
丛枝菌根(AM)对水稻生长促进及化肥减量研究
2010-03-14 06:38张淑娟李世阳吴洁婷
哈尔滨工业大学学报 2010年6期
张淑娟,王 立,马 放,李世阳,张 雪,吴洁婷
(哈尔滨工业大学城市水资源与水环境国家重点实验室,哈尔滨150090,wli@hit.edu.cn)
农业面源污染是我国地表水体产生富营养化的主要原因,因其影响范围广、难于控制一直是环境治理领域的难题[1-3].近年来随着粮食需求的不断增加,为获得农作物高产,往往在农田生态系统中施加大量的化肥,这些化学产物的当季利用效果较差,氮、磷、钾肥当季利用率分别为30%~35%,10%~25%,35%~50%[4].大部分没有被利用的营养物质流失于土壤、水体和空气中,并在灌溉与降水的淋溶与迁移作用下通过地表径流进入受纳水体,形成严重的农业面源污染[3].
丛枝菌根(Arbuscular mycorrhiza,AM)作为植物根系和丛枝菌根真菌所建立的共生系统,能够有效增强作物的物质生产和抗病能力,因此广泛应用于园艺[5]、林木[6-7]、药材[8-9]等方面.近年来,AM在“绿色肥料”和“绿色农药”开发方面的巨大潜力日益引起人们的重视[10-13].一方面,AM可通过增加根际土壤中微生物的数量[14-15]、促进作物对中微量营养元素的吸收[16]、改善植物的碳、氮、磷素营养状况[17-20]等途径促进植物的生长,减少化肥的施用量;另一方面AM通过提高作物的抗病虫害能力[21-22],减轻化学杀虫剂的环境施入负荷.AM的优良特性能够在保证作物丰产的同时,减少化肥和杀虫剂等面源污染物的施用量,减轻对农产品、地下水以及地表水资源的污染程度,是新型生态农业的发展方向,并有望成为农业面源污染源头减量的有效生物技术.
水稻作为世界上最重要的粮食作物之一,其生产带来的农业面源污染也极其严重.此外,作为典型的湿生作物,水稻种植环境的特殊性更易导致最为严重的面源污染物扩散问题[23-25].基于此,本试验选用水稻为试验材料,研究丛枝菌根对其生长繁殖的影响,藉以揭示丛枝菌根在农业面源污染化肥减量过程中可能发挥的重要作用.
1 材料与方法
1.1 供试材料
试验在城市水资源与水环境国家重点实验室的试验基地进行.
气候条件:哈尔滨市位于北纬45°40′的寒温带地区,本试验在作物生长季5月到9月进行,此季哈尔滨历年平均气温为20.8℃,历年变化为19.4~22.4℃;平均降水为379 mm;历年变化为197.5~633.7 mm.
供试菌剂:供试AMF为广适菌种——摩西球囊霉Glomus mosseae(G.mosseae).
供试基质:以田间土壤、沙、蛭石(2∶5∶3)混合配制的营养土为栽培基质.田间土壤pH值为7.7,全氮6.75 g/kg,全磷13.07 g/kg.
盆栽试验:盆钵高30 cm,盆口直径34 cm,盆底直径18 cm.移栽试验样本前用0.1%的高锰酸钾溶液浸泡30 min,5 h后加入相应基质8 kg.
供试植物:选用农作物水稻(Oryza sativa L.),品种为合粳1号,种子来于黑龙江省农科院.种子用水浸泡24 h后,30℃保湿催芽.待芽长至1 cm左右时,筛选芽长基本一致的水稻种子均匀的撒在长条花盆中,表面覆土约2 cm每天早晚浇水两次.待幼苗长至3叶期移栽到盆钵中进行试验.
1.2 试验设计
试验设置AMF空白基质、自然基质和AMF强化基质3种基质条件.空白基质在压力(0.15 MPa)、温度(121℃)条件下蒸气灭菌2 h.AMF强化基质条件中每盆接种有效菌剂140 g,在自然基质和空白基质中添加140 g菌剂的滤纸滤液,以保证3个实验条件除菌根真菌以外微生物群落的一致性.每个基质条件设置3个重复.所有基质灌水至盆底有水溢出,2 d后移栽水稻幼苗,每盆20穴,每穴3株.花盆底部的阀门控制水稻的灌水和退水,水分按当地高产栽培要求进行控制.
