
热胁迫对中间球海胆抗氧化酶活性及线粒体结构与功能的影响
2022-07-14 01:23王文配韩泠姝张向磊杨新宇张伟杰常亚青
广东海洋大学学报 2022年4期
王文配,韩泠姝,2,张向磊,刘 鹏,杨新宇,王 荦,张伟杰,常亚青,丁 君
(1.大连海洋大学农业农村部北方海水增养殖重点实验室,辽宁 大连 116023;2.宁波大学海洋学院,浙江 宁波 315211)
中间球海胆(Strongylocentrotus intermediu)生长快、品质优,但对温度变化敏感,2017 年山东、辽宁等地增养殖中间球海胆曾因夏季高温导致大量死亡[1-3]。海洋生物在急性高温胁迫下会调整生理调节机制,线粒体代谢机制的调整对生物补偿温度变化所产生的影响至关重要[4-5]。ATP 酶是一类分布广泛的膜结合蛋白酶,参与生物体能量代谢等生理生化过程。鲤(Cyprinus carpio)和团头鲂(Megalobrama amblycephala)在受到高温胁迫时其肝脏细胞线粒体超微结构受损[6-7];凡纳滨对虾(Litope‐naeus vannamei)和银鲳(Pampus argenteus)在受到高温胁迫时,其ATP 酶活性显著降低[8-9]。但未见对中间球海胆线粒体的相关报道。
此外,高温也会激活生物体液免疫,改变抗氧化酶活性。抗氧化系统中超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、谷胱甘肽过氧化物酶(GSH-PX)和过氧化物酶(POD)等起主要作用,SOD 将生物体产生的有毒物质转化为H2O2,CAT 和GSH-PX 可以将H2O2转变为水和分子氧,以清除氧自由基和保护细胞免受氧化损伤[10]。POD 广泛分布于动物、植物和微生物细胞中,在底栖海洋无脊椎动物中如贻贝(Mytilus edulis)和蛤类,POD是参与免疫调节的主要免疫酶类[11]。在牙鲆(Paralichthys olivaceus)[12]及长牡蛎(Crassostrea gigas)[13]的研究中发现,高温使其体内SOD、CAT等抗氧化酶的活性显著升高,而罗氏南极鱼(Notothenia rossii)[14]在高温状态下其肾脏中CAT 酶活性显著降低;中间球海胆1 ℃/d 从15 ℃升至26 ℃的过程中,SOD及CAT活性呈先升高后降低趋势,该变化可以保护海胆免受氧化损伤[15]。
本研究以中间球海胆为研究对象,开展急性高温胁迫下中间球海胆线粒体结构与功能以及抗氧化酶活性变化特征研究,有助于从生理水平上阐明中间球海胆如何响应高温胁迫,为海胆响应高温胁迫及高温育种提供基础。
