
不同噪声对安徽树蛙行为响应的影响
2025-02-21 00:00:00史素心江清悦王亚伦张保卫方光战
野生动物学报 2025年1期
关键词:背景噪声
关键词:同种广告鸣叫;雌性选择;雄性竞争;背景噪声;安徽树蛙
动物通讯指个体间信息交流的方式[1],对鸟类、两栖类和昆虫等大多数发声动物而言,声音通讯是信息传递和交流的主要方式[2]。动物声音信号通常包含发声者的位置、大小和个体身份等重要信息[2−4],具有保卫领地、促进繁殖等功能;同时,动物还可利用声音定位资源或危险[5−7]。对绝大多数无尾两栖类动物而言,雄性竞争和雌性选择几乎完全依赖声音通讯[3,8−9],但无尾类动物聚集而成的求偶场往往容纳着不同物种及同一物种的诸多雄性个体,从而形成极其嘈杂的声学环境[4]。在嘈杂环境中同种和异种声音形成的背景噪声会掩盖个体声音信号并降低其信噪比,进而给交流和配偶选择带来困难[10−12]。此外,动物发声也会吸引天敌注意,从而使发声者面临着被捕食的巨大风险[13],如雄性泡蟾(Physalaemus pustulo⁃sus)最具性吸引力的鸣声也最能增加被缨唇蝠(Tra⁃chops cirrhosus)捕食的风险[14]。相反,噪声对某些物种而言也可能有益,这取决于环境及接收者对噪声的响应[15],如溪流声可增强雄性小湍蛙(Amolops torren⁃tis)鸣声对雌性的吸引力[16]。另一方面,合作假说(cooperative hypothesis)认为集群鸣声不但可增加群体对雌性的吸引力,同时可干扰捕食者定位特定个体[17],如雌性犬吠蛙(Hyla gratiosa)可被远处同种雄蛙的集群鸣叫吸引[18],在求偶场中鸣叫的雄性泡蟾可降低被捕食风险,并提高交配成功率[19]。据此推测,动物在响应被噪声干扰的同种声音时可能存在“trade-off”,以权衡收益与被捕食风险[20−21],但尚不清楚雌雄两性如何响应被噪声干扰的同种声音信号。
背景噪声包括生物噪声和非生物噪声,前者指同种和异种声音信号,后者包括风声、雨声或流水声[22−23]及人为噪声等[24]。对声音接收者而言,其面临的主要挑战是区分来自不同物种的鸣声信号和背景噪声[25]。对于发声者而言,通常需发出有别于背景噪声的鸣声,以便于接收者有效感知和识别[23]。发声动物可采用不同的发声策略应对噪声干扰,如减少发声活动[26−29],提高鸣叫振幅、鸣叫速率和鸣叫次数[30−32]以及延长发声持续时间[33]等。因而雄性发声策略具有动态性,可根据竞争者的数量或鸣叫率、同种或异种广告鸣叫的存在、雌性的接近以及噪声环境特征等调节自身鸣叫特征,产生更多非重叠鸣叫,提高吸引雌性的概率[2]。此外,对背景噪声的熟悉程度可能影响动物行为,如凹耳臭蛙(Odorranatormota)、牛蛙(Rana catesbeiana)和青铜蛙(R. clami⁃tans)的雄性个体可辨别熟悉的邻近个体声音,对其声音产生较少的行为响应,即“dear enemy”现象[34−36];但目前尚不清楚不同声学特征(包括不同熟悉程度)的背景噪声如何影响雌雄两性的行为响应。