1.3 调查项目及测定方法
AMF侵染率:待幼苗生长7周后,随机选取各条件中1~2 cm长根尖进行染色[26],每个重复观察100个根段.
AMF侵染率(%)=(侵染根段数/观察根段数)×100%[26].
千粒重:200粒籽实称得重量×5,重复5次测得千粒重.
总生物量:地下部分洗净泥土并用滤纸吸干表面水分后和地上部分105℃杀青5 min后60~80℃烘干至恒重,称量得地上、地下生物量,两部分之和为总生物量.
菌根依赖性=AMF强化水稻总生物量/AMF空白水稻总生物量×100%.
根总表面积:美国Micromeritics公司生产的ASAP2020型比表面积与孔径分析仪,测得BET比表面积.根总表面积=根比表面积×地下生物量.
2 结果与分析
2.1 不同基质条件下AMF的定殖情况
为了确定各基质条件水稻根部AMF的侵染及产孢情况,在幼苗移栽的7周后对水稻根部进行了染色、镜检,结果见图1.计算各基质条件下AMF对水稻根部的侵染率,结果见图2.
由图1可知,不同基质条件下水稻根部在生长7周时已呈现明显的AMF侵染分异.由图1、2可知,AMF空白基质中水稻根部没有AMF孢子和菌丝的存在,侵染率为0%;自然基质中水稻根部被侵染的部分与AMF强化基质的相比孢子和菌丝数量较少,侵染率为25±1.024 9%;AMF强化基质中孢子数量较多,且菌丝粗壮、显微镜下观察较清晰,侵染率高达42±2.003 6%,比自然基质的高68%.自然基质中水稻根部的侵染源于土壤中土著AMF孢子的存在,因此侵染率较低,且菌丝细弱.AMF强化基质中接入的菌剂内含有大量孢子、菌丝、菌根根段或(和)孢子果等繁殖体,这些高活性的繁殖体同土壤中的土著AMF孢子对水稻根系形成强势侵染,因此侵染率较高.

图1 不同基质条件下AMF的定殖情况

图2 不同基质条件下水稻根部AMF侵染率
2.2 AMF接种对水稻营养生长的影响
2.2.1 AMF接种对水稻株高的影响
水稻株高是反映水稻营养生长状况最直观的指标.试验于水稻生长的第15天,60天,75天,95天,105天测量了各基质条件下水稻的株高,结果见图3.

图3 AMF接种对水稻株高的影响
如图3可知,AMF强化基质中水稻株高在15 d后与AMF空白基质和自然基质中培育的相比,分别提高24.5%、20.6%.表明在强化基质中,AMF与水稻根部在第15天时便形成了良好的共生关系.AM共生结构的形成,可有效促进作物对营养物质的吸收,促进作物生长.AMF空白基质和自然基质的株高差异不显著,表明AM共生结构只有达到一定数量时才能发挥其对水稻营养生长的促进作用.
2.2.2 AMF接种对水稻总生物量和根系总表面积的影响
于水稻生长季末考察了作物植株的总生物量及根系总表面积,结果见表1.

表1 AMF接种对水稻总生物量和根系总表面积的影响
由表1可知,AMF强化基质条件下水稻的总生物量较空白基质条件和自然基质条件分别提高29%和30%.由总生物量计算水稻菌根依赖性为123%,表明水稻最大生长量对摩西球囊霉有一定的依赖性,判定水稻为菌根依赖性作物.另外,3种基质条件下水稻具有不同的根系总表面积,自然基质与AMF空白基质差异不显著(P>0.05),说明自然条件下形成的少量菌根结构不能明显改变根系结构与吸收面积.而AMF强化基质条件下根系总表面积较空白基质条件和自然基质条件分别增大了33.6%、36.6%.大量菌丝的存在扩大了水稻根系的总表面积,增加了基质中营养的空间有效性,使原来在空间上对水稻无效的养分变成有效养分,改善水稻营养水平以保证作物旺盛生长.
2.3 AMF接种对水稻籽实产量的影响
在水稻腊熟期后对其进行取穗考产,调查各基质条件下水稻的产量,结果见表2.
