
遮荫对单性花同株植物麻疯树性比与构件大小的影响
2010-02-26 02:17何亚平费世民蔡小虎王乐辉陈秀明蒋建强雷彻虹
四川林业科技 2010年4期
何亚平,费世民**,蔡小虎,王乐辉,陈秀明,蒋建强,雷彻虹
(1.四川省林业科学研究院,四川 成都 610081;2.长江造林局攀枝花分局,四川攀枝花 617000;
3.攀枝花市林业局,四川 攀枝花 617000)
一般而言,在植物开花阶段,雌性适合度主要表现在雌性生殖器官的数量与质量上,而对于单性花同株同花序植物而言,雌性适合度则体现在雌花的数量、质量、雌雄花比例与两性资源配置等方面。很多研究都揭示了异质性光照环境对雌性适合度的影响,林地多年生植物雌性适合度也随着光照强度的增加而增加(Gregg,1975,1978);林地 1 a生植物在光照强度增加的环境中也存在明显的雌性偏向的两性配置(Cid-Benevento,1987)。一些研究都涉及到光照对性比波动或变异的影响问题,但对所有研究物种都没有得出一致的结论(Cid-Benevento,1987)。其本身可能忽略了个体大小自身对性比的塑造作用,而 Cid-Benevento(1987)在研究 1 a生林地植物 Acalypha rhomboidea(Euphorbiaceae)and Pilea pumila(Urticaceae)光照对雌雄配置的影响时,考虑了植株大小与环境湿度。植株大小或其他限制性环境因子、遗传因素存在差异而使得环境影响存在明显的不一致性,而使得对性比变异影响表现出复杂性与综合性。何亚平等(2009a)研究麻疯树构件层次上的性比变异时也发现,性比与构件大小指标呈现极显著正相关。针对木本植物而言,从构件层次上看性比变异,则明显看到性比实质上在构件之间呈现频度分布(何亚平等,2009a),即性比不是固定不变的,而是随着光照、湿度、土壤厚度和植株大小,乃至遗传因素而变异的(Lloyd,1972;Gregg,1975,1978;Cid-Benevento,1987;何亚平等,2008,2009a),性比对环境存在明显的可塑性反应(Conn&Blum U,1981;Costich,1995;Jean-Michel&Daniel,2003;尹春英,李春阳,2007)。
从综述来看,有些报道主要是关于雌雄异株植物性比波动问题、生境对雌雄性个体的选择问题,以及雌雄性个体的适应性表现问题(尹春英,李春阳,2007),而对单性花同株植物性比对环境的可塑性反应研究草本植物有所涉及。对于木本喜光植物而言,对其形态与生活史存在更为强烈的塑造作用,光照对其生殖器官及其相关特性的影响更为明显。一些研究都报道了性比对光照环境的适应性反应,在光照资源增加的条件下性比呈现偏雌性增加(Gregg,1975,1978;Cid-Benevento,1987;Cid-Benevento,1987),而对单性花同株木本植物的性比变异报道很少。基于影响性比变异因素的复杂性与综合性,探讨光照对喜光植物性比的影响时需要排除其他因素,野外调查性研究需要考虑植物个体大小、大气与土壤因素。麻疯树是一个适应于世界热带地区的一个全球广布泛热带植物(贾治邦,2008),具有耐干旱、耐热、喜光、耐瘠薄的特性,被印度人称之为“坚强的植物”(Openshaw,2000)。麻疯树光合作用研究(窦新永等,2008;黄红英等,2009;张诗莹,樊卫国,2005;夏亮等,2008)也表明,其本身光补偿点与光饱和点在木本植物中均较高,表现出明显的喜光特性。显然,遮荫条件下麻疯树可能会因为自身充分生长对光照强度的依赖性而呈现出可塑性反应,这种光限制会对单纯依赖营养供给来实现功能的生殖器官而言,表现得更为强烈,尤其是性比或两性配置。此外,性比对光照可塑性变异可能还存在一个潜在的因素,那就是雌雄性器官自身在生物学与生态学差异存在性别差异。
