
反刍动物甲烷生成机制及调控
2010-01-30 01:32董国忠
中国草食动物科学 2010年2期
苏 醒,董国忠
(西南大学动物科技学院,重庆 400715)
甲烷(CH4)是一种重要的大气微量成分。它与CO2一样,是一种长寿命的温室效应气体。研究者普遍认为甲烷的温室效应是二氧化碳的20~30倍。甲烷对全球气候变暖的影响作用占到所有影响气候变暖因素作用的15%~20%[1]。联合国政府间气候变化专业委员会(IPCC)的报告中指出,全球每年排入大气的CH4量是 535 Tg(410~660Tg)(1Tg=1012g),而来自动物,主要是反刍动物的达85Tg(65~100Tg),对全球温室效应的贡献率达2%[2]。我国农业部环境保护科研监测所对我国家养动物和动物废弃物甲烷排放量的估算结果是,甲烷排放量平均每年大约以2.34%的速度上升[3]。
反刍动物以嗳气的形式排出体外的甲烷主要是由瘤胃中甲烷菌通过CO2和H2进行还原反应产生的,化学性质稳定,一旦发生后很难被代谢。在反刍动物瘤胃代谢过程中,瘤胃内甲烷的产生是瘤胃发酵能量损失的主要原因,依据饲养水平、饲粮组成、消化率的不同,以甲烷形式损失的能量占饲料总能的 2%~15%[4]。因此,降低瘤胃内甲烷的产量不仅对缓解温室效应有重要意义,而且可减少反刍动物瘤胃发酵过程中的能量损失,提高饲料利用率。减少反刍动物CH4排放量是国际上十分关注并重点研究的问题。本文综述了瘤胃甲烷生成的机制、影响瘤胃甲烷产生的因素和控制瘤胃内甲烷产生量的措施。
1 瘤胃甲烷生成的机制
反刍动物生成甲烷与其特有的消化特点有关。饲料在被动物吞食后首先在瘤胃内进行厌氧发酵,瘤胃内的微生物把碳水化合物和其它植物纤维发酵分解成挥发性脂肪酸(VFA)、氢气和二氧化碳等。产生的二氧化碳、甲酸、乙酸、甲胺和次甲胺等在甲烷生成菌的作用下生成甲烷。从瘤胃中已鉴定出的甲烷生成菌主要包括甲烷短杆菌、甲烷微菌、甲烷杆菌和甲烷八叠球菌等4属。甲烷的产生途径主要有以下3种:

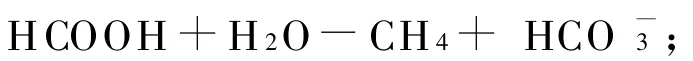
2 影响瘤胃甲烷产生的因素
研究表明,在供给相同饲料的情况下,虽然品种、年龄和性别不同,但动物的甲烷排放量相似,从而说明反刍动物甲烷排放量主要受碳水化合物类型、采食量、环境温度等多种因素的影响,与品种、性别、年龄的关系不大[5]。
2.1 碳水化合物类型 甲烷的生成受饲料发酵类型的影响,主要通过改变pH值和瘤胃微生物区系而产生。当反刍动物采食富含淀粉的日粮或一定剂量的可溶性碳水化合物时,丙酸产生量增加,甲烷产生量降低,且乙丙酸比例也随之降低。这主要是由于可溶性碳水化合物或淀粉摄入瘤胃后,瘤胃pH值降低,从而抑制了粗饲料的消化和原虫、甲烷菌的活动。这种瘤胃发酵模式由于瘤胃降解率提高,有利于丙酸的生成。当给动物饲喂粗饲料时,纤维素分解菌大量增殖,瘤胃主要进行乙酸发酵,产生大量的氢,瘤胃氢分压升高。这时就会刺激甲烷菌大量增殖,甲烷产量增加。研究发现,当瘤胃发酵的乙丙酸比为0.5时,不产生甲烷,即饲料能的损失为0;而当碳水化合物发酵全部生成乙酸时,以甲烷的形式损失的能量约为33%[6]。
2.2 瘤胃VFA对甲烷产生量的影响 瘤胃内挥发性脂肪酸与甲烷产生量有一定关系。研究表明,丙酸产量与甲烷产量呈较高的负相关,乙酸产量和乙酸与丙酸比例同甲烷产生量呈较高的正相关[7]。这在另一方面也反映了不同日粮对甲烷产量的影响。低质粗料由于乙酸含量高,会导致甲烷产生量高;增加日粮精料比例,丙酸含量提高,因此甲烷产生量降低。
