
珍珠龙胆石斑鱼豆粕诱导型肠炎肠道潜在生物标志物筛选
2022-07-14 01:23谭北平庞奥博邓君明杨奇慧张海涛
广东海洋大学学报 2022年4期
张 卫,谭北平,庞奥博,邓君明,杨奇慧,张海涛
(1.广东海洋大学水产学院,广东 湛江 524088;2.农业部南方水产与畜禽饲料重点实验室,广东 湛江 524074)
鱼粉(FM)价格上涨迫使寻找新的替代物,大豆制品产量高且氨基酸组成较平衡,是目前应用最广的植物型替代蛋白源。但饲料中添加高剂量豆粕(SBM)易引起鱼类肠炎反应[1],这与其含有大豆抗原蛋白、蛋白酶抑制剂、植酸盐、皂苷、凝集素、植物甾醇和低聚糖等抗营养因子有关[2]。目前,关于SBM 诱导的鱼类肠炎(SBMIE)确切原因和机制尚不完全清楚,对于肠炎形成的分子机制仍缺乏系统研究。醇溶性抗营养因子,尤其是大豆皂苷极可能是主要的潜在致病因素[3]。但大豆皂苷直接作用复杂。单独喂食或与玉米面筋、向日葵、菜籽或蚕豆蛋白联用时,未见或几乎无炎症,然而大豆皂苷与豌豆蛋白合用时,组织学和转录水平发生了显著变化[4]。皂苷引起的肠炎有剂量依赖性,较高含量的皂苷(2~4 g/kg)可引起大西洋鲑(Salmo salar)肠炎,与基础饲料蛋白源种类无关[3]。此外,低水平皂苷似对一些鱼类生长还有促进作用,可能与皂素佐剂特性的免疫促进作用有关[5]。豆粕引发的肠炎也可能是大豆皂苷膜干扰特性的次要作用[6]。新的组学技术对研究鱼类营养与免疫间复杂关系有巨大潜力[7]。
珍珠龙胆石斑鱼(Epinephelus fuscoguttatus♀×Epinephelus lanceolatus♂)是典型的海水肉食性鱼类,有生长快、市场价值高、抗病性强等优点,在养殖过程中也发现SBM 引起的肠道黏膜受损及肠炎症状[1]。本研究对珍珠龙胆石斑鱼肠道组织进行非靶向代谢组学分析,以筛选与饲料SBM 模式相关的差异代谢物,从综合性视角在SBM 饲料替代鱼粉模式下,筛选SBM 诱导的肠炎候选生物标志物,为鱼类肠道健康研究提供参考。
1 材料与方法
1.1 实验饲料
实验饲料组成见表1。分别配制3 种等氮(粗蛋白质量分数约50%)和等脂(粗脂肪质量分数约10%)的实验饲料(20 kJ/g),以相应的SBM 蛋白替代质量分数0%、20%和40%的FM 蛋白,分别命名为FM(对照组)、SBM20 和SBM40。在实验饲料中加入赖氨酸和蛋氨酸进行氨基酸配平。饲料原料磨成细粉,过250 μm 筛,并根据配方准确称量。用造粒机制备粒径为2.0、3.0 mm 的颗粒饲料,阴凉处风干,分装,在-20℃储存备用。饲料中必需氨基酸含量(四川威尔检测技术股份有限公司检测)见表2。
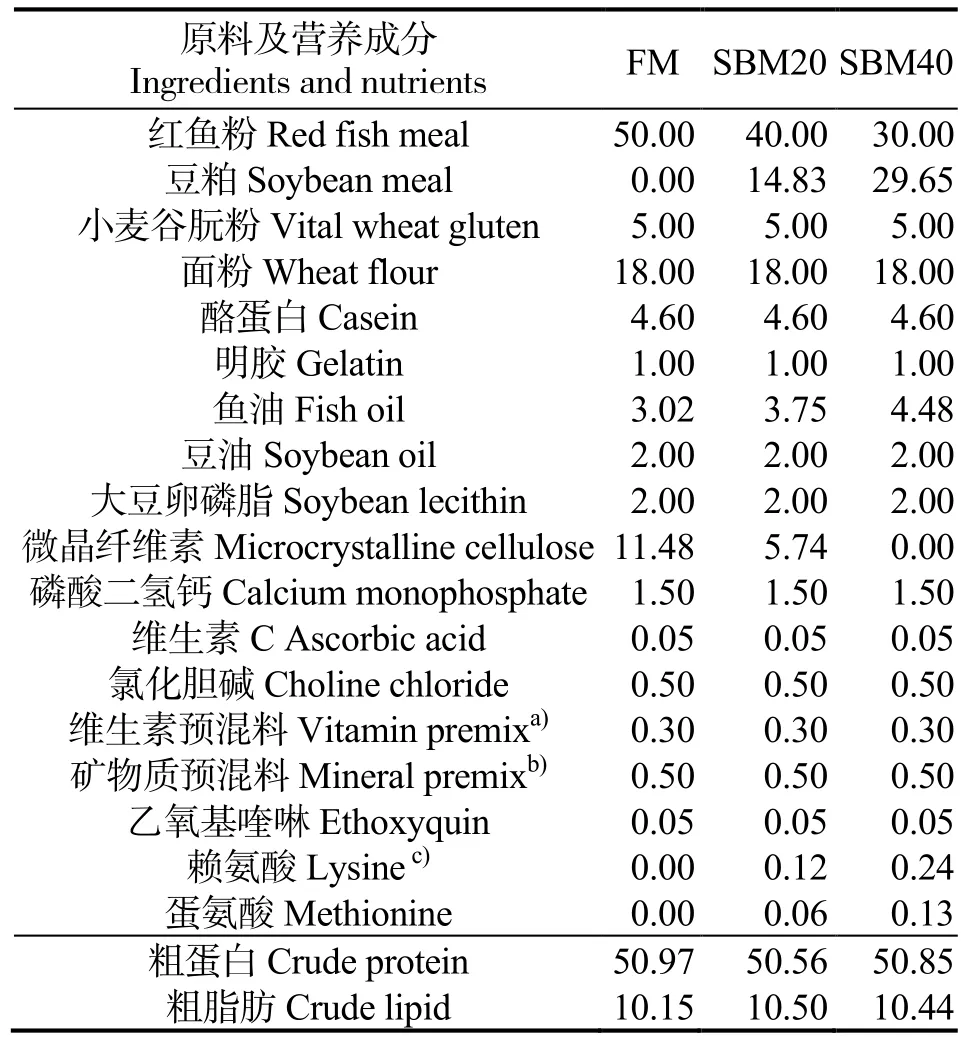
表1 饲料配方和基本组成质量分数 (干物质)Table 1 Formulation and proximate composition of experimental diets (dry matter) %
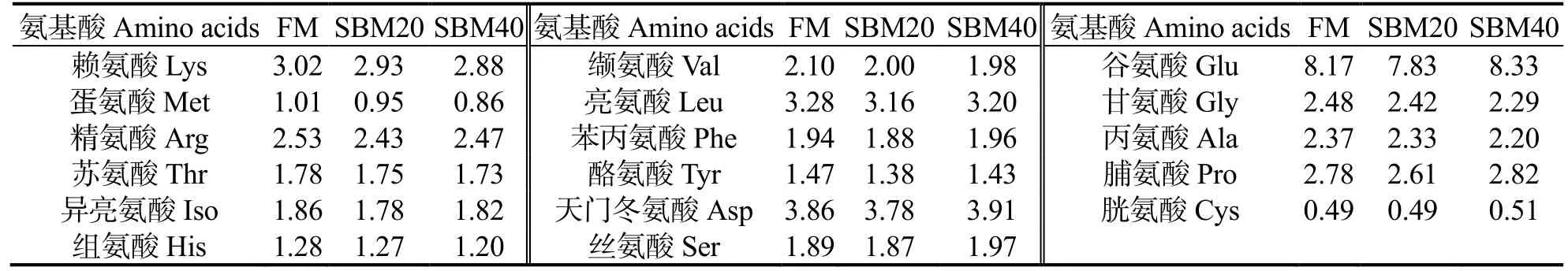
表2 饲料中17 种必需氨基酸质量分数Table 2 Mass fraction of 17 amino acids in diets %
1.2 实验过程
平均体质量约9 g 的健康石斑鱼幼鱼购自湛江市。在实验条件驯养1 周,喂食商品饲料。实验前停饲24 h,用丁香酚(体积分数0.01%)麻醉,分组。规格相似幼鱼随机分配至1 000 L 圆柱形玻璃钢桶中,每桶60 尾,每组4 个重复。每日8:00、16:00投喂至表观饱腹水平,实验周期10 周。实验过程中记录饲料消耗。室内静水养殖,所有养殖桶用气石连续充气,自然光照,温度(29 ± 1)℃,氨和硝酸盐低于0.03 mg·L-1,溶解氧不少于7 mg·L-1。前2周每天每桶换水量为60%;后期每桶每天换水量约100%,为减少鱼的应激,用保证鱼自由游动的最低水位流水式换水,待水足够清澈时加至原有水位。