1 材料与方法
1.1 实验材料
选取45 只饲养于农业农村部北方海水增养殖重点实验室的1.5 龄健康中间球海胆,体质量为(35.30±1.93)g,壳径为(47.01±5.35)mm,壳高为(28.99±2.80)mm。
1.2 实验设计
实验设计15、20和25 ℃3个温度梯度,以15 ℃为对照组。每个梯度3 个重复,每个重复5 只。实验开始将45 只中间球海胆放置于15 ℃控温水槽内暂养7 d,其中15 只暂养7 d 后立即取样。再将剩余30 只实验用中间球海胆随机分配到2 个控温水槽中,其中15 只以1 ℃/h 速度逐渐升温至20 ℃后,维持24 h 后立即取样;另外15 只以1 ℃/h 的速度逐渐升温至25 ℃后,维持24 h后立即取样。
1.3 样品采集
每个重复随机取3 只,每个实验组9 只,置于冰盒上活体解剖,每只抽取约5 mL 体腔液装入10 mL离心管内,离心(转速9 000 r/min)10 min 后取上清液转入新的离心管内。抽取体腔液后立即剪取肠组织液氮速冻,同离心后的体腔液和沉淀的体腔细胞放置于-80 ℃冰箱内保存备用。
1.4 检测指标
肠组织检测指标是ATP 和Ca2+浓度、Ca2+-ATP酶、Ca2+-Mg2+-ATP 酶、Na+-K+-ATP 酶,体腔液检测的指标是SOD、CAT、GSH-Px、POD。线粒体提取、ATP 浓度测定、Ca2+浓度测定的试剂盒均购自上海杰美基因医药科技有限公司,其具体实验方法见说明书;ATP 酶活测定、抗氧化酶(GSH-Px、CAT、POD、SOD)活性测定的试剂盒均购自南京建成生物工程研究所,其具体实验方法见说明书。超薄切片制备借鉴刘晓辉的实验方法[6],最后使用透射电镜(JEM-2000EX)观察。
1.5 数据处理
实验结果均用平均值±标准差表示。实验数据用SPSS19.0 软件进行统计分析,分别对各组的实验数据进行单因素方差(One-Way ANOVA)分析和Duncan’s多重比较,显著性水平α=0.05。柱状图使用GraphPad Prism v8.0.2.263制作。
2 结果
2.1 急性高温胁迫下中间球海胆体腔细胞线粒体结构变化
对照组(15 ℃)体腔细胞边界清楚(图1(a))。线粒体呈圆形或椭圆形,且在细胞边缘排列,线粒体膜完整,嵴清晰可见且靠近线粒体两端排列(图1(b))。
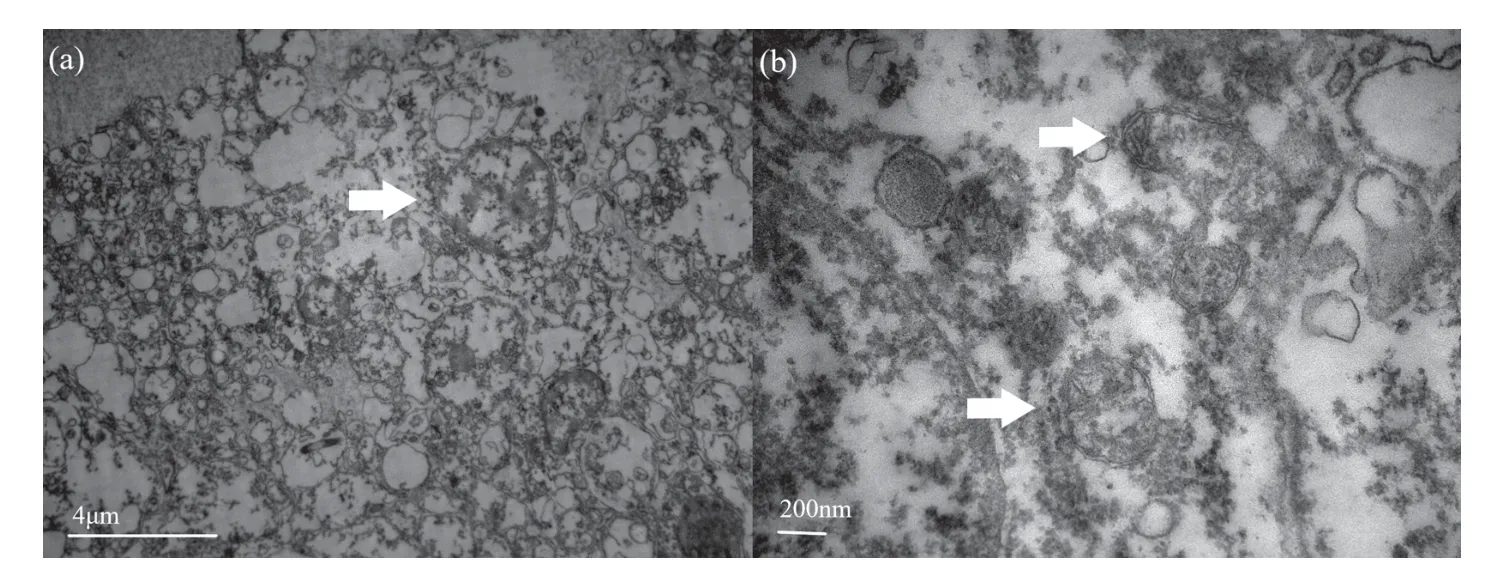
图1 15 ℃下的中间球海胆体腔细胞结构Fig.1 The structure of coelomocytes of the Strongylocentrotus intermedius at 15°C