2.3 环境温度 反刍动物瘤胃甲烷产量随着环境温度的降低而降低。在寒冷的条件下,绵羊瘤胃甲烷的产量会降低30%,这是由于随着温度的降低,瘤胃发酵更趋于丙酸发酵,引起甲烷产量的降低[8]。也有人认为,温度的降低也可能会提高动物食糜的后送速度,导致甲烷产量的降低。
2.4 采食量 提高动物的采食量,因甲烷损失的日粮总能可下降到1.6%[9]。还有研究也证实了动物采食量提高,甲烷产量下降[10]。当采食量提高到维持水平的2倍时,总的甲烷产量上升,但是单位摄入能量的甲烷损失下降12%~30%[11]。
2.5 饲料的加工和贮存 牧草的成熟期、贮存方式、化学处理和物理加工过程(如切碎、碾碎、制粒)都影响每单位可消化牧草的甲烷产量。甲烷的产生量随牧草成熟期的延长而增加。同时牧草制干比青贮产生更多的甲烷量。粗制切碎牧草比细致碾碎或制粒的牧草损失更多的甲烷能[12]。这主要是由于动物自由采食这些小粒饲料加快了瘤胃内容物流通速率,降低了微生物对细胞壁碳水化合物的消化率,从而降低了甲烷产量。因此,当饲草具有降低消化率或延长瘤胃饲料残渣滞留时间的性能时,一般来说会增加每单位可消化饲草的甲烷产量。
3 瘤胃甲烷生成的调控
适量地降低反刍动物甲烷的生成和排放,具有经济和环境双重效益,为此,研究人员探索了种种方法。一是通过育种和营养措施来提高单位动物的生产性能,减少动物存栏数量,从而降低甲烷气体的总排放量;二是通过对瘤胃甲烷菌生成甲烷过程进行调控,这是当前研究的重点。甲烷生成的调控机理主要有3种模式:一种是生物学调控,即直接抑制产甲烷菌的生长,减少甲烷菌的数量,从而减少甲烷的生成;另一种是通过减少生成甲烷的底物——H2生成量或通过替代性受体争夺H2而减少甲烷生成量;再一种途径是通过特异性抑制甲烷菌合成甲烷途径中的某些酶的活性而抑制甲烷的生成。
3.1 生物学调控
3.1.1 抗生素 莫能菌素和盐霉素等聚醚类离子载体抗生素能通过影响细胞膜通透性、改变微生物代谢活动而抑制产甲烷菌、产氢菌和产甲酸菌以达到减少甲烷产量的目的,同时它们还有利于产琥珀酸菌和丙酸菌,结果增加丙酸产量,提高饲料利用率。据报道,这类离子载体类抗生素能减少25%的甲烷生成,同时提高饲料转化效率[13]。然而,长期使用抗生素可出现细菌耐药性而导致效果不理想,并且畜产品中残留的抗生素将对人体健康造成威胁。
3.1.2 驱除原虫 大量研究表明有20%的产甲烷菌寄生在原虫表面,并且带纤毛的原虫能与产甲烷菌形成内源共生系统,因此,驱除原虫可间接减少产甲烷菌数量,进而显著降低反刍动物甲烷的排放量。反刍动物的除虫方式有:①日粮调控:瘤胃低pH值可导致原虫死亡,突然换成或延长全谷物日粮以降低瘤胃pH值,从而可达到驱虫目的。另外,植物油、乳脂、C18不饱和脂肪酸等对原虫具有毒性,故也可用来驱除原虫。研究表明,肉牛日粮中添加400g/d的葵花籽油,甲烷的排放减少了22%,总能的损失减少了21%[14]。②化学合成药品:自从1930年Becker等首次用硫酸铜驱除原虫以来,过氧化钙、硫代琥珀酸二辛基钠盐和去污剂相继被用来驱除原虫。③天然化合物,如维生素A、蜕皮激素、非蛋白性氨基酸及其类似物都能并已被用作瘤胃除原虫试剂。研究发现,皂角苷对原虫有很强的毒性,单宁具有杀灭寄生虫的作用[5]。研究发现,日粮中添加缩合单宁能减少山羊甲烷的排放[15]。
3.1.3 其他方法 一些卤族化合物可直接抑制产甲烷菌,其中碘化合物效果最好,但长期使用微生物会产生适应性;此外,甲羟戊二酸单酰辅酶A还原酶的抑制剂-Mevastatin和Lovastatin也是甲烷菌的抑制剂,它们只抑制甲烷菌的生长,而对其他细菌无影响,在减少甲烷生成同时增加了饲料利用率[9]。
