
淡水壳菜在深圳市原水管道中的繁殖规律
2009-04-28 07:48罗凤明张重祉胡向萍
科技经济市场 2009年3期
罗凤明 张重祉 胡向萍
摘要:本文研究了淡水壳菜在深圳市原水管道中的繁殖生境、雌雄性比、受精方式及个体发育和繁殖期。结果表明,优势雌雄性比约为2:1,各月份的优势雌性维持在65%左右;有体内受精和体外受精两种受精方式;个体发育经历了卵裂、囊胚期、原肠胚期、担轮幼虫、面盘幼虫和稚贝;繁殖期2月~9月,最佳繁殖期3月~7月。
关键词:淡水壳菜; 繁殖规律
1材料与仪器
实验材料:采自东湖水厂格栅井的淡水壳菜
主要仪器:生物显微镜XSZ-HS1型重庆光电仪器有限公司 体视显微镜SZM-45T3型 重庆光电仪器有限公司
2实验方法
2.1采样方法
(1)每月定期用定制的长柄刮网从电站水泥池壁上刮取淡水壳菜;
(2)之后,在有流动原水的水泥池中驯养2-3天,以备实验。
2.2性征观测方法
(1)每月随机挑选200个淡水壳菜进行解剖,用体视显微镜观察其性腺发育程度和内部结构的变化;
(2)在生物显微镜下进行性腺、外套膜涂片观察,辨别雌雄,计算每月的雌雄性比。
2.3受精方式观测方法
(1)用生物显微镜观察雌体鳃间腔中的胚胎形态及其数量变化;
(2)计算出每月具胚胎的雌体在总样本中所占比例;
(3)实验室观察淡水壳菜在原水中的排精放卵和发育过程。具体做法是:a、选贝。取已驯养的亲贝12~15个,阴干12h,去足丝和壳面污物,经原水洗净后投入盛有2.5~3.0L原水的玻璃缸中,让它排泄粪便和污物,并及时更新原水。发现排放的个体,立即捡出置于玻璃缸中。b、观察。在显微镜下观察精子、卵细胞、受精卵、卵裂、担轮幼虫、面盘幼虫及幼体的出现。
3结果
3.1生境
原水管道水流速为0.1-2.5 m/s利于淡水壳菜附着生长;管道表面的粗糙度越大,浮游生物和有机碎屑等营养物质越丰富,附着密度越大;光照对淡水壳菜的生长有抑制作用;水温也会影响淡水壳菜的生长,适宜水温16℃~28℃。
3.2雌雄性比
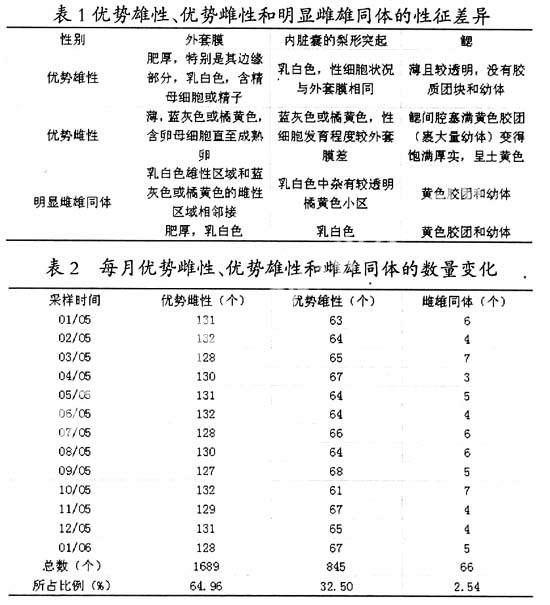
解剖发现,深圳采集的淡水壳菜基本上属于雌雄同体类型。即同一个体中,有潜在的卵巢组织和潜在的精巢组织,绝大多数个体(约2/3)的卵巢部分远比精巢部分发达,掩盖了雄性组织,称之为优势雌性个体;而较少的个体相反,精巢部分发育得更好,遮盖了雌性组织,称之为优势雄性个体;而极少数个体的卵巢和精巢都发育较好,以致雌雄组织同时显现出来,称为明显雌雄同体个体。表1是优势雄性、优势雌性和明显雌雄同体在生殖季节里的性征差异。每月优势雌性、优势雄性和雌雄同体的数量变化见表2。从表2可知,优势雌雄性比约为2:1,优势雌性在各月份所占的百分率在65%左右。
3.3受精方式及个体发育
3.3.1受精方式
体内受精:解剖繁殖期间淡水壳菜的外套膜,能看到精子、
卵细胞、受精卵、各时期的胚;解剖雌体的鳃间腔,可观察到密生纤毛的担轮幼虫和面盘幼虫。说明精卵细胞在外套膜中成熟,受精作用在外套膜中完成,最后,受精卵在鳃间腔内发育成幼体。由此说明,淡水壳菜可进行体内受精。
体外受精:将阴干的淡水壳菜放入原水中,一段时间后就可观察到亲贝排放精卵,排精放卵持续时间约为2~6h。把含精子和卵细胞的原水混合,之后可观察到受精卵、桑椹胚等。由此表明,淡水壳菜也能进行体外受精。
3.3.2个体发育
(1)生殖细胞与受精
壳长8mm的雄性和6mm的雌性个体就具有繁殖能力。精液呈乳白色烟雾状,精子长约50μm,头部略呈圆锥形,尾部细长。成熟卵子呈浅橙黄色,细颗粒状,大小均匀,呈圆球状,直径约70μm,外包一层胶质膜。成熟的精卵可以在体内外结合成受精卵。
(2)2细胞期至桑椹胚
2细胞期: 受精后5min,受精卵进行第一次分裂;4细胞期:第一次分裂完成5min左右,大卵裂球形成第二极叶。5min后,第二次分裂开始,形成一大三小的四个卵裂球;8细胞期:受精后50min左右,分裂前仍有极叶出现,之后以左螺旋式进行第三次分裂,形成8个细胞;16细胞期:8细胞形成后10min左右,仍以左螺旋式进行第四次分裂,形成16个细胞;32细胞期及桑椹胚:在16细胞期的基础上又向右旋转,分裂形成32个细胞。之后继续分裂,形成桑椹胚。
(3)囊胚和原肠期
囊胚期:受精后约3h进入囊胚期。囊胚期的幼体,其表面布满短小的纤毛,能在水中游动旋转;约在受精后3.5h,动物极细胞下包、植物极细胞内陷,逐渐形成原肠胚。
(4)担轮幼虫
雌体鳃间腔观察到球形胚胎,直径200~250μm,呈淡黄色,能在水中不断旋转。早期担轮幼虫出现在受精后9~10h,胚体渐变圆,梨形顶端膨大,细胞加厚,有顶纤毛束,能借助纤毛摆动作旋转运动,但中央还没有出现粗大的鞭毛,经常游于水表层,胚孔区还没内陷形成口凹。晚期担轮幼虫左右略变扁平,背部尖,腹部宽,顶端扁平,四周细胞隆起,顶纤毛束的中央有一或二根粗大的鞭毛,胚孔区内陷,逐步形成口凹。
(5)面盘幼虫和稚贝
