G R,Sun X M,etal.Depression of net ecosystem CO2exchange in semi-aridLeymus chinensissteppe and alpine shrub[J].Agricultural and Forest Meteorology,2006,137:234-244.
[33]Fu Y L,Yu G R,Wang Y F,et al.Effect of water stress on ecosystem photosynthesis and respiration of aLeymus chinensissteppe in Inner Mongolia[J].Science in China Series D:Earth Sciences,2006,36(Supp.Ⅱ):196-206.
[34]伏玉玲,于贵瑞,王艳芬,等.水分胁迫对内蒙古羊草草原生态系统光合和呼吸作用的影响[J].中国科学D辑:地球科学,2006,36(增刊Ⅰ):183-193.
[35]郝彦宾.内蒙古羊草草地碳通量观测及其驱动机制分析[D].北京:中国科学院研究生院,2006.
[36]赵亮,徐世晓,李英年,等.青藏高原矮嵩草草甸和金露梅灌丛草甸CO2通量变化与环境因子的关系[J].西北植物学报,2006,26(1):133-142.
[37]Zhao L,Li Y N,Gu S,et al.Carbon dioxide exchange between the atmosphere and an alpineshrubland meadow during the growing season on the Qinghai-Tibetan Plateau[J].Journal of Integrative Plant Biology,2005,47(3):271-282.
[38]徐世晓,赵亮,赵新全,等.青藏高原高寒灌丛非生长季节CO2通量特征[J].西北植物学报,2006,26(12):2528-2532.
[39]徐世晓,赵新全,李英年,等.青藏高原高寒灌丛生长季和非生长季CO2通量分析[J].中国科学D辑:地球科学,2004,34(增刊Ⅱ):118-124.
[40]Xu L L,Zhang X Z,Shi P L,et al.Characteristics of net ecosystem carbon dioxide exchange(NEE)from August to October of Alpine meadow on the Tibetan Plateau,China[J].Frontiers of Biology in China,2006,1:418-422.
[41]徐世晓,赵亮,李英年,等.降水对青藏高原高寒灌丛冷季CO2通量的影响[J].水土保持学报,2007,21(3):193-200.
[42]张法伟,李英年,赵新全,等.一次降水过程对青藏高原高寒草甸CO2通量和热量输送的影响[J].生态学杂志,2008,27(10):1685-1691.
[43]黄祥忠,郝彦宾,王艳芬,等.极端干旱条件下锡林河流域羊草草地净生态系统碳交换特征[J].植物生态学报,2006,30(6):894-900.
[44]赵亮,徐世晓,伏玉玲,等.积雪对藏北高寒草甸CO2和水汽通量的影响[J].草地学报,2005,13(3):242-247.
[45]Yu G R,Fu Y L,Sun X M,et al.Recent progress and future directions of China FLU[J].Science in China Series D:Earth Sciences,2006,49:1-23.
[46]温学发,于贵瑞,孙晓敏.基于涡度相关技术估算植被/大气间净CO2交换量中的不确定性[J].地球科学进展,2004,19(4):658-663.
[47]于贵瑞,张雷明,孙晓敏,等.亚洲区域陆地生态系统碳通量观测研究进展[J].中国科学D辑:地球科学,2004,34(增刊Ⅱ):15-29.
[48]耿元波,董云社,齐玉春.草地生态系统碳循环研究评述[J].地理科学进展,2004,23(3):74-81.
[49]于贵瑞,伏玉玲,孙晓敏,等.中国陆地生态系统通量观测研究网络(ChinaFLUX)的研究进展及其发展思路[J].中国科学 D辑:地球科学,2006,36(增刊Ⅰ):1-21.
猜你喜欢
成都信息工程大学学报(2022年4期)2022-11-18
水土保持学报(2022年5期)2022-10-10
青海草业(2022年2期)2022-07-23
西北农林科技大学学报(自然科学版)(2022年3期)2022-04-01
四川冶金(2021年3期)2021-01-25
草业科学(2019年5期)2019-06-04
活力(2019年21期)2019-04-01
天津师范大学学报(自然科学版)(2016年4期)2016-12-14
河北林业科技(2016年5期)2016-11-08
中国学术期刊文摘(2016年1期)2016-02-13