很多学者都探讨了单性花同株植物性比变异的植株大小依赖性问题(Solomon,1989;Bickel and Freeman,1993;Shaanker and Ganeshaiah,1984;Carmen,1987)。光照对营养生长的可塑性研究很多,其本身也通过限制光合作用而塑造生殖器官发育的营养载体,进而影响到生殖性状的发育。性比波动与植株大小紧密相连,这本身也是生殖器官与营养器官之间相关性的体现。众多研究都表明,光照限制时,植物叶片、个体、枝条、生物量配置、光合等都呈现明显的可塑性(肖春旺,刘玉成,1999;何维明,钟章成,2000;刘传照等,1991;廖咏梅等,2006;张教方等,1993;吴载璋,陈绍栓,2004;张效平,1990;陈绍云,周国宁,1992;樊后保,王义弘,1992;闫兴富,曹敏,2007;戴凌峰等,2008)。光照对营养器官的塑造必然会影响到生殖器官性状。
关于麻疯树性比问题,本文作者们做了一些研究。麻疯树为单性花同株同花序植物,花序为腋生聚伞花序,雌雄花在花序上存在明显的规律性分布(何亚平等,2008),性比的遗传作用极为强烈。但,初步研究表明,麻疯树雌雄花结构特性也会被数量发育所改变,其中雌花和雄花会因为某种因素而滞育,性比也具有明显的样地差异,环境可塑性明显;而且在构件水平上,性比存在明显的频度特征,与构件大小直接相关(何亚平等,2008,2009a)。尽管生殖器官自身也存在光合生产能力(Bazzaz et al.,1979),考虑到生殖器官自身依赖于营养器官提供物质才能建成器官,而构件是光合营养的单位,在研究研究遮荫对麻疯树性比的影响时,需要对与性比相关的构件营养器官部分特征进行相关调查与分析,兼顾传统性比(或生殖特性)研究中对雌雄性功能自身载体重视的不足。在本文研究中,重点考虑以下两个问题:(1)麻疯树遮荫条件下的构件大小特征;(2)麻疯树遮荫对雌花分化与性比特征的效应;(3)光照影响雌性适合度的过程问题。重点讨论光照资源限制通过影响构件大小来实现对性配置影响的过程,阐明性比环境可塑性的营养限制本质。
1 材料和方法
麻疯树群落物种组成较为复杂,但一般都呈现斑块状分布格局,或者与灌木呈现稀树灌木草丛状混交,不存在明显的遮光现象。研究表明,攀西地区麻疯树群落存在明显的乔木混交,主要混交树种有攀枝花、桉树、凤凰木、台湾相思等乔木与小乔木 14种,占植物种数的 31.82%(罗增斌等,2009)。而路旁、江边坡面防护林中存在乔木与麻疯树的混交林,并且成为金沙江攀枝花段支流路边防护林的主要特征。典型的遮光群落在倮果桥左侧存在麻疯树与高大乔木的混合群落,其乔木树种特征如表 1所示,构成群落郁闭度在 0.8左右。而林外有一块麻疯树林存在没有高大乔木,仅有混交灌木种类,如夹竹桃等,两块样地的麻疯树林分年龄差异不明显,土壤状况类似,样地概况如表 2所示。倮果桥林下存在部分薄土层个体,但大部分都为人工废物倾倒导致的松土堆积,土壤厚度大部分较大。可以把其土壤厚度平均为 60 cm以上,高大乔木生长良好。土壤厚度相似时可以认为其土壤养分和水分供给环境基本相同,二者具有相似的土壤环境,为分析光照环境影响和遮光处理提供了方便。
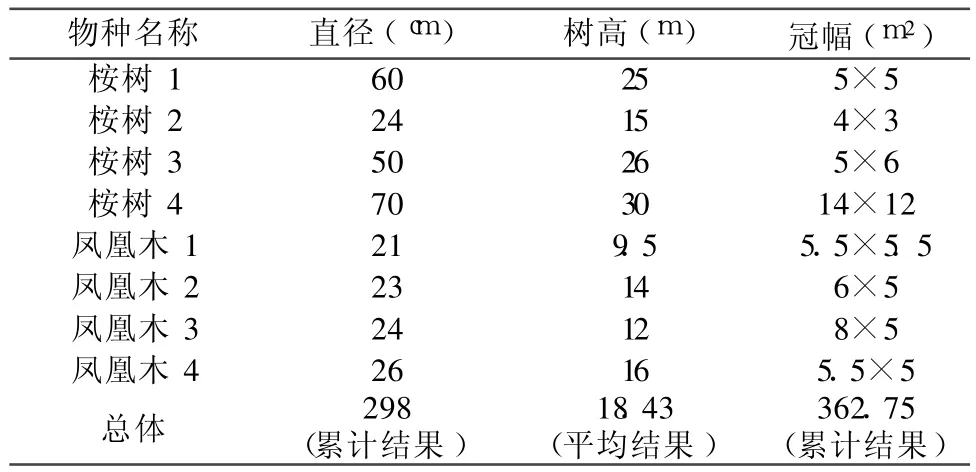
表 1 倮果桥乔木林树种组成与大小参数

表 2 倮果桥林下和林外麻疯树种群的群落状况与土壤环境