1.3 样品收集
养殖实验结束时,停饲24 h,统计石斑鱼数量和体质量。每桶随机取鱼6 尾,取血清,置-80 ℃,以分析血清酶活力;取后肠和肝脏,清除肠系膜和脂肪组织,置液氮中,取样后移至-80 ℃以备酶活性分析;部分肠道组织剪碎,置于装有RNAlater的试管,置4 ℃下12 h,移至-80 ℃用于基因表达测定。
在最后摄食时间点前对实验鱼肠道组织进行代谢组学取样。剖取肠道,轻轻挤出粪便,用去离子水冲洗干净。每处理组取12 个样本(同一桶3尾鱼肠道组织等量为一个样本)。样品置冻存管,液氮速冻后,移至 -80 ℃以备代谢组学分析。
1.4 生长生理指标
1.4.1 生长性能 选用增重率(Weight gain rate,WGR,%)、特定生长率(Specific growth rate,SGR,%/d)、肝体指数(Hepatosomatic index,HIS,%)和存活率(Survival rate,SR,%)作为生长性能指标,计算公式:

式中,m0,初始体质量;mt,终末体质量,t试验时间,d。
1.4.2 肠道免疫基因表达 用 Trizol 试剂盒(Invitrogen,Carlsbad,CA,USA)参照说明书提取肠道组织总RNA。取质量合格的RNA 样品,用Evo M-MLV 反转录试剂盒(Takara,Japan)制备cDNA,于-20 ℃下保存待用。用Primer Premier 5.0 设计il1β、il12、tnfα促炎相关基因和il10、igm、cd4抗炎相关基因的引物(表3)[8],内参为β-actin。通过实时荧光定量PCR 检测基因表达量。所用引物模板均来自广东海洋大学水产动物营养与饲料实验室珍珠龙胆石斑鱼2 代和3 代全长转录组测序结果,原始数据的NCBI 登录号分别是PRJNA664623、PRJNA66441。
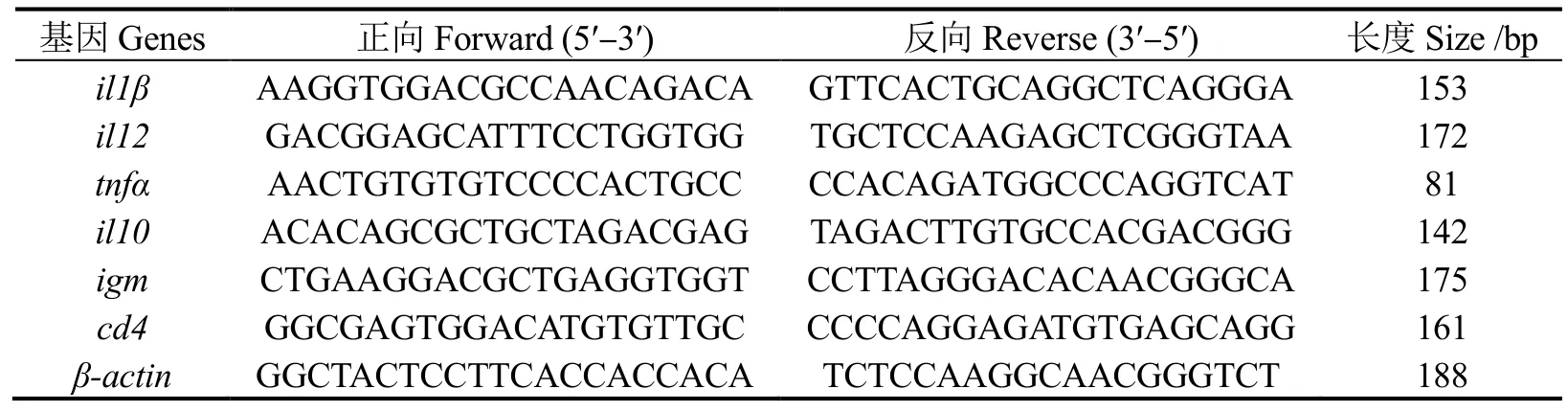
表3 珍珠龙胆石斑鱼肠道免疫相关基因PCR 引物Table 3 PCR primers for intestinal immune related genes of grouper
1.4.3 组织切片及炎症指标分析 每缸随机取鱼3尾,剖取适宜长度后肠,置于体积分数4%多聚甲醛通用组织固定剂(武汉塞维尔生物技术有限公司,武汉,中国)中24 h,制作H&E 切片。根据半定量方法,对H&E 染色切片进行光学显微镜随机盲评,后肠SBM 诱导型肠炎程度评价方法见文献[9]。
1.5 代谢组学分析
1.5.1 代谢物提取及UPLC-MS 分析 肠道样品经液氮充分研磨,用预冷的体积分数80%甲醇和体积分数0.1%甲酸再悬浮。冰上孵育5 min,于15 000 r/min、4 ℃条件下离心5 min。部分上清液用LC-MS级水稀释至甲醇体积分数60%。将样品移至有0.22 μm 过滤器的新鲜Eppendorf 管,于15 000g、4 ℃条件下离心10 min。将滤液注入LC-MS/MS 系统。
1.5.2 质量控制 为控制样本质量,在样品处理的同时制备质量控制(QC)样品。QC 样品是实验样品的等份混合样品,用于平衡色谱-质谱系统,监测系统性能,并在整个实验过程中评价系统稳定性。QC 样本相关性越高(越接近1),整个方法稳定性越佳。在PCA 分析图中反映的是QC 样品的分布聚集在一起[10]。同时建立空白样品去除背景离子。