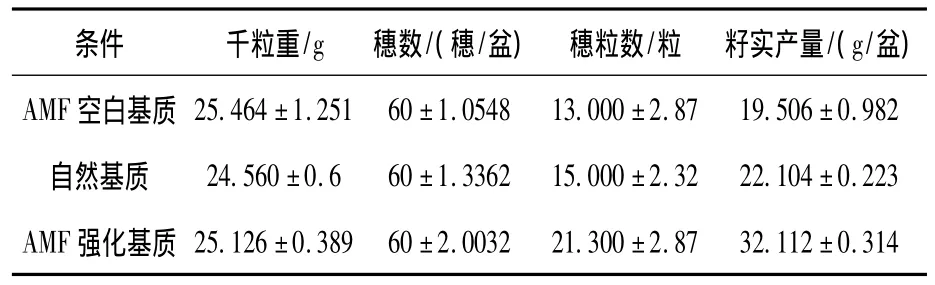
表2 AMF接种对水稻籽实产量的影响
由表2可知,3种基质条件下水稻的千粒重和穗数差异不显著(P>0.05),但是穗粒数差异达到了极显著水平(P<0.01).穗粒数的大小顺序为AMF空白基质<自然基质<AMF强化基质,自然基质的比空白基质的高15.4%,AMF强化基质分别比 AMF空白基质和自然基质高15.38%、42%.从籽实产量来看,各处理的大小顺序同穗粒重,且自然基质的比空白基质的高13.33%,AMF强化基质分别比AMF空白基质和自然基质高64.6%、45.3%.因此,AMF强化基质籽实产量较高的原因与千粒重无关,但与穗粒数关系密切(相关系数为0.999 3).表明籽实颗粒是一个较为稳定的生物学性状[27-28],环境可塑性小,AMF对作物产量的促进是通过提高结实率来实现的.
2.4 AMF接种在农业面源污染化肥减量中的作用
通过对AMF强化和化肥对植物生长促进状况的对比分析,可知AMF在农业面源污染化肥减量中的作用效果.采用自然基质条件和AMF强化基质条件生产100 kg水稻所需的化肥量之差来衡量AMF的强化作用.计算过程中涉及到的公式
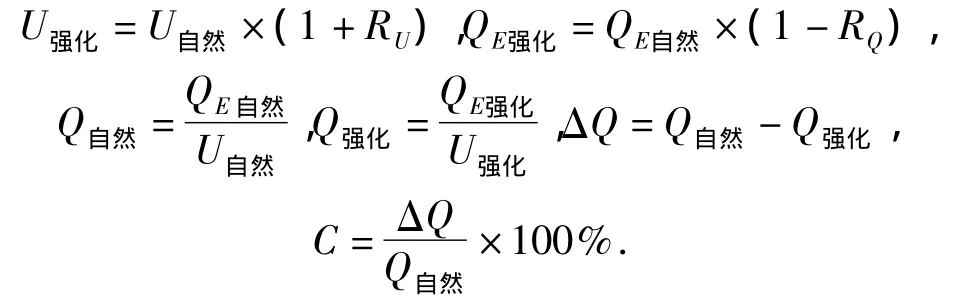
式中:U强化/U自然为强化/自然条件下化肥当季利用率(%);QE强化/QE自然为强化/自然条件下生产100 kg水稻元素需要量(kg);Q强化/Q自然为强化/自然条件下生产100 kg水稻化肥施用量(kg); RU/RQ为强化基质条件下水稻总生物量/产量较自然基质增加的百分比(%);Q为生产100 kg水稻时AMF的强化作用(kg);C为削减程度(%).
计算结果见表3.