样地的自然地理概况与麻疯树物种特性如文献(何亚平等,2008,2009a,2009b)所示,为干热河谷向干暖河谷交错地带,其自身的植被存在明显的差异。主要表现为灌木林增加,而草丛减少,主要呈现灌木林。为中山峡谷地带,坡度较大,地势陡峭,坡面土壤厚度较薄,植被发育为灌木林已经达到顶级。
调查方法:大枝取样,测量头年生枝条的直径与枝长,统计雌花与雄花的数量。
数据分析:构件直径以 0.4 cm划分大小级,长度以 5 cm划分大小级;雌花数量以 3为单位划分大小级,性比以 0.02为单位划分大小级。把构件直径、枝长、雌花数量与性比进行频度分析,并作林内与林外比较,用 Spss for Windows 13.0进行 oneway ANOVA analysis。
2 结果与分析
2.1 遮光环境中的构件大小特征
遮光使得麻疯树光合作用受到光资源限制的影响,整体光合作用减弱,分支能力减弱,而头年生枝条发生显著性的变化,使得长度增加而直径减少。倮果桥林下与林外麻疯树构件头年生枝径、枝长如表 3所示,表明,林下构件直径为0.8112 cm±0.1973 cm,长度为 8.3255 cm±10.057 cm,而林外麻疯树构件直径为0.9125 cm±0.2809 cm,枝长为18.548 cm±19.3817 cm,并且直径与枝长在林下与林外存在极显著差异(如表 4所示)。从构件大小的频度分布来看,麻疯树林下构件直径频度主要集中在直径 0.4 cm~1.1 cm之间,占到 90%以上,而林外构件直径则以 0.4 cm~1.5 cm为主体,也占到90%以上(如图 2所示)。表明林下麻疯树直径偏小,而林外麻疯树构件直径明显偏大,与平均值相一致。构件枝长的频度分布在林外与林下差异明显(如图 2所示)。林下麻疯树构件枝长在较小长度级上明显较高,并随着长度级增加而逐渐减少。林外麻疯树构件枝长在较小长度级上频度较低,而在较大长度级上频度相对较高,并且频度随着长度级的增加而逐渐增加。从整体上看,林下构件长度偏小,而林外麻疯树构件长度明显偏大,与构件长度平均值结果一致。光照资源不足使得林下麻疯树相对高生长明显,而直径生长不明显,构件相对较细;而林外光照充足,麻疯树构件生长较为充足,直径较粗,构件绝对直径较大,相对较粗。不同光照资源使得构件大小存在明显差异,这种差异必然会对雌花分化与性比产生影响,这种影响也与雌雄花分化的生态学机制紧密相关。
2.2 遮光环境中的雌花数量与性比特征
遮光使得雌花分化能力呈现明显的差异,主要体现在雌花数量与性比上(如表 3所示)。倮果桥林下麻疯树雌花数量为2.7160/支 ±3.3052/支,而林外为5.0417/支 ±4.4106/支,二者差异极为明显(如表 4所示)。倮果桥林下性比为0.0278/支 ±0.0215/支,林外麻疯树性比为0.0378/支 ±0.0738/支,二者存在显著差异。从雌花数量与性比的频度上考虑,林下麻疯树雌花数量在低数量级上比例较高,如 0~2朵雌花的占到 60%以上,随着雌花数量级的增加而频度逐渐降低;林外麻疯树雌花数量在较低数量级较大,从图 2看频度都是逐渐降低。表明,林下雌花数量低数量级频度相对较大,而林外麻疯树构件雌花数量在同等级上逐渐增加,在高雌花数量级上频度相对较大(如图 2所示)。麻疯树性比以 0.02为等级,划分性比级,整体上攀枝花麻疯树性比级以低性比级为主体,而呈现倒 J型(何亚平等,2009)。林下遮光环境中麻疯树构件性比在 0~0.01上相对较高,而从 0.02级 ~0.03级以后逐渐降低,频度相对降低,并持续到较高的性比等级。而林外麻疯树构件性比在较低性比等级上较低,而在较高性比等级上性比频度逐渐增加。显然,遮光环境中麻疯树性比以低性比级频度相对较高,而强光环境中构件性比级以较高性比等级的构件频度相对较高,与林下与林外性比平均值差异一致(如图 2所示)。总体看来,遮光环境中麻疯树构件雌花分化能力相对较低,这与光照限制环境对构件生长的塑造有关,而性比与雌花分化现状与构件大小紧密相关。