1.5.3 数据处理和代谢物鉴定 用Compound Discoverer 3.0(CD 3.0,Thermo Fisher)处理UPLC-MS/MS的原始数据,对每种代谢物进行峰对齐、峰提取和定量。主要参数:保留时间耐受性为0.1 min,实际质量公差为5×10-6,信号强度公差为30%,信噪比为3,最低强度为100 000。将峰值强度归一化为总光谱强度。利用归一化数据根据添加剂离子、分子离子峰和碎片离子预测分子式。将峰值与数据库进行比对,以获得准确的定性和相对定量结果。将所选数据导入EZinfo 软件用于主成分分析(PCA)和偏最小二乘判别分析(PLS-DA)。统计分析前对数据进行Pareto 标度处理,用PCA 分析评价数据集的可靠性(包括QC 样本),并提供所有样本可视化总体分离结果。为更好区分不同群体,建立PLS-DA模型。此外,为避免PLS-DA 模型过度拟合,用SIMCA-P+14.0 软件(Umetrics AB,Umea,Sweden)对200 个随机排列进行7 次交叉循环检验。用PLS-DA 模型变量在投影中的重要性(VIP)值和t检验的P值寻找差异代谢产物。VIP 值反映每个变量对模型的贡献。VIP 值较大则可区分对照组和实验组主要潜在生物标志物。VIP >1,P<0.05,对数值差异倍数(Fold change,FC)≥2 或FC≤0.5 的代谢产物视为差异性代谢物[11]。
1.5.4 差异代谢物分析 用火山图筛选代谢物的log2(FC) 和-lgP,用z-Score 图展示同一水平代谢物的相对含量,用cor.mtest() 函数计算不同代谢物间协同或互斥关系,显著相关的阈值水平为P<0.05。
1.6 统计分析
统计分析用SPSS 22.0 进行,结果以平均±标准差表示。为检验各组间的差异,同质方差检验后进行单因素方法分析。显著性阈值为P<0.05。对代谢物进行受试者工作特征曲线(ROC)分析以确定曲线下面积(AUC),比较代谢物预测能力。
2 结果
2.1 生长性能
与对照组相比,随着SBM 替代水平的增加,各实验组增重率和特定生长率依次显著降低(P<0.05);各组肝体比和存活率无显著变化(P>0.05)(表4)。

表4 不同水平豆粕蛋白替代鱼粉蛋白对珍珠龙胆石斑鱼生长的影响Table 4 Effects of different levels of soybean meal protein substitute for fish meal protein on growth of pearl gentian grouper
2.2 免疫相关基因表达
表5 显示,与对照组相比,随着替代水平的增加,各实验组促炎相关基因il1β、il12、tnfα表达量总体上呈显著增加趋势(P<0.05),SBM40 组表达量最高;随着替代水平的增加,与对照组相比,抗炎相关基因il10、igm、cd4表达量呈显著下降趋势(P<0.05),SBM40 组表达量最低。

表5 豆粕替代鱼粉蛋白对珍珠龙胆石斑鱼后肠免疫相关基因表达的影响Table 5 Effects of soybean meal protein substitute for fish meal protein on immune related gene expression in hindgut of pearl gentian grouper
2.3 组织切片和炎性指标
组织切片和半定量分析表明,与对照组相比,不同替代水平的SBM 均引起珍珠龙胆石斑鱼后肠炎症反应,有显著差异(P<0.05)。其中SBM40组炎症最严重,SBM20 组次之,有较明显的炎症表征,如肠道黏膜皱襞缩短、固有膜肿胀及各种炎性细胞浸润等(图1 和表6),说明本研究SBM 替代水平诱发了珍珠龙胆石斑鱼的肠炎症状。
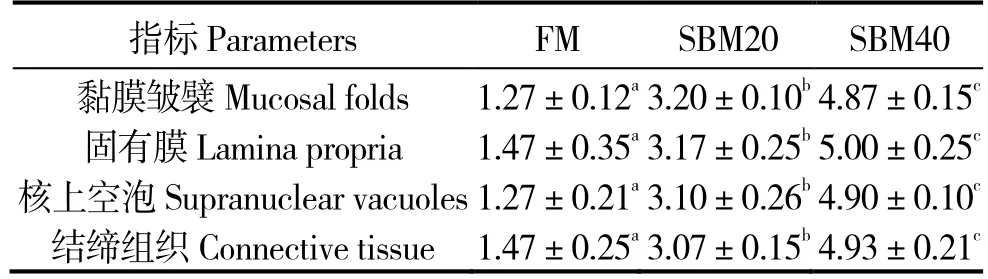
表6 肠道切片半定量组织学肠炎评估得分Table 6 Semi-quantitative histological evaluation of intestinal sections
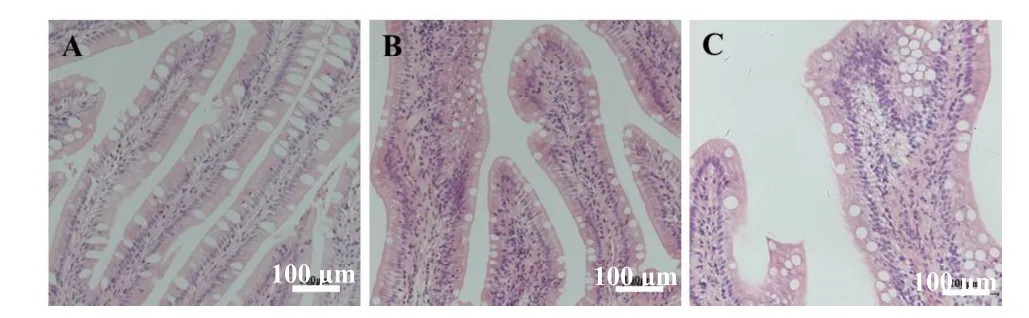
图1 珍珠龙胆石斑鱼后肠Fig.1 Hindgut of pearl gentian grouper