担轮幼虫两天左右就发育成面盘幼虫。面盘幼虫略呈角锥形,角锥的基部向内凹陷,能在水中翻滚旋转。移入培养皿后五六天(水温20℃~22℃)附着变态为稚贝,长出双壳,足逐渐形成,并能伸出壳外爬行。此时,在幼虫壳的边缘长出成体壳,并分泌足丝营附着生活,滤食水中的浮游植物和有机碎屑。
3.4繁殖期
对雌体鳃间腔中的胚胎进行了为期一年(2005年1月~2006年1月)的观察,计算出具胚胎的雌体在200个总样本中所占的比例,并总结了其与全年的月平均水温的关系,结果见图1。

图1表明,鳃间腔中含有胚胎的雌体呈现先升高后降低的趋势,与水温变化的趋势基本一致。淡水壳菜的繁殖期为2~9月,期间的平均水温由15.8℃逐月升高到28℃,其最低繁殖水温接近16℃。最佳繁殖时期是3月~7月,平均水温为16.2℃~27.6℃,具胚胎雌体的百分率为50%~61.9%。
1~4月,鳃间腔中含胚胎的雌体比例逐月增加,性腺发育主要集中在2月,3月开始进入了快速繁殖期;5~6月,鳃中具胚胎雌体的比例有所下降,繁殖速度相对放慢,进入短暂的生理恢复期;7月份具胚胎的雌体比例有所升高,这表明淡水壳菜的性腺是分批成熟;8~10月,淡水壳菜繁殖速度明显降低, 12月才基本结束;性腺约从9月开始萎缩。
4讨论
本文认为淡水壳菜具有体内和体外两种受精方式,而不是单一的受精方式,这与以往的研究结论(Morton,1982;Choi and Kim,1985)有较大不同。
根据淡水壳菜在深圳的壳长生长速度,只需6个月平均壳长可达9.07mm,就能达到性成熟,而不是一年(Darrigran,2004)。淡水壳菜的性腺集中在2月成熟,9月开始萎缩,生殖腺成熟和开始萎缩时间比阿根廷(Darrigran,2004)早,可能与两地的水温有关,这有待于进一步研究。在深圳原水管道中,淡水壳菜具有较长的繁殖期,约8个月(2~9月),且促发淡水壳菜开始繁殖的“门槛”水温为16~17℃,这与Daniel 和 Boltovskoy(2000)、Darrigran 等(1999) 观察结果基本一致。
参考文献
[1]Morton B. The reproductive cycle in Limnoperna fortunei (Dunker 1857)(Bivalvia:Mytilidae) fouling Hong Kong's raw water supply system [J]. Oceanol. Limnol. Sinica, 1982,13 (4) :312 ~ 324.
[2]Choi SS and Kim JS. Studies on the metamorphosis and the growth of larva in Limnoperna fortunei [J]. Korean J Malacol, 1985,1:13~18.
[3]Darrigran G, Maronas M and Colautti D. Air exposure as a control mechanism for the golden mussel, Limnoperna fortunei, (Bivalvia: Mytilidae) [J]. Journal of Freshwater Ecology, 2004,1 ~9.
[4]Daniel H and Boltovskoy D. Yearly reproductive activity of Limnoperna fortunei (Bivalvia) as inferred' from the occurrence of its larvae in the plankton of the lower Parana river and the Rio de la Plata estuary( Argentina ) [J]. Aquatic Ecology , 2000,34:307~317.
[5]Darrigran G, Penchaszadeh P and Damborenea MC. The reproductive cycle of Limnoperna fortunei(Dunker, 1857) (Mytilidae) from a neotropical temperate locality [J]. J Shellfish Res, 1999,18:361~365.
猜你喜欢
当代水产(2022年3期)2022-04-26
供水技术(2022年1期)2022-04-19
现代畜牧科技(2021年7期)2021-07-28
中国动物保健(2021年1期)2021-05-10
小哥白尼(野生动物)(2020年2期)2020-06-16
学生天地(2020年25期)2020-06-01
中国资源综合利用(2017年1期)2018-01-22
水科学与工程技术(2016年1期)2016-07-12
少儿科学周刊·儿童版(2015年11期)2015-12-17
绿色科技(2014年4期)2015-01-06