3.2 对甲烷底物氢气的调控
3.2.1 抑制氢气的生成 减少瘤胃发酵的有机物和调节生成乙酸与丙酸之间的平衡均可降低氢气的生成。因此,一些能提高瘤胃流通速率和增加过瘤胃率的饲料加工措施和饲喂方法可在一定程度上减少反刍动物甲烷的生成。可以通过适当的日粮加工和配合,使瘤胃发酵过程中乙酸与丙酸的比例尽可能接近0.5,从而降低甲烷的生成。离子载体抗生素莫能菌素除可抑制产甲烷菌本身外,还可降低瘤胃中氢气的生成。其主要机制在于莫能菌素可抑制甲酸脱氢酶的活性,从而减少了甲烷产生所需要的氢源,使得甲烷产量降低;此外,莫能菌素还可选择性降低发酵过程中乙酸生成的比率。
3.2.2 添加氢气竞争性受体 添加氢气竞争性受体,如硝酸盐或硝酸盐与半胱氨酸的混合物、不饱和脂肪酸等,与产甲烷菌竞争氢气以抑制甲烷菌的生长。研究发现,粗饲料作物中含有硝酸盐时能显著降低瘤胃甲烷产量,但硝酸盐还原产生的中间产物亚硝酸盐对瘤胃微生物和动物都有毒性,限制了其作为甲烷抑制剂的使用[15]。试验结果表明延胡索酸能在不影响纤维消化的情况下有效抑制甲烷生成[8]。
除甲烷菌可利用瘤胃发酵过程中产生的氢气外,乙酸菌也可利用氢气为营养物质来合成乙酸。由于乙酸是可被反刍动物利用的主要养分之一,而且乙酸菌与产甲烷菌的竞争性利用氢气是一个自然、安全的过程,不会对动物健康及其产品安全造成威胁,因此一些学者希望能通过建立有利于乙酸菌的瘤胃微生态新平衡,进而减少甲烷生成[10]。
3.3 抑制甲烷合成相关酶的活性 蒽醌类物质(Anthraquinone,AQ)直接作用于甲烷菌,阻断电子传递链,并在电子传递和与细胞色素有关的ATP合成的藕联反应中起解藕联作用,从而阻止CH3-CoM被还原成甲烷。体外研究证实了添加蒽醌能明显降低甲烷的产量[14]。四氢甲烷蝶呤(tetrahydromethanopterin)是产甲烷菌体内合成甲烷的关键辅助因子,RFA-P合成酶催化四氢甲烷蝶呤合成的第一步反应,体外试验表明,添加RFA-P合成酶抑制剂降低了甲烷的生成,而对VFA的生成和乙酸菌的生长没有影响[6]。
4 结束语
牛羊嗳气中的甲烷不仅能损失能量,而且会加剧温室效应,促进气候变暖。但甲烷是瘤胃碳水化合物发酵不可避免的产物。随着对瘤胃甲烷生成机制的认识,对影响甲烷产量因素的了解,已经找到了一些行之有效的甲烷生成量的调控方法。然而反刍动物甲烷的生成是一个复杂的过程,甲烷菌对于维持瘤胃微生态的正常具有重要意义。试验证明,甲烷菌的存在能保持瘤胃内低水平的H2分压,提高其他微生物特别是分解纤维素微生物的发酵能力。甲烷的生成是反刍动物进化过程中形成的维持其瘤胃微生态平衡的有效机制。甲烷菌作用的多向性使得对甲烷生成的调控变得很复杂。如何选择既可降低甲烷的生成又不影响动物的健康和生产性能,是今后的重要任务。随着科学技术的发展与科研工作者的探索,相信人们一定会找到对环境、对动物健康及其产品安全都有益的有效调控甲烷产量的新方法。
[1]孙维斌,胡建红.反刍动物瘤胃甲烷的产生与调控研究进展[J].黄牛杂志,1999,25(6):37-39.
[2]IPCC:Intergovern mental panel on climate change.T he supplementary report to the IPCC scientific assessment[C].New York:Cambridge University Press,1992.