20 ℃组体腔细胞边界清楚(图2(c)),线粒体呈圆形或椭圆形,在细胞核周围排列,与对照组相比线粒体数目显著增多。极大部分线粒体膜完整且嵴在线粒体中间排列;小部分线粒体膜溶解破损,嵴脱落不清晰,线粒体基质与对照组相比密度增大(图2(d))。

图2 20 ℃下的中间球海胆体腔细胞结构Fig.2 The structure of coelomocytes of the Strongylocentrotus intermediu at 20°C
25 ℃组体腔细胞边界清楚(图3(e)),线粒体呈圆形或椭圆形。线粒体数目与20 ℃组相比数目降低;大部分线粒体膜完整,部分线粒体膜破损,结构破损情况与20 ℃组相似。嵴破损溶解脱落在线粒体基质中浑浊不清晰,线粒体基质密度比20 ℃组大(图3(f))。

图3 25 ℃下的中间球海胆体腔细胞结构Fig.3 The structure of coelomocytes of the Strongylocentrotus intermedius at 25°C
2.2 急性高温胁迫下中间球海胆肠组织线粒体功能变化
图4显示,随温度升高,ATP浓度变化不显著(P>0.05)。随温度升高至20 ℃,Ca2+浓度显著下降(P<0.01);当升温至25 ℃,与20 ℃组相比Ca2+浓度变化不明显(P>0.05),与对照组相比差异极显著(P<0.01)。
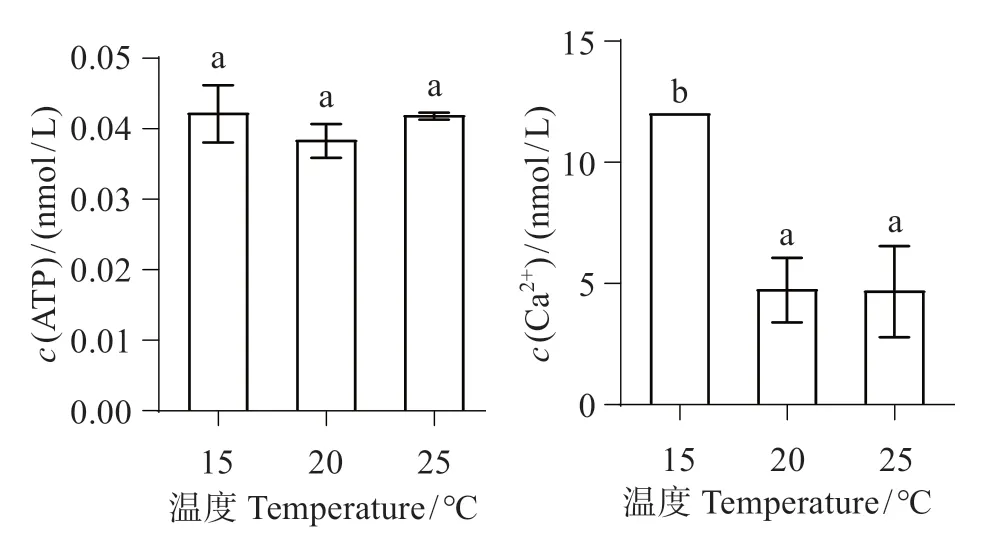
图4 不同温度下中间球海胆肠组织线粒体ATP、Ca2+浓度Fig.4 Mitochondrial ATP,Ca2+concentration of intermediate Strongylocentrotus intermedius tissue at different temperatures
随温度升高,Ca2+-ATP 酶和Ca2+-Mg2+-ATP 酶活性降低,Na+-K+-ATP 酶活性升高。20 ℃组Ca2+-ATP酶活性与对照组相比差异不显著(P>0.05),20 ℃组Ca2+-Mg2+-ATP酶、Na+-K+-ATP酶活性与对照组相比差异显著(P<0.05);25 ℃组Ca2+-ATP 酶、Ca2+-Mg2+-ATP 酶活性与对照组和20 ℃组相比差异极显著(P<0.01)。25 ℃组Na+-K+-ATP 酶活性与20 ℃组相比差异不显著(P>0.05),与对照组相比差异极显著(P<0.01)(图5)。