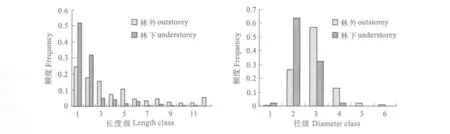
图 1 倮果桥林下与林外麻疯树构件直径与长度级的频度

图 2 倮果桥林下与林外麻疯树性比与雌花数量级的频度
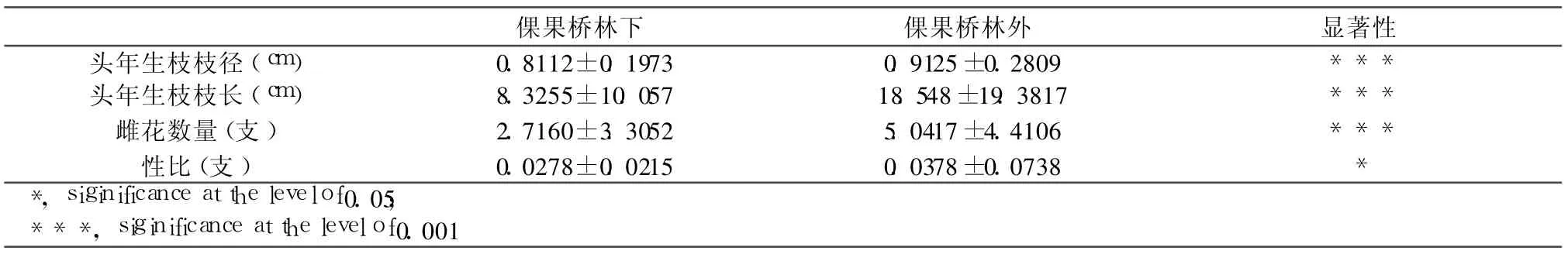
表 3 遮荫与不遮荫林地麻疯树群体构件头年生枝径、枝长、性比与雌花数量及其差异性

表 4 遮荫与不遮荫林地麻疯树群体构件头年生枝径、枝长、性比与雌花数量的ANOVA分析
3 讨论
光照是绿色植物生长发育不可或缺的资源,光照限制明显影响植物个体性状发育,使得植物产生与光照相关的形态与结构表现,这种影响在构件、器官、个体与群体上都有所体现。国内很多研究都探讨了光照限制条件下植物个体生长发育与开花结实的特性(肖春旺,刘玉成,1999;何维明,钟章成,2000;刘传照等,1991;廖咏梅等,2006;张教方等,1993;吴载璋,陈绍栓,2004;张效平,1990;陈绍云,周国宁,1992;樊后保,王义弘,1992;闫兴富,曹敏,2007),在形态与生理上从个体水平上阐明了光照对植物生长与生殖性状的影响表现。生殖是营养生长的副产物,与营养生长能力紧密相关。麻疯树雌花分化与性比表现都与构件营养生长能力紧密相关,麻疯树性比与构件大小的多项指标呈现二次多项式关系,表明了极为强烈的构件大小依赖性(何亚平等,2008,2009a)。同时,麻疯树雌花分化与营养物质的净配置有关,是营养物质配置在自身营养器官构建与生殖器官发育二者之间权衡的结果。在群体水平上,作为单性花同株同花序植物麻疯树雌花分化与性比状态显然会受到群体营养环境的影响,尤其是决定光合作用的光照条件。在遮光条件下,生物柴油植物麻疯树雌花分化可能会受到光照因素的影响,并且这种影响是通过光照对营养器官的塑造来实现的。
对于麻疯树群体而言,度量群体生长特征的基本指标就是群体构件大小与相关的构件性状了,如构件直径、长度等,而群体生殖性状则主要体现在生殖器官上,如雌花数量与性比等。雌花或雌性器官由于在雌性适合度实现中作用明显,从而具有较长的生理寿命,雄花在实现雄性适合度——花粉传递后迅速萎蔫而丧失生活力。生殖是营养生长达到一定阶段的产物,雌花或雌性器官发育状况最能体现资源状况,资源较为充沛,雌花发育较为强烈,性比较高,否则雌花发育较为一般,性比较低。植物对雌性的保障需要用足够的能量来维持雌性器官受精后胚胎的发育,乃至种子成熟,足以显示雌花发育背后蕴藏的巨大资源供给潜力。从这一方面来看,雌花发育自身的数量指标表征了植物自身的资源状况。理论推理,在光照资源限制时,群体光照资源不足则光合能力受到限制,使得营养物质合成不足,营养器官构建受到限制,从而生殖器官发育尤其是雌性生殖器官也受到制约,可能会出现雌花数量减少与性比降低的现象。这一推理得到了数据的支持。倮果桥林下麻疯树群体雌花数量相对较少,性比相对较低,这就是在遮光环境中整体无机资源供给不足而导致生殖器官发育不良的结果。遮光环境中雌花数量较低(2.7 vs 5.04),其本身在雌花受精后胚胎发育中耗费资源较少,是对光照资源限制环境的一种适应对策。麻疯树为腋生聚伞花序,雌花与雄花具有明显的位置配置,雌花位于有限花序顶端,而雄花位于无限花序顶端,并且在雌花与雄花位置转换时呈现出资源充足发育为雌花而资源不足时发育为雄花的特点(何亚平等,2008)。遮光环境中麻疯树性比相对较低(0.027 8 vs 0.037 8),表明部分雌花位点发育不良或者为雄花所代替,从而降低了雌花比例或性比,性比本身也是资源状况的一种表征指标。