2.4 肠道组织UPLC-MS 代谢谱分析
2.4.1 UPLC-MS 代谢谱分析 肠道组织的代谢谱图如图2。图3 展示LC-MS 正负离子两种模式下肠道组织样本的PCA 得分。QC 样本位于中心,聚类紧密,表明数据质量好,模型稳定可靠。图4 展示QC 样本的相关性。
图2 正、负离子模式下后肠组织的典型UPLC-MS 光谱Fig.2 Representative UPLC-MS spectra of the distal intestine tissues in positive and negative modes
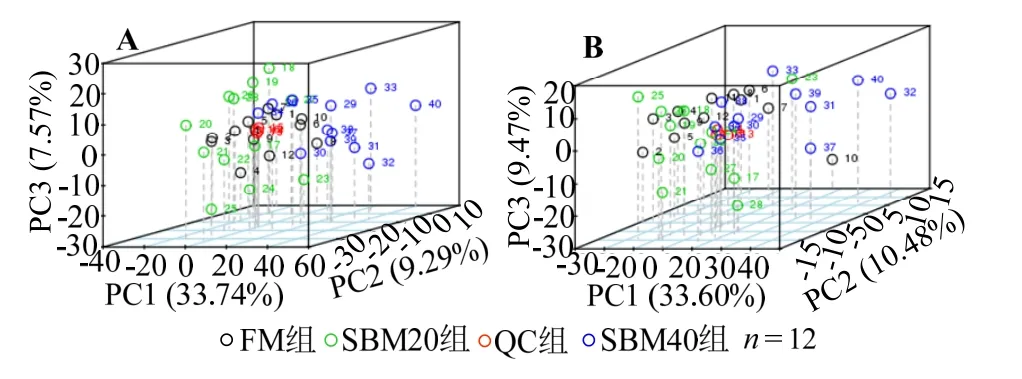
图3 正、负离子模式下后肠总样本主成分分析(PCA)Fig.3 Total sample PCA score plots resulting from UPLC-MS spectra of distal intenstine in positive and negative modes

图4 正离子和负离子模式下后肠组织质控样品相关性Fig.4 Correlation of QC samples of distal intestine tissues in positive and negative modes
图5 展示正、负离子模式下肠道组织差异代谢物PLS-DA 模型的结果。两组间分离清晰,各组样本基本在95%置信椭圆内。用正、负两种离子模式进行置换检验,以防止模型过度拟合 (图6)。FM 和SBM20 组的置换检验R2Y 和Q2Y 参数在正离子模式中为分别0.73和0.67,在负离子模式中分别为0.61和0.73;FM 和SBM40 组的置换检验R2Y 和Q2Y参数在正离子模式中为分别0.67 和0.62,在负离子模式中分别为0.57 和0.65。其值均大于0.5,左边的红线(Q2Y)和蓝线(R2Y)比右边起始点低,表明这些模型过拟合风险低。
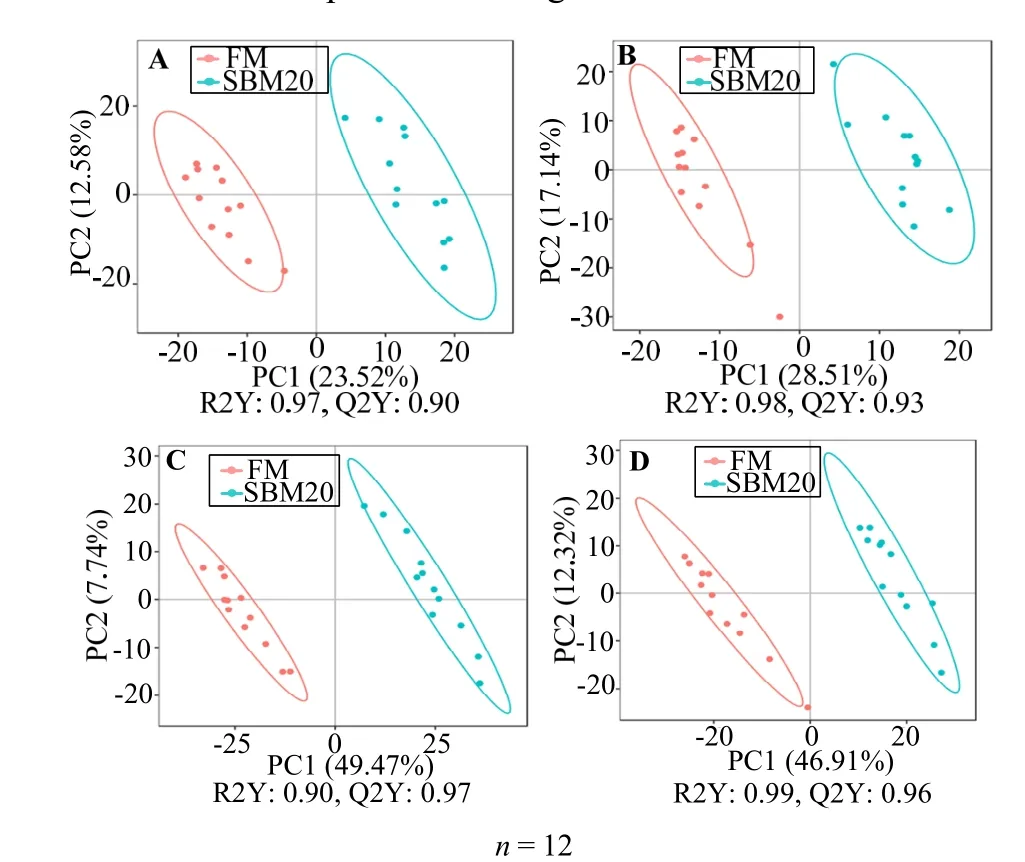
图5 正离子(A,C)和负离子(B,D)模式下后肠组织总样本PLS-DA 评分Fig.5 Total sample PLS-DA score resulting from the UPLC-MS spectra of distal intestine tissues in positive (A,C) and negative modes (B,D)
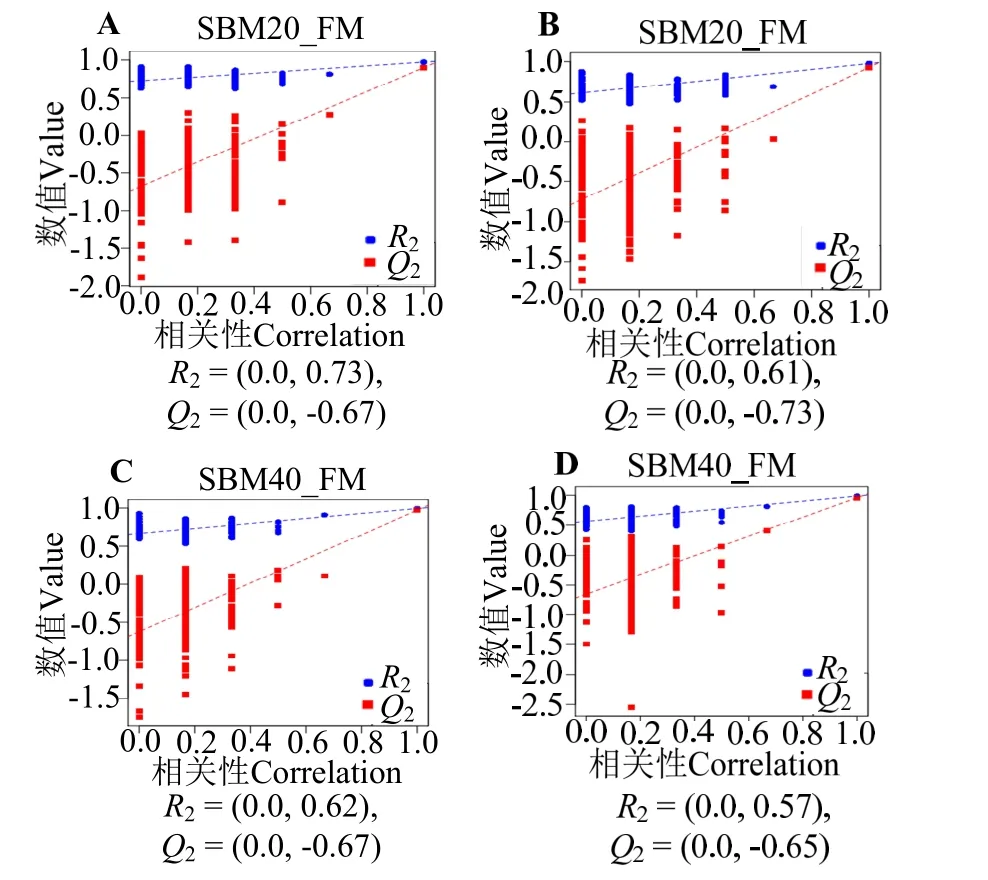
图6 正离子(A,C)和负离子(B,D)模式下后肠组织PLS-DA模型置换检验Fig.6 Permutation test result of the PLS-DA models of distal intestine tissues in the positive (A,C) and negative (B,D) modes
2.4.2 差异代谢物分析 差异代谢物的PLS-DA 得分图(图5)显示,FM 组与SBM20 组和SBM40 组之间可明显分离,且未过拟合。与FM 对照组相比,正离子和负离子模式下,SBM20 组和SBM40 组中,从肠道组织中各鉴定2 260 种差异代谢物(正离子模式1 404 种,负离子模式856 种)。所鉴定物质的保留时间、质/荷比(m/z)和碎片模式与标准物质比较,正离子模式下,SBM20 组差异代谢物中有53 种代谢物显著上调,120 种代谢物显著下调(图7);SBM40 组差异代谢物中有69 种代谢物显著上调,293 种代谢物显著下调。同时,负离子模式下,SBM20组差异代谢物中有52 种代谢物显著上调,77 种代谢物显著下调;SBM40 组差异代谢物中有46 种代谢物显著上调,190 种代谢物显著下调。