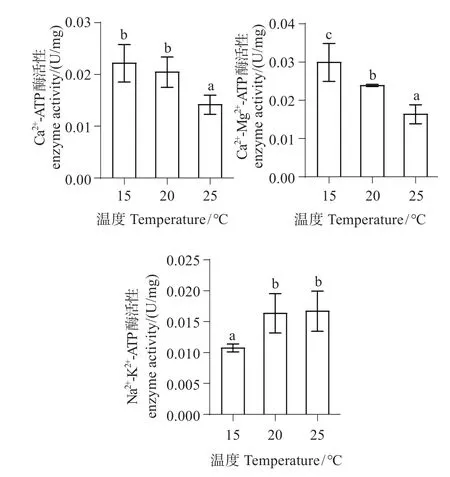
图5 不同温度下中间球海胆肠组织线粒体Ca2+-ATP、Ca2+-Mg2+-ATP和Na+-K+-ATP酶活性Fig.5 The mitochondrial Ca2+-ATP,Ca2+-Mg2+-ATP and Na+-K+-ATP enzyme activity of Strongylocentrotus intermedius tissue at different temperatures
2.3 急性高温胁迫下中间球海胆体腔液抗氧化酶活性变化
图6显示,随温度升高,SOD活性显著升高(P<0.05),25 ℃组较20 ℃组呈下降趋势,变化不明显(P>0.05);CAT活性下降明显(P<0.05),20 ℃组和25 ℃组之间CAT 活性变化不明显(P>0.05);GSHPx活性显著下降(P<0.01),但25 ℃组和20 ℃组之间酶活性变化不明显(P>0.05);POD活性显著升高(P<0.05),20 ℃组和25 ℃组相比差异显著(P<0.05)。
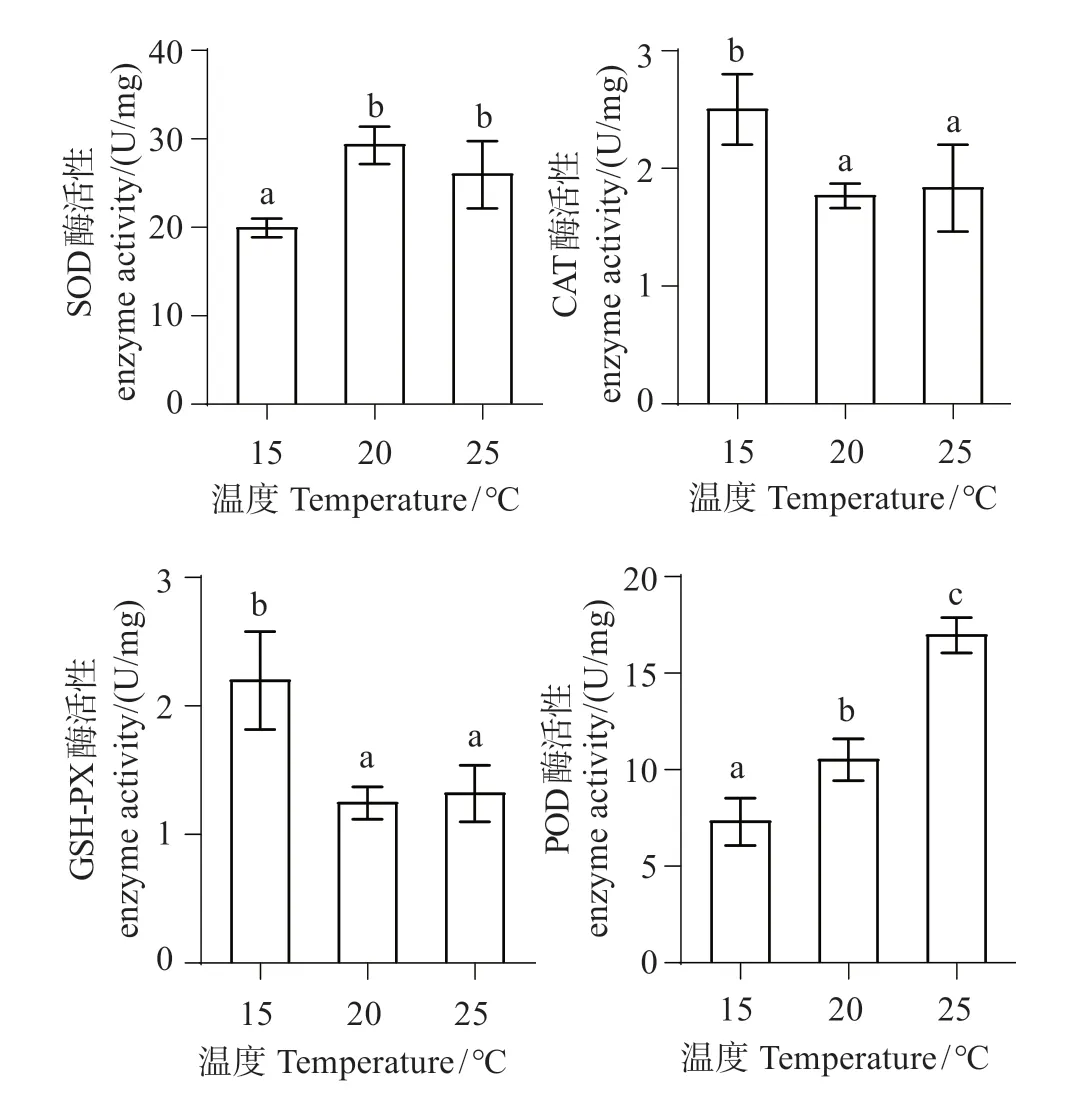
图6 不同温度下中间球海胆体腔液抗氧化酶活性变化Fig.6 Changes in the activity of antioxidant enzymes in Strongylocentrotus intermedius cavity fluid at different temperatures
3 讨论
3.1 急性高温胁迫下中间球海胆体腔细胞线粒体结构变化
高温对中间球海胆体腔细胞结构产生一定影响。本研究结果显示,随温度升高,体腔细胞膜、线粒体数量、线粒体膜及嵴变化趋势明显。盛连喜等[16]对鲤鱼进行胁迫,发现其肝细胞线粒体腔室内有聚集状态的非胶质物质出现。刘晓晖等[6]对鲤鱼进行急性胁迫,发现鲤鱼肝细胞中线粒体数量先增多后减少,第一次热胁迫时部分嵴断裂脱落在线粒体基质中;随温度继续升高,线粒体受损严重,膜结构破坏,嵴脱落等。本实验与其研究结果一致的是,当升温至20 ℃时,体腔细胞内线粒体数目显著增多,推测这与温度突然升高、中间球海胆运动幅度加大、线粒体需提供更多能量有关[17]。当温度升高至25 ℃,线粒体数量减少,这可能是由于高温损害了线粒体的结构和酶的附着位点。本研究结果与鲤鱼急性高温胁迫后线粒体结构不一致的是,随温度升高,鲤鱼肝细胞逐渐出现均质化,线粒体出现空泡化,而中间球海胆的体腔细胞和线粒体逐渐出现浑浊;而在热胁迫小鼠早期囊胚细胞,发现其线粒体数目减少,线粒体发生肿胀、空泡化,线粒体基质变浅呈透明形态、嵴变短变少甚至消失,有的线粒体嵴排列紊乱不清晰[18]。这是脊椎动物与无脊椎动物在高温胁迫下其细胞超微结构的异同点。