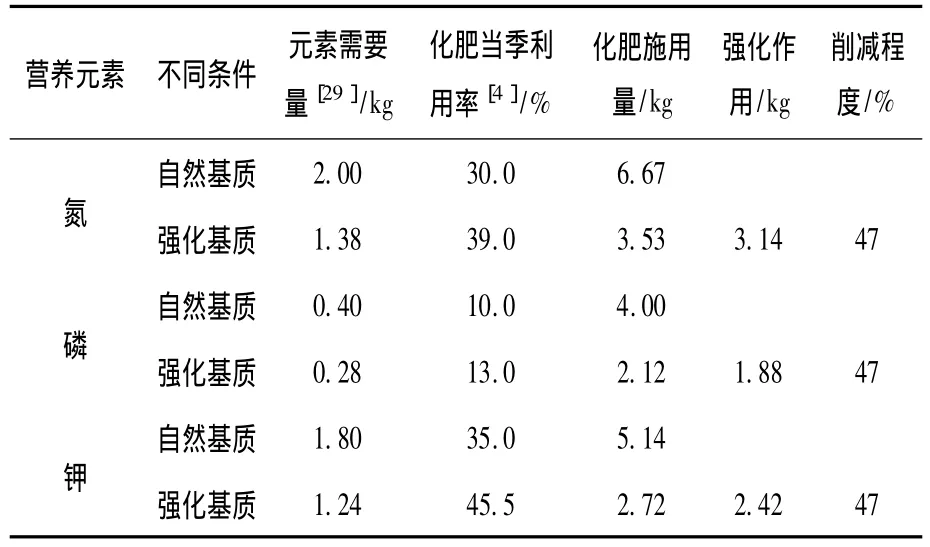
表3 AMF接种在农业面源污染化肥减量中的作用
由表3可知,每生产100 kg水稻,AMF强化条件相对于自然条件下氮肥、磷肥、钾肥施用量分别减少3.14,1.88,2.42 kg,削减程度高达47%.
3 结论
1)人工强化条件下水稻根部AMF的侵染率与自然基质条件相比提高了68%,这不仅突破了湿生环境AMF接种这一技术问题,还保证了将丛枝菌根用于水稻的科技种田及农业面源污染源头控制的基本前提.
2)与自然基质条件相比,AMF强化使水稻的株高、总生物量、根系总表面积、产量分别提高了20.6%、30%、36.6%、45.3%.说明水稻为菌根依赖性植物,计算其依赖程度达123%.这一结论为菌根技术用于水稻的清洁生产奠定了坚实基础.
3)人工强化接种AMF能使根系总表面积增加36.6%.根系巨大的总表面积是保证水稻生育后期较大的绿叶面积、较强的光合生产能力及获得高产的重要条件,因此在农业面源污染控制过程中对水稻进行AMF人工接种具有重要意义.
4)随着国际粮价全面上涨,特别是国际大米价格创出20年来的最高点,粮食问题再次成为一个热门话题.本研究发现AMF强化应用于水稻生产过程,其产量与自然基质相比提高了45.3%.将丛枝菌根用于水稻生产这一举措,可以在减少化肥的施用量的同时获得丰产,避免粮食产量下滑问题,保证我国粮食安全.
5)水稻作为种植面积最大的粮食作物,其生产带来的面源污染物流失问题极其严重.试验表明每生产100 kg水稻,AMF强化条件与自然条件相比化学肥料的削减程度可高达47%,因此可作为新型绿色生物载体应对当前日益严重的农业面源污染问题.丛枝菌根广泛应用于农业生产,是有效减少面源污染化肥施入并真正意义上实现绿色生态农业的必然发展趋势.
致谢:对黑龙江大学宋富强老师在菌种筛选过程中的帮助表示由衷的感谢.
[1]WANG Xiaoyan.Management of agricultural nonpoint source pollution in China:Current status and challenges[J].Water Sci Technol,2006,53(2):1-9.
[2]USEPA.National management measures to control nonpoint source pollution from agriculture[J].Office of Wetlands,Oceans and Watersheds 2003,42(3):55-60.
[3]ENTRYA J A,SOJKA R E,WATWOOD M,et al. Polyacrylamide preparations for protection of water quality threatened by agricultural run off contaminants[J]. Environmental Pollution,2002,120:191-200
[4]马继侠,袁旭音,徐海波.浅谈农田面源污染的生态控制技术[J].浙江水利科技,2009,162:1-3.
[5]李瑞卿,刘润进,李敏.园艺作物菌根及其在生态农业的应用[J].中国生态农业学报,2002,10(1):24-26.
[6]陈羽,弓明钦,仲崇禄.菌根菌剂在华南地区林业及农业上的应用[J].广东林业科技,2004,20(1):50-53.
[7]HELGASON T,MERRYWEATHERJ W,DENISON J,et al.Selectivity and functional diversity in arbuscular mycorrhizas of co-occurring fungi and plants from a temperate deciduous wood land[J].J Ecol,2002,90:371-384.
[8]任嘉红,刘瑞祥,李云玲.三七丛枝菌根(AM)的研究[J].微生物学通报,2007,34(2):224-227.