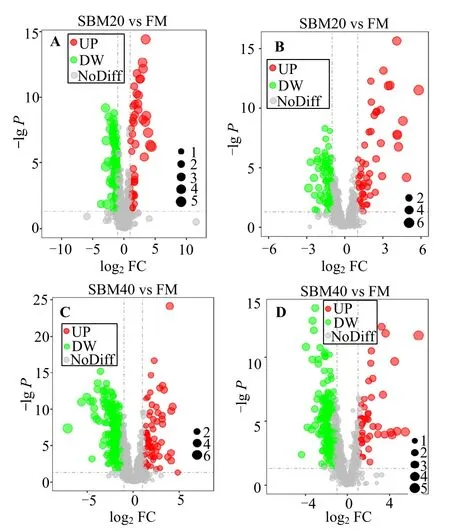
图7 正离子(A,C)、负(B,D)离子模式下组间P 值火山图Fig.7 Volcano plot of P values between FM,SBM20 and SBM40 groups in positive (A,C) and negative (B,D) modes
2.4.3 潜在生物标志物筛选 根据VIP >1,|log2(FC)| >1,P<0.05,在正离子模式下,在SBM20 组中,筛选的前10 种可区分FM 和SBM20 组最有影响差异代谢物分别为染料木素、大豆黄酮、染料木素-4'-O葡萄糖醛酸酯、癸二酸、鸡黄豆素、2-O-乙基抗坏血酸、柠檬苦素、吲哚-3-丙烯酰甘氨酸、2-苯基-1,4-二羟基萘和白三烯B4 二甲胺;在SBM40 组中,筛选的前10 种可区分FM 和SBM40 组最有影响差异代谢物分别为2-氨基-3,4-二甲基咪唑(4,5-F)喹啉、柠檬苦素、吲哚-3-丙烯酰甘氨酸、N-甲基蒽酰胺、N,N-二甲基甲氧基噻吩、染料木素、染料木素-4’-O 葡萄糖醛酸酯、白三烯B4 二甲胺、大豆黄酮和5-羟基吲哚-3-乙酸。
在负离子模式下,在SBM20 组中,筛选的前10种可区分FM和SBM20组最有影响差异代谢物分别为染料木素、维生素C、鸡黄豆素、黄芩苷、大豆黄酮、葡萄糖二酸、大豆皂苷Ⅰ、苜蓿酸3-O-三葡萄糖苷、小肽(Leu-Gly-Pro)和戊基-4-羟脯氨酸;在SBM40组中,筛选的前10 种可区分FM 和SBM40组的最有影响差异代谢物分别为维生素C、染料木素、葡萄糖二酸、大豆黄酮、黄芩苷、小肽、戊基-4-羟脯氨酸、大豆皂苷Ⅰ、鸡黄豆素和磷酸肌酸。SBM20 组替代水平下,正负离子模式下的20 种潜在肠炎标志物中,有3 种是共有的;SBM40 组替代水平下,正负离子模式下的20 种潜在肠炎标志物中,有2 种是共有的。因此,SBM20 和SBM40 组分别选出17 种和18 种代谢物作为珍珠龙胆石斑鱼肠道组织豆粕诱导型肠炎(SBMIE)的潜在生物标志物。
为进一步明确潜在生物标志物特征,图8 展示差异代谢物正负离子模式下的z-得分(z-score)。表7 展示正、负离子模式下,SBM20 组潜在肠炎标志物的离子强度;表8 展示正、负离子模式下,SBM40组潜在肠炎标志物的离子强度。与FM 组对比,SBM20 组癸二酸、柠檬苦素、吲哚-3-丙烯酰甘氨酸、小肽Leu-Gly-Pro、戊基-4-羟脯氨酸和磷酸肌酸离子强度显著降低(P<0.05),而其他12 种代谢物的强度显著增加(P<0.05);SBM40 组2-氨基-3,4-二甲基咪唑(4,5-F)喹啉、柠檬苦素、吲哚-3-丙烯酰甘氨酸、N-甲基蒽酰胺、N,N-二甲基甲氧基噻吩、小肽Leu-Gly-Pro、戊基-4-羟脯氨酸和磷酸肌酸的离子强度显著下降(P<0.05),其他10 种代谢物强度显间内,代谢物AUC 均超过0.9(图9),表明对潜在生物标志物预测能力较佳。
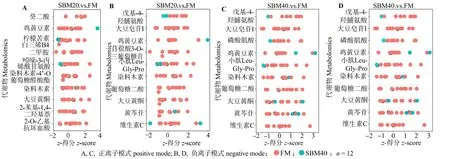
图8 正离子(A,C)和负离子(B,D)模式下FM 与SBM20、SBM40 组间潜在生物标记物的z-Score 评分Fig.8 z-Score plot of potential biomarkers for comparison between FM and SBM20 and SBM40 in positive and negative modes
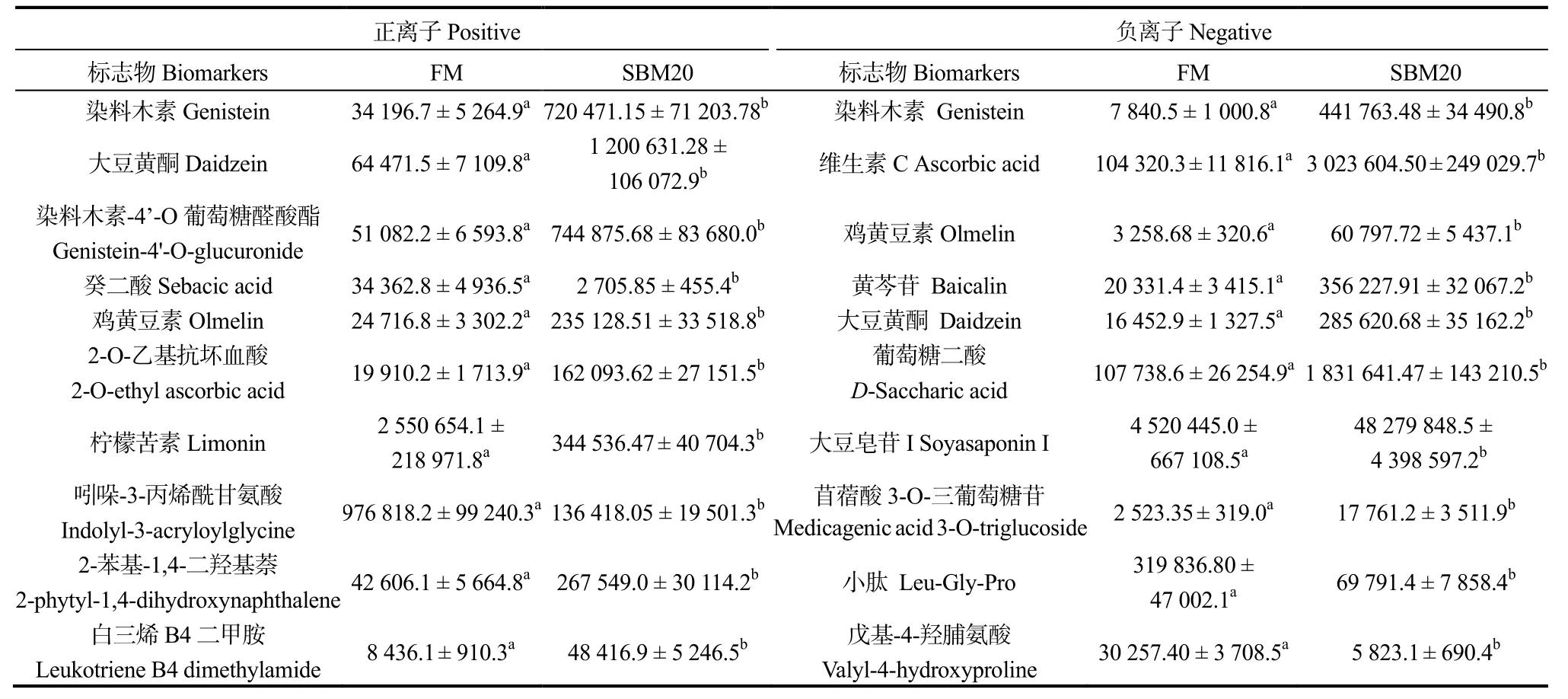
表7 SBM20 组正负离子模式下肠炎潜在标志物的离子强度Table 7 Intensities of metabolites identified as the potential biomarkers of SBM20 vs.FM in positive and negative modes