分析麻疯树构件两性配置认为,麻疯树性比样地之间差异可能是资源限制所致(何亚平等,2009),光照资源限制就是较为普遍的一种,如乔木遮荫、坡向弱光、大密度遮荫、异种大个体遮荫等等,这都造成了局部构件雌雄花发育的光照资源限制,其结果就是导致雌花发育不良,使得雌花数量降低,性比降低。表明,在群体缺乏资源或者资源限制时,两性配置都会呈现雌性不良而雄性相对较旺的配置格局,使得雌性适合度较低而雄性适合度较高,成为花粉输出的源地,而雌性适合度相对较低,种子产量也随之降低。光照限制时,雌性适合度降低,以适应有机营养不足导致的雌性适合度保障不力的无机资源现状。显然,在光照限制时,麻疯树的雌性适合度明显降低(雌花减少,性比降低),这是一种对群体无机资源环境的一种生态适应策略。
林下与林外雌性生殖特性的相对现状必然与营养器官特性相对影响,在生殖器官发育的构件特性上也有相对应的生长发育表现。遮光环境中,由于光照的减弱而使得叶片光合能力不足,绞股蓝的相对生长速率、净同化速率和总生物量随光照强度减弱而降低,总叶面积、比茎长、株高、分枝角度、叶面积率和比叶面积却增加(何维明,钟章成,2000)。在构件水平上,遮光也使得蒙古栎构件高生长明显减弱,而强光下使得高生长存在明显的次生生长现象(樊后保,王义弘,1992)。在个体水平上,红松遮光条件下个体高度生长减弱,尽管其为耐荫性,但光照较强时高生长加快(刘传照等,1991)。麻疯树为喜光植物,光照饱和点较高,补偿点也较高,遮荫时麻疯树光合作用受到影响,出现适应性的生理性变化,光合能力降低(戴凌峰等,2008)。遮光降低了其光照强度,使得麻疯树在低光照强度下营养生长受到限制,而使得光合产物积累降低,除了在生殖器官配置资源以适应生殖生长外,主要把光合营养配置到茎上构建身体,并为将来营养器官发育与生殖器官发育储备资源,显然这种储备会按照比例或者加权而降低。一般而言,光环境限制时,植物生长会出现节间延长而直径降低的可塑性反应。进一步研究还表明,前者限制因素出现时,会降低对后续出现的限制因素,即植物茎出现节间延长时,对后续的昆虫捕食反应不明显(Kurashige and Agrawal,2005)。麻疯树构件出现光照胁迫时,构件自身相对较细(0.811 2 vs 0.912 5),长度缩短 (8.325 5 vs 18.548),而林外则呈现明显的正常直径生长与长度效应。对比林外、林外,麻疯树构件对光照反应更为明显的是枝长,而林外枝长是林内构件枝长的 2倍多!观测表明,麻疯树枝条直径生长较为缓慢,一般而言,当年生枝条与头年生枝条直径仅差 1 mm~2mm,直径生长在紧邻年际间直径生长差异不明显,这与麻疯树为节律性生长的构型特性有关(陈波等,2002)。其营养生长量差异主要体现在长度生长上,显然麻疯树林内林外长度生长效应极为明显本身就体现了营养生长量的实质性差异。而麻疯树在光照限制环境中,枝条出现了对光照的可塑性反应,而后续的生殖生长则也明显无法维持较高的水平,理论上也会使得生殖生长呈现比例性或加权性的降低。关于林外与林内麻疯树生殖性状的适应性反应在前面一段已经得到阐明,生殖性状与营养性状自身的状况极为吻合,表明生殖性状的可塑性反应与营养器官的可塑性反应紧密相连,或者说群体光环境限制对生殖性状的影响是通过影响营养生长性状来实现的,光环境自身无法直接作用于生殖器官,生殖器官是营养生长的副产物(如下图 3所示)。
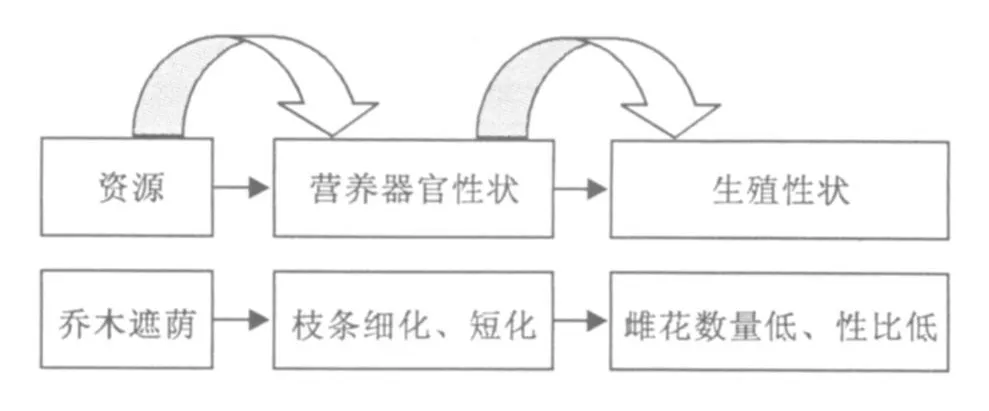
图 3 麻疯树雌花分化的光照调控机制