[9]杨光,郭兰萍,黄璐琦,等.药用植物的丛枝菌根接种方法研究.资源科学[J],2008,30(5):778-785.
[10]王立,贾文奇,马放,等.菌根技术在环境修复领域中的应用及展望[J].生态环境学报,2010,19(2):487 -493.
[11]王立,林楠,其其格,等.从枝菌根真菌(AMF)特性及其在环境生态修复中的应用[C]//第十二届海峡两岸环境保护学术研讨会论文集.高雄:海外华人环保学会,2008:5-12.
[12]王立,马放,任汲东,等.菌根真菌对环境的改造作用及其在环境修复领域中的应用[C]//2008年中国微生物生态学年会论文集.长沙:中国微生物生态专业委员会2008:79-84.
[13]张淑娟,王立,马放,等.菌根技术在农业面源污染防治中的应用[C]//植物内生菌开发与应用学术研讨会论文集.昆明:中国农业科技学会,2009:110-114.
[14]ÖPIK M,MOORA M,LIIRA J,et al.Composition of root colonizing arbuscular mycorrhizal fungal communities in different ecosystems around the globe[J].J Eco,2006,194:778-790.
[15]YAMATO M,IWASE K.Community analysis of arbuscular mycorrhizal fungi in a warm-temperate deciduous broad-leaved forest and introduction of the fungal community into the seedlings of indigenous woody plants[J].Mycoscience,2005,4(6):334-342.
[16]ARVALHO L M,CACADOR I,MARTINS-LOUCAO M A.Arbuscular mycorrhizal fungi enhance root cadmium and copper accumulation in the roots of the salt marsh plant Aster tripoliumL.[J].Plant Soil,2006,285:161 -169.
[17]TITUS J H,TITUS P J,NOWAK R S,et al.Arbuscular mycorrhizae of mojave desert plants[J].West Nam Naturalist,2002,62:327-334.
[18]ATUL-NAYYAR A,HAMEL C,HANSON K,et al. The arbuscular mycorrhizal symbiosis links N mineralization to plant demand[J].Mycorrhiza,2009,19:239 -246.
[19]ALONSO LM,KLEINER D,ORTEGA E.Spores of the mycorrhizal fungus Glomus mosseae host yeasts that solubilize phosphate and accumulate polyphosphates[J]. Mycorrhiza,2008,18:197-204.
[20]KLIRONOMOS J N.Variation in plant response to native and exotic arbuscular mycorrhizal fungi[J].Ecology,2003,84:2292-2301.
[21]李敏.AM真菌对西瓜抗枯萎病的效应及其机制[D].北京:中国农业大学,2005.
[22]朱红惠,龙良坤,羊宋贞,等.AM真菌对青枯菌和根际细菌群落结构的影响[J].菌物学报,2005,4(1): 137~142.
[23]张爱平.宁夏黄灌区稻田退水氮磷污染特征研究[D].银川:中国农业科学院,2009.
[24]黄沈发,沈根祥,唐浩.上海郊区稻田氮素流失研究[J].环境污染与防治,2005,27(9):651-654.
[25]许仙菊.上海郊区不同作物及轮作农田氮磷流失风险研究[D].上海:中国农业科学院,2007.
[26]刘润进,陈应龙.菌根学[M].北京:科学出版社,2007.
[27]赵利梅.水稻强化栽培农艺性状及其生态环境效应的研究[D].杭州:浙江大学,2009.
[28]张荣萍.不同灌水方式对水稻生育特性和产量及其水分利用率的影响[D].成都:四川农业大学,2006.
[29]鲁如坤,谢建章,蔡贵信,等.土壤——植物营养学原理和施肥[M].北京:化学工业出版社,1997.
猜你喜欢
中国农业科学(2022年17期)2022-09-19
商品与质量(2021年43期)2022-01-18
云南农业科技(2021年6期)2021-12-30
园林科技(2020年2期)2020-01-18
中成药(2018年3期)2018-05-07
中国资源综合利用(2016年9期)2016-01-22
中国果业信息(2014年2期)2014-01-23
植物营养与肥料学报(2011年5期)2011-11-06
植物营养与肥料学报(2011年6期)2011-10-24
植物营养与肥料学报(2010年3期)2010-11-16