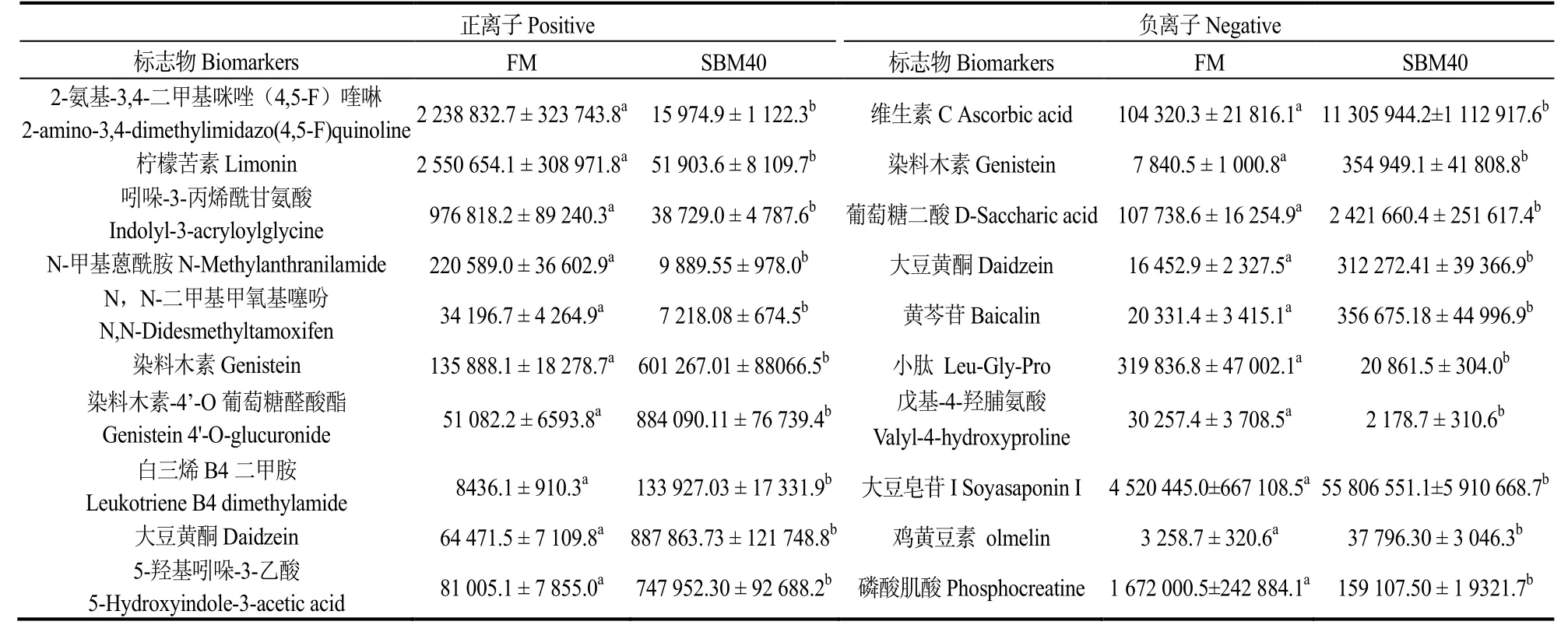
表8 SBM40 组正负离子模式下肠炎潜在标志物的离子强度Table 8 Intensities of metabolites identified as the potential biomarkers of SBM20 vs FM in positive and negative modes
2.4.4 潜在生物标志物筛选 图9 表明,SBM20 组中,正离子模式下,柠檬苦素、吲哚-3-丙烯酰甘氨酸与染料木素-4'-O 葡萄糖醛酸酯、2-O-乙基抗坏血酸、2-苯基-1,4-二羟基萘、白三烯B4 二甲胺呈显著负相关;吲哚-3-丙烯酰甘氨酸与大豆黄酮呈显著负相关,白三烯B4 二甲胺与癸二酸呈显著负相关。其他潜在肠炎生物标志物间呈显著正相关;负离子模式下,小肽(Leu-Gly-Pro)和戊基-4-羟脯氨酸与其他标志物大部分呈显著负相关。正、负离子模式中,其他生物标志物间呈显著正相关(空白处除外)。
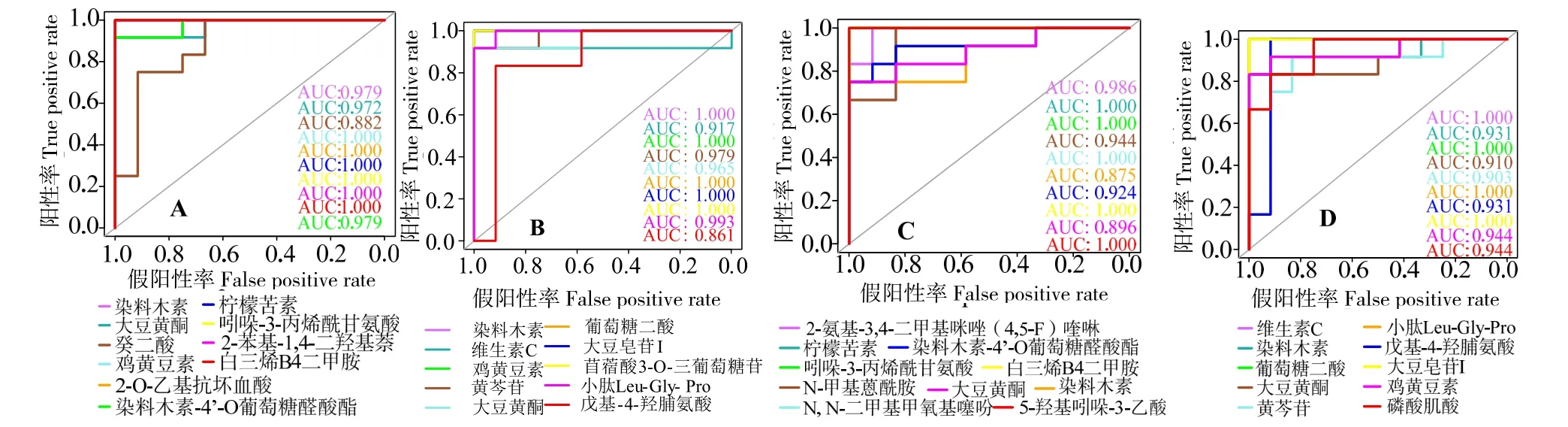
图9 正离子(A,C)和负离子(B,D)模式下组间潜在生物标志物的ROC 分析Fig.9 ROC analysis of potential biomarker metabolites between groups and FM in positive (A,C) and negative (B,D) modes
SBM40 组中,正离子模式下,染料木素-4′-O葡萄糖醛酸酯、白三烯B4 二甲胺、5-羟基吲哚-3-乙酸与2-氨基-3,4-二甲基咪唑(4,5-F)喹啉、柠檬苦素、吲哚-3-丙烯酰甘氨酸、N,N-二甲基甲氧基噻吩呈显著负相关;此外,柠檬苦素还大豆黄酮呈显著负相关。负离子模式下,除戊基-4-羟脯氨酸和磷酸肌酸外,小肽(Leu-Gly-Pro)与其他其中标志物呈显著负相关。戊基-4-羟脯氨酸与维生素C、染料木素、葡萄糖二酸、黄芩苷、大豆皂苷Ⅰ呈显著负相关。磷酸肌酸与维生素C、葡萄糖二酸、大豆皂苷Ⅰ呈显著负相关。正离子和负离子模式中,其他生物标志物间呈显著正相关(空白处除外)。
对比不同SBM 蛋白替代水平肠炎潜在生物标志,发现SBM20 组和SBM40 组有13 种标志物较为保守,称之为核心生物标志物(表9)。除核心生物标志物外,SBM20 组特有的标志物包括癸二酸、2-O-乙基抗坏血酸、2-苯基-1,4-二羟基萘和苜蓿酸3-O-三葡萄糖苷;SBM40 组特有的标志物包括磷酸肌酸、5-羟基吲哚-3-乙酸、N,N-二甲基甲氧基噻吩、N-甲基蒽酰胺、2-氨基-3,4-二甲基咪唑(4,5-F)喹啉。

表9 不同替代水平肠道组织肠炎潜在生物标志物对比Table 9 Comparison of potential biomarkers of enteritis in intestinal tissues at different substitute levels
3 讨论
SBMIE 在水产养殖中普遍存在,对鱼类生长和饲料利用有明显的负面影响,如显著降低养殖对象的增重率、特定生长率,增加饲料系数等。SBMIE为非感染性亚急性肠炎,其组织学特征主要表现为黏膜褶皱缩短,固有膜和黏膜下层肿胀,各种炎性细胞浸润,肠上皮细胞吸收,空泡减少[12]。肠炎主要发生在后肠部分,后肠是依靠胞吞作用吸收蛋白质的主要部位,这种特殊的吸收方式,导致对各种外源抗原的接触更加密切,因此更易诱发食物感染引起的肠道疾病[13]。本研究发现,实验水平的SBM引起了珍珠龙胆石斑鱼肠炎的产生,生长性能和肠道生理结构的变化也证实这一结果。