[3]沈跃.黄牛品种改良与减排甲烷潜力的研究[J].农业环境保护,1995,14(1):15-17.
[4]Holter,Yang J.M ethane production in dry and lactating Holstein cows[J].J Dairy Sci,1992,75:2165.
[5]赵玉华,杨瑞红,王加启.反刍动物甲烷生成的调控[J].中国饲料,2005(3):18-20.
[6]Miller T T,Wolin M J,Zbao Heta1.Characteristics of methanogen isolated from ovine rumen[J].App1 Environ Micr,1988,51:201-202.
[7]韩继福,冯仰廉.阉牛不同日粮的纤维消化、瘤胃内VFA对甲烷产生量的影响[J].中国兽医学报,1997,17(3):278-280.
[8]Ferry G G.Biochemistry of methanogenesis[J].Mo1 Bil,1992,27:473-503.
[9]Johnson D E,T M Hill,B R Carmean,et al.Energy metabolism of farm animals[J].J Aim Sci,1991,20:376-379.
[10]Okine E K,Mathison G W,Hardin R T.Effect of changes in frequency freticularon tractioli on fuidadn particulate passageratesia[J].Anim Sci,1989,67:3388-3396.
[11]Blaxter K L,J L Clapperton.Prediction of the amount of methane produced by ruminants[J].Br J Nutr,1965,19:511-519.
[12]Kalmakoff M L,Bartlett F,Teather R M.Are ruminal bacteir armed with bactericoins[J].J Dairy Sci,1996,79:2297-2306.
[13]Johnson K A,Johnosn D E.Methane emissions from cattle[J].J Aim Sci,1995,73:2483-2492.
[14]T edeschi L O,D G Fox,T P Tylutki.Potential environmental benefits of ionophores in ruminant diets[J].J Environ Qual,2003,32:1591-1602.
[15]韩继福.反刍动物瘤胃内甲烷产生及其调控[J].当代畜牧,1996,20(4):8-l1.
猜你喜欢
中国感染控制杂志(2022年11期)2022-11-24
吉林畜牧兽医(2022年6期)2022-11-16
养殖与饲料(2021年6期)2021-11-30
现代畜牧科技(2021年4期)2021-07-21
新农业(2020年18期)2021-01-07
中国动物保健(2020年6期)2020-11-27
科学(2020年2期)2020-08-24
智富时代(2019年5期)2019-07-05
智富时代(2019年5期)2019-07-05
中国兽医杂志(2019年2期)2019-06-25