麻疯树为单性花同株同花序植物,性比与雌花分化在构件水平上就得以体现,这一点与单性花同株异花序植物明显不同,其雌花分化与性比在个体水平上才得以体现。也与雌雄异株植物截然不同,其雌花分化与性比需要在种群水平上界定。显然,麻疯树性表达特点为研究生殖性状的生态适应带来方便。同时,麻疯树自身喜光、耐热,对光照强度较高、温度较高的热带河谷地区较为“偏好”,在个体与构件水平上的营养生长对光照环境也较为敏感,可塑性反应强烈。对光照环境强烈的适应性反应与麻疯树自身的生物学特性有关。考虑雌花分化与构件之间的联系,其本身就是雌花分化与性比的构件大小依赖性,这在文献(何亚平等,2008,2009a)中已经得到初步的阐明。倮果桥麻疯树林外与林内中种群光照强度不同,林下群体雌性器官分化能力降低,与弱光照使得枝条变小,小构件的相关生殖生长器官也小,使得雌雄花数量减小,质量降低,表现为雌花数量低、性比较低。从本质上考虑,小构件生殖器官相应变小是相关生长理论在起作用,即构件大小与生殖功能之间可能存在幂函数关系。小构件自身的光合同化能力、蒸腾拉力、水分和养分输导能力、养分输送能力等都出现比例性或加权性降低,使得生殖器官也出现类似的反应。构件是麻疯树生长发育的触手,是捕获光能,利用光能的反应器,光照对雌花分化与性比的影响是通过降低光合作用而减少营养器官物质构建与积累,进而产生生殖效应的。
对于麻疯树雌花数量与性比而言,主要体现为频度分布,而无绝对统一的数值,这一点在文献(何亚平等,2009a)中也得到体现。一个构件对应一个雌花数量和性比,而其本身由于构件大小紧密相关,表明性比与雌花数量的差异就是构件大小差异所致,其频度或变异性本身与构件的大小、分布格局有关。在群体水平上,构件位置层次不同、方位不同,接受的光照环境不同,物质同化能力不同,雌花分化与性比也不同。空间位置使得光照环境不同,导致了构件自身的雌花分化能力呈现出差异,这就是影响生殖性状的结构限制,即构件分布的位置性因素使得雌花分化能力不同。结构限制背后一般都蕴藏着资源限制,而资源限制发生的实质则是构件的大小依赖性,即构件大小之间存在物质输导、光合同化与光能利用上的差异。大小依赖性可能是环境调节生殖性状的实质性解释工具。统计上考虑,林外与林内构件直径、头年生枝长的频度差异实质体现在大小构件频度上,雌花数量与性比也是如此。总体上就是光限制时,小构件频度较大,低雌性数量构件较多。从统计上考虑,遮光降低了大构件的频度,降低了雌性器官分化能力,使得小雌花数量与性比的频度提高。任何性状主要表现为频度,这是数据表述的一种方法,这种频度自身就是生态因素时空作用的变异性产物,需要引起重视。一些研究表明,林地多年生植物雌性适合度也随着光照强度的增加而增加(Gregg,1975,1978);林地一年生植物在光照强度增加的环境中也存在明显的雌性偏向的两性配置(Cid-Benevento,1987)。本文研究木本植物麻疯树雌花数量与性比随着光照的减弱而降低,表明在弱光环境中生殖潜力降低。基于麻疯树单性花同株同花序的性表达特点,对性比的揭示较为容易,同时构件大小容易度量,使得性比与大小关系很容易就得到了阐明。显然,性比与雌性适合度研究可知,其本身对环境可塑性研究较为容易探讨,这是一个很传统的命题,而对于大小在塑造性比与雌性适合度中的作用则很难。而一旦抓住大小依赖性这一本质则性比和雌性适合度则很容易得到认识清楚。大小依赖性在生殖性状塑造中的作用可能还会引起生态学领域的普遍关注,其本身在生活史理论中的地位有待阐明。同时,本文研究再一次强调了植物生理学中的一个古老命题,即营养器官与生殖器官的相关性问题,需要从系统整体上把握研究问题,而不是孤立看待事物,从而揭示相互联系事物的本质或本性及其维持机制。