本研究中,珍珠龙胆石斑鱼后肠组织代谢表型在FM 组和实验组之间表现出显著的模式差异。在PLS-DA 评分图中,FM 组与SBM20、SBM40 组明显分离,表明饲料中SBM 的添加引起了后肠组织代谢谱变化,SBMIE 在代谢水平上有一定差异表征。
VIP 值反映变量的重要性,在代谢组学中常用来筛选潜在的生物标志物[14]。本研究中,在SBM20和SBM40 替代水平下,分别根据VIP 值筛选到17和18 种代谢产物作为珍珠龙胆肠道组织SBMIE 的潜在生物标志物,其中有13 种为保守的核心生物标志物。与FM 组相比,SBM 替代FM 后肠道组织中异黄酮和皂苷等明显增加。
异黄酮在结构上与天然雌激素相似,在动物体内有类似雌激素的生物学效应[15]。SBM 富含植物雌激素(称为异黄酮)。目前,普遍认为大豆异黄酮主要由12 种化合物组成,可分为游离型苷元和结合型糖苷两种类型。游离型苷元主要包括染料木素、大豆苷元和大豆黄素;结合型糖苷主要包括染料木苷、大豆苷、丙二酰染料木苷和丙二酰大豆苷等。本研究的13 种潜在核心生物标志物中,有5 种属于黄酮类化合物,包括染料木素、大豆黄酮、染料木素-4′-O-葡萄糖醛酸酯、鸡黄豆素和黄芩苷。大豆异黄酮也被称为生长促进剂,用于家禽养殖以提高产量。然而,大豆异黄酮对鱼类生长性能、饲料利用和免疫反应的影响是有争议的,可能与其类型、用量及实验对象性别等有关[16-17]。当大豆异黄酮在饲料中添加量(质量分数)达到 0.8%时,牙鲆(Paralichthys olivaceus)后肠完整性受到损害;SBM中大豆异黄酮质量分数为0.10%~0.35%,当大豆异黄酮添加量不超过 0.4% 时,对异育银鲫(Allogynogentic silver crucian carp)和牙鲆的生长和肠道生理几乎无负面影响[18-19]。然而,当SBM 以原料形式整体添加以取代FM 时,大豆异黄酮的负面影响不能被排除,因为饲料中的抗营养因子之间可能存在协同作用,如本研究图10 显示,大豆皂苷与其他抗营养因子之间存在这种现象。
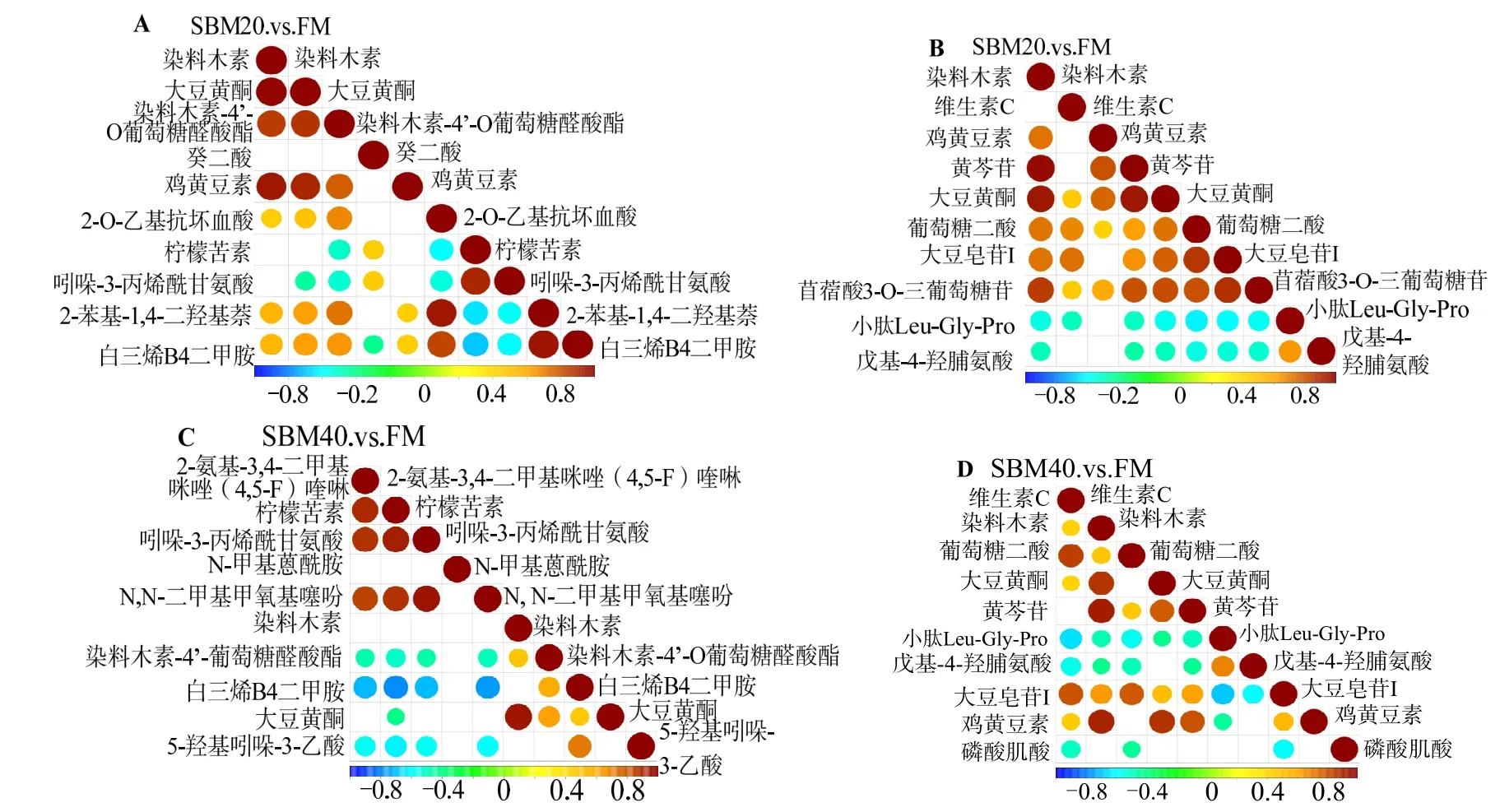
图10 正离子(A,C)和负离子(B,D)模式下组间潜在生物标志物相关性分析Fig.10 Correlation analysis of the potential biomarkers between groups and FM in positive (A,C) and negative (B,D) modes
在水产养殖中,大豆皂苷是一种研究较为广泛的抗营养因子。本研究中,二替代组中大豆皂苷I含量均显著增加。SBM 的皂苷质量分数为5~7 g/kg[20]。在大西洋鲑中,大豆皂苷是引起肠炎的主要抗营养因子[21]。但是,大豆皂苷引起肠炎的直接作用是复杂的,不同抗营养因子间的协同或拮抗作用亦较为重要[3]。Zhang 等[19]发现,虽然用SBM 代替FM 饲喂异育银鲫的负面效应可能与大豆异黄酮无关,但大豆皂苷与大豆异黄酮之间存在协同效应。本研究中,不同潜在生物标志物与肠炎之间的关系有待进一步研究。
13 种潜在核心生物标志物中剩余7 种分别是柠檬苦素、吲哚-3-丙烯酰甘氨酸、白三烯B4 二甲胺、维生素C、葡萄糖二酸、小肽Leu-Gly-Pro 和戊基-4-羟脯氨酸。柠檬苦素是一类三萜类化合物,有抗炎、抑菌、抗氧化、解毒和生物除虫等多种生物活性[22],在治疗溃疡性结肠炎方面有显著的疗效[23]。吲哚类化合物有抗炎、抗癌、抗菌、抗病毒、清除自由基和保护神经等作用[24]。吲哚-3-丙烯酰甘氨酸可能是色氨酸生物合成过程中的代谢中间体,由色氨酸经吲哚丙酸和吲哚乙酸合成。Bull 等[25]研究发现,吲哚-3-丙烯酰甘氨酸可作为诊断自闭症谱系障碍的尿液标记物。白三烯B4 是一种与炎性反应有关的白三烯类物质。受病原感染后,组织释放的白三烯B4 可提高巨噬细胞和中性粒细胞对病原菌的吞噬和杀伤功能,并促进其释放抗菌性活性介导物[26]。