3.2 急性高温胁迫下中间球海胆肠组织线粒体特征酶分析
ATP 是重要能量分子,在细胞生理和病理过程中起着重要作用[19]。钱令嘉等[20]对小鼠的热胁迫实验及朱孟凯等[21]对凡纳滨对虾的高温胁迫实验发现线粒体中ATP 浓度显著增高。而本研究结果显示,中间球海胆肠组织线粒体中ATP 浓度无显著变化,推测在高温下中间球海胆消耗更多能量以保持机体内正常平衡[22]。
Ca2+是调节细胞器的重要因子,当细胞内Ca2+浓度发生变化,这是导致细胞死亡的致命一击[23]。随温度升高,Ca2+-ATP酶活性呈下降趋势,推测是由于高温使线粒体氧化代谢能力下降,膜脂质过氧化所导致的膜酶蛋白构象的改变,这降低了细胞内部Ca2+的线粒体的重摄取能力,导致细胞中钙超载。线粒体Ca2+浓度降低是由于线粒体膜上的Ca2+-ATP酶可分解ATP产能,转运Ca2+储存于线粒体内,Ca2+-ATP 酶活性降低,所以Ca2+浓度降低。本实验结果与熊大林等[8]对凡纳滨对虾进行热胁迫,发现Ca2+-ATP酶活性呈下降趋势的结果一致。
Na+-K+-ATP 酶和Ca2+-Mg2+-ATP 酶在维持细胞正常生理功能方面起重要作用。汤平涛等[24]对雄性大鼠进行热胁迫120 min后,发现大鼠肝线粒体中的Ca2+-Mg2+-ATP 酶活性下降,本研究结果与其研究结果一致,可能由于线粒体膜通透性升高,不能将Ca2+通过逆浓度梯度及时运回,从而出现热应激反应[25]。随温度升高,线粒体中Na+-K+-ATP酶活性明显升高,推测温度能够影响其生物膜结构、膜上Na+-K+-ATP酶的构象以及酶对反应离子和底物的亲和力[26]。
3.3 急性高温胁迫下中间球海胆体腔液抗氧化酶活性变化特征
本研究结果表明,中间球海胆随着温度升高,其体腔液中的抗氧化酶活性均发生变化。正常生理状态下,水生动物的抗氧化系统会及时从机体中去除代谢过程中产生的活性氧,而在环境胁迫下,机体内会有大量活性氧产生,从而造成机体损伤[27]。
SOD可将机体产生的毒性O2-转化为H2O2,后者继续被CAT 代谢为无毒的H2O[28-29]。丁文君等[15]对中间球海胆以每天升温1 ℃进行急性胁迫,结果显示SOD 活性均呈先升高后降低趋势。本研究结果显示,中间球海胆随温度升高,SOD 活性显著升高,推测随温度升高中间球海胆机体产生应激反应,导致细胞和组织中自由基迅速增加,所以机体需更多SOD 来应对过多的自由基,从而提高了SOD 活性[30]。当温度升高至25 ℃时,SOD 活性呈下降趋势,但是变化不显著,推测是由于机体中自由基含量增多,超过了SOD清除能力,导致SOD活性下降。