在生产实践中,麻疯树生物柴油原料林的高产稳产经营可以避免光照限制,需要在斑块中进行强光经营,避免乔木与异种灌木对麻疯树的遮光作用,并调节个体密度与构件密度,使得雌花发育与雌性适合度表达的光照限制解除,保证雌花良好发育的光照环境。同时,生产中需要明确营养生长与生殖生长的关系,以及种子生产依赖的雌性适合度表达的大小依赖性,来促进营养生长,通过较强的营养生长来保证生殖生长,提高雌花分化能力和性比,为提高种子生产打基础。此外,营养系统与生殖系统为物质生产与输出的关系,促进雌花分化与性比提高的技术措施能够促进物质生产系统的物质源向生殖系统汇转移,从而形成拉动光合作用、提高资源利用效率,进而提高麻疯树雌性的生殖产量的调节措施。
农林复合经营具有较高的资源利用效率,并且在经营中可以以耕代抚,间接提高麻疯树经营管理水平,进而促进种子的高产与稳产。但,林分地表弱光环境使得光补偿点较低,、耐受(或喜好)弱光的矮杆作物高产获得可能,喜光植物麻疯树农林复合经营的林农间作型需要考虑喜弱光植物的选育和模式配置,以保证山地麻疯树“林油一体化”、“林药一体化”与“林农一体化”林业生态产业体系构建,并保证生物柴油原料培育的高产与稳产。需要研究光能利用与产量最大化的密度问题、策略问题与实践可行性问题,这些问题的实质都是麻疯树构件与个体水平上雌性适合度最大化表达问题,雌花分化能力的提高问题。显然,乔木麻疯树混交林是不可取的,而透光度提高则利于性比的提高和雌花分化。
[1] Bazzaz F A,etal.Contribution to reproductive effort by ohotosy nthesis of flowers and fruits[J].Nature,1979,279:554~555.
[2] Carmen R.Cid-Benevento.Relative effects of light,soilmoisture availability and vegetative size on sex ratio of twomonoeciouswoodland annual herbs:Acalypha rhomboidea(Euphorbiaceae)and Pilea pumila(U rticaceae)[J].Bulletin of the Torrey Botanical Glub,1987,114(3):293~306.
[3] Conn JS,Blum U.Sex Ratio of Rumex hastatulus:The Effect of Environmental Factorsand Certation[J].Evolution,1981,35(6):1108~1116.
[4] Costich D E.Gender Specialization Acrossa Climatic Gradient:Experimental Comparison of Monoecious and Dioecious Ecballium[J].Ecology,1995,76(4):1036~1050.
[5] Gregg K B.The effect of light intensity on sex expression in species of Cycnochesand Catasetum(Orchidaceae)[J].Selbyana,1975,1:101~113.
[6] Gregg K B.The interactionsof light intensity,plant size,and nutrition in sex expression in Cycnoches(Orchidaceae)[J].Selbyana,1978,2:212~223.