但相关研究也指出,过量的白三烯B4 合成与释放,可能会导致各种炎性疾病,如支气管哮喘和关节炎等[27]。小肽有多种生物学功能,如保护肠道结构和功能、促进蛋白质的吸收利用、改善动物的生长性能、提高动物免疫性能及抗氧化作用等。体外实验发现,沙丁鱼肌肉水解小肽(如Val-Tyr、Ile-Tyr、Tyr-Val、Trp-His)可诱导收缩主动脉的松弛,用以预防动脉粥样硬化[28]。也发现Ile-Arg-Trp、Ile-Gln-Trp(蛋),Val-Pro-Pro、Ile-Pro-Pro、Leu-Pro-Pro(发酵豆粕)、Val-Pro-Tyr(豆粕)等食物来源的小肽有抗炎、降压、降糖和改善肾功能等作用[29]。羟脯氨酸在FM 中含量丰富,而在植物原料中含量极少甚至几乎不存在。以往研究指出,羟脯氨酸除可改善鱼类生长性能外,还有清除氧化剂、调控细胞凋亡的作用,且游离的羟脯氨酸可下调炎性基因的表达[30]。本研究发现,SBM 替代FM 后,柠檬苦素、吲哚-3-丙烯酰甘氨酸、白三烯B4 二甲酸、小肽Leu-Gly-Pro 和戊基-4-羟脯氨酸的含量显著降低,提示这些物质的缺乏与肠炎的发生有密切关系。维生素C 是鱼类必需的水溶性维生素,不仅影响生长和饲料利用,还有免疫调节作用,是与水生动物免疫、铁代谢和血液学相关的营养素之一[31]。维生素C 对鱼类免疫有有益影响,如溶菌酶及补体活性、吞噬活性、抗糖化、呼吸爆发和黏膜免疫反应等[32]。葡萄糖二酸是一种无毒的葡萄糖衍生物,通常以手性化合物D-葡萄糖二酸的形式存在。葡萄糖二酸天然存在于樱桃、柑橘和豆类等水果和蔬菜中,少量哺乳动物和人体内也有分泌[33]。葡萄糖二酸在水产养殖中的应用鲜有报道,在哺乳动物中发现,与葡萄糖二酸可相互转化的葡萄糖-1,4-内酯有很强的解毒和抗氧化能力,还可缓解肠道黏膜损伤[34]。尿液中葡萄糖二酸在人类药物性肝炎诊断和鉴别中也视为一种标志物[35]。本研究中,SBM 替代FM 后,维生素C 和葡萄糖二酸的含量显著升高,提示肠炎状态下,鱼类可能通过增加某些物质的含量缓解肠道损伤或炎症。
不同替代水平下,珍珠龙胆肠道组织中筛选的潜在生物标志物中,SBM20 组特有的是癸二酸、2-O-乙基抗坏血酸、2-苯基-1,4-二羟基萘和苜蓿酸3-O-三葡萄糖苷,SBM40 组特有的是磷酸肌酸、5-羟基吲哚-3-乙酸、N,N-二甲基甲氧基噻吩、N-甲基蒽酰胺和2-氨基-3,4-二甲基咪唑(4,5-F)喹啉。通过本研究,可将此类物质作为判定珍珠龙胆石斑鱼SBMIE 程度的标志性物质。但在鱼类SBM 诱导的肠炎研究中,对这些代谢物与肠炎关系鲜有报道。从本研究结果看,虽然这些潜在肠炎标志物之间相关性较高,但在实际生理过程中,其对肠炎的作用是独立的、协同的还是拮抗的,尚不完全清楚,仍有待于进一步研究。
4 结论
基于代谢组学分析,本研究在20%和40%SBM替代水平下,从珍珠龙胆肠道组织中分别筛选到17和18 种SBMIE 的潜在生物标志物;有13 种较为保守的核心生物标志物为两实验组共有,反映了珍珠龙胆石斑鱼SBMIE 的典型表征;两组分别特有的潜在肠炎标志物可作为肠炎严重程度的判定依据。不同替代水平下,大部分生物标志物间存在显著的正相关或负相关作用,但其与肠炎之间的关系和作用机制仍需进一步实验验证。
猜你喜欢
上海建材(2022年3期)2022-11-04
中国典型病例大全(2022年10期)2022-05-10
浙江林业科技(2022年1期)2022-02-20
科学与生活(2021年3期)2021-11-10
表面技术(2021年3期)2021-04-07
今日农业(2021年1期)2021-03-19
中国应急管理科学(2021年9期)2021-03-16
家庭医药·快乐养生(2020年11期)2020-12-03
保健文汇(2020年11期)2020-12-03
人物画报(2020年29期)2020-03-14