有研究高温对刺参(Apostichopus japonicus)的影响,结果显示CAT 活性随温度升高而降低[31-32]。谢明媚等[9]对银鲳幼鱼急性高温胁迫,其血清中的GSH-Px 活性呈降低趋势。本研究结果显示,随温度升高CAT 及GSH-Px 活性呈下降,表明高温对中间球海胆已造成损伤,抗氧化能力减弱,体内活性氧堆积超过了CAT 和GSH-Px 的清除能力,所以体内过多有毒自由基对CAT和GSH-Px的活性产生了抑制作用[33]。
POD 与生物应激有关,能将过氧化氢水解,可以对细胞起保护作用。本实验结果显示,中间球海胆随热胁迫温度升高,体腔液中POD 活性显著上升,本实验结果与丁文君等[15]、丁小丰等[34]、Wang等[32]研究结果一致,即随水温升高,海胆机体因高温产生应激反应,POD活性升高以保护细胞免受损伤。
4 结论
随温度升高,中间球海胆体腔细胞线粒体结构、肠组织线粒体的ATP 浓度、Ca2+浓度、ATP 酶活性以及抗氧化酶均发生一定变化。高温使中间球海胆的线粒体发生损伤,其中线粒体膜破损,线粒体基质密度显著增大,嵴溶解;Ca2+浓度、Ca2+-ATP酶、Ca2+-Mg2+-ATP 酶活性显著下降,Na+-K+-ATP 酶活性显著升高,与高温胁迫陆生脊椎动物相比不同的是,热胁迫下中间球海胆ATP 浓度没有显著性变化。此外,中间球海胆热胁迫后体腔液SOD 酶、POD 酶活性显著升高,CAT、GSH-Px 酶活性显著下降。综上所述,在生理水平上,中间球海胆通过调节线粒体结构与功能及抗氧化酶活性以响应热胁迫。
本实验有助于从生理水平上阐明中间球海胆如何响应高温胁迫,为以后筛选耐高温中间球海胆提供了一定参考,也为后续研究海胆响应高温胁迫的分子机制提供依据。
猜你喜欢
医学研究生学报(2022年5期)2022-12-07
临床肺科杂志(2022年3期)2022-11-26
小猕猴学习画刊(2022年10期)2022-11-01
中华实用诊断与治疗杂志(2022年1期)2022-08-31
水产科学(2022年3期)2022-06-08
渔业科学进展(2021年6期)2022-01-04
中国卒中杂志(2021年7期)2021-11-29
幼儿画刊(2021年9期)2021-09-20
北方人(2016年9期)2016-05-14
少林与太极(2015年10期)2016-03-09