[7] Jean-Michel G,Daniel F.Environmental Sex Determination in Response to Light and Biased Sex Ratios in Equisetum gametophytes[J].The Journal of Ecology,2003,91(1):49~57.
[8] Kurashige NS,Agrawal A A.Phenotypic plasticity to lightcompetition and herbivory in Chenopodium album(Chenopodiaceae)[J].American Journal of Botany,2005,92(1):21~26.
[9] Lloyd DG.Breeding Systems in Cotula L.(Compositae,Anthemideae).II.Monoecious Populations[J].New Phytologist,1972,71(6):1195~1202.
[10] Openshaw K.A review of Jatropha curcas-an oil plant of unfulfilled prom ise[J].Biomass and Bioenergy,2000,19(1):1~15.
[11] 陈波,宋永昌,达良俊.木本植物的构型及其在植物生态学研究的进展[J].生态学杂志,2002,21(3):52~56.
[12] 陈绍云,周国宁.光照强度对山茶花形态、解剖特征和生长发育的影响[J].浙江农业科学,1992,(3):144~146.
[13] 窦新永,吴国江,黄红英,等.麻疯树幼苗对干旱胁迫的响应[J].应用生态学报,2008,19(7):1425~1430.
[14] 樊后保,王义弘.不同光照条件下蒙古栋物候期及树高生长节律的研究[J].福建林学院学报,1992,12(2):148~153.
[15] 何维明,钟章成.攀援植物绞股蓝幼苗对光照强度的形态和生长反应[J].植物生态学报,2000,24(3):375~378.
[16] 何亚平,费世民,蔡小虎,等.攀枝花市麻疯树生殖构件特征与雌雄花配置研究[J].四川林业科技,2009a,30(1):8~17.
[17] 何亚平,费世民,蔡小虎,等.麻疯树构件的生殖配置研究[J].四川林业科技,2009b,30(3):1~7.
[18] 何亚平,费世民,徐嘉,等.四川麻疯树花序结构和雌雄花动态研究[J].四川林业科技,2008,29(2):1~8.
[19] 黄红英,窦新永,孙蓓育,等.两种不同生态型麻疯树夏季光合特性的比较[J].生态学报,2009,29(6):2861~2867.
[20] 廖咏梅,雷泞菲,陈劲松.异质性光照条件下甸甸茎草本野草蓦的克隆整合[J].广西植物,2006,26(5):503~506.
[21] 刘传照,李俊清,金奎刚.林下光照条件与红松幼树生长的相关性研究[J].东北林业大学学报,1991,19(3):103~108.
[22] 吴载璋,陈绍栓.光照条件对楠木人工林生长的影响[J].福建林学院学报,2004,24(4):371~373.
[23] 夏亮,邢杰,王胜华,陈放.麻疯树光合生理特征研究[J].四川大学学报(自然科学版),2008,45(2):413~418.
[24] 肖春旺,刘玉成.不同光环境的四川大头茶幼苗的生态适应[J].生态学报,1999,19(3):422~426.
[25] 闫兴富,曹敏.不同光照梯度的遮荫处理对绒毛番龙眼幼苗生长的影响[J].热带亚热带植物学报,2007,15(6):465~472.
[26] 尹春英,李春阳.雌雄异株植物与性别比例有关的性别差异研究现状与展望[J].应用与环境生物学报,2007,13(3):419~425.
[27] 张教方,刘晓东,于海滨,等.毛百合繁殖生物学研究亚光照梯度上的形态性状差异与繁殖对策[J].植物学通报,1993,13(3):285~293.
[28] 张诗莹,樊卫国.麻疯树的光合特性[J].种子,2005,24(8):13~15.
[29] 张效平.光照强度对唐菖蒲花芽发育的影响[J].南京农业大学学报,1990,13(4):35~38.
[30] 贾治邦.麻疯树实用栽培技术[M].北京:中国林业出版社,2008.
[31] 戴凌峰,崔令军,张志翔.遮阴处理对小桐子幼苗生长的影响[J].安徽农业利学,2008,36(14):5729~5731.
猜你喜欢
阅读与作文(小学低年级版)(2018年9期)2018-10-26
阅读与作文(小学低年级版)(2018年10期)2018-10-25
农业知识(2018年18期)2018-05-28
妇女之友(2017年3期)2017-04-20
创新作文(3-4年级)(2016年9期)2016-12-22
中国药物应用与监测(2015年5期)2015-12-11
意林·少年版(2009年1期)2009-12-29
北方人(2009年12期)2009-02-01
中学英语之友·上(2008年11期)2008-12-08