在繁殖季节,雄性安徽树蛙(Zhangixalus zhou⁃kaiyae)一般在田埂底部边缘筑洞并发出由十多个甚至更多音节组成的广告鸣叫[37−38],同时根据环境动态调节竞争策略[38];雌性则通过广告鸣叫识别和评估雄性个体特征,选择最适雄性作为配偶[25]。大别山姬蛙(Microhyla dabieshanensis)与安徽树蛙同域分布,二者繁殖季节重叠[39]。相比于空间分布稀疏的安徽树蛙而言,大别山姬蛙种群密度极大,其鸣声会形成高强度的背景噪声。安徽树蛙雌雄两性可能通过选择性注意等方式,有效减轻噪声对其听觉感知的影响,较为准确地区分同种和异种广告鸣叫。因此,安徽树蛙是研究噪声干扰下雌雄两性行为响应的理想模型。
本研究以安徽树蛙为试验对象,向其回放同种广告鸣叫、同域异种大别山姬蛙广告鸣叫(熟悉)及异域异种宝兴树蛙(Zhangixalus dugritei)广告鸣叫(非熟悉)、白噪声以及同种广告鸣叫与其他3种声音组合而成的声音,记录并分析雌雄两性的行为响应,探究非同种声音对雌性选择和雄性竞争的影响。笔者预测:(1)雌性安徽树蛙更为偏爱信噪比高的同种广告鸣叫;(2)雄性安徽树蛙会根据声音信号,动态调整鸣叫行为;(3)由于安徽树蛙呈稀疏分布,个体鸣叫更易吸引捕食者,即雄性可能面临更大的被捕食压力,据此推测雄性可能更偏爱与组合声音竞争,从而吸引雌性并降低被捕食风险。
1 材料与方法
1. 1 研究地点和实验动物
2023年3月末—5月初(安徽树蛙繁殖期),在安徽省金寨县天马国家级自然保护区(31°10′—31°20′ N,115°20′—115°50′ E;海拔871 m)完成相关试验。本研究分别对雄性和雌性安徽树蛙进行回放试验和趋声性试验;为避免视觉干扰和人为噪声的影响,所有试验均在21:00—04:00进行。回放试验在雄蛙洞穴附近进行,趋声性试验在放置于安徽树蛙栖息地边缘的实验箱中进行。共记录32只雄性[体质量(4. 82 ± 1. 25)g,体长(36. 01 ± 6. 34) mm]和54 只雌性[体质量(10. 53 ± 2. 92) g,体长(46. 84 ±3. 53) mm]的行为响应数据。试验结束后,测量动物的体质量、体长并将其放置于全封闭的围栏内(5. 0 m × 1. 5 m × 1. 0 m),以免重捕获。每隔3 d向围栏内投放活蟋蟀;待所有试验结束后,拆除围栏。试验期间平均气温(10. 2 ± 2. 7)℃,相对湿度(75. 6 ±17. 7)%。本研究通过了安徽大学实验动物伦理与管理委员会的审查(IACUC(AHU)-2022-043)。
1. 2 声音刺激
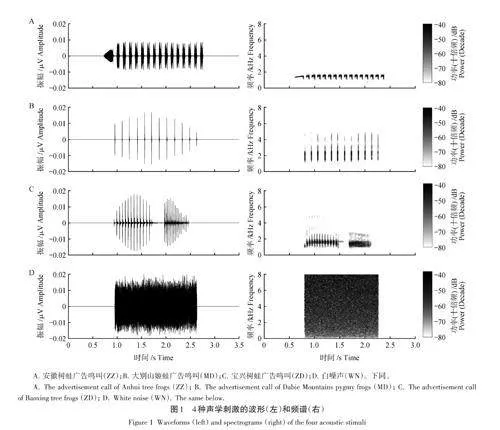
采用4种声音刺激(图1):同种广告鸣叫(ZZ)、同域分布的大别山姬蛙广告鸣叫(MD)、异域分布的宝兴树蛙广告鸣叫(ZD)和白噪声(white noise,WN,与MD等长,10 ms的淡入和淡出时长)。使用声压计(AWA 6291,爱华,杭州)将每个刺激的声压(sound pressure level,SPL)调整为(80. 0 ± 0. 5) dB(C计权,距离音箱1 m处测量);对后3种刺激另保存(68. 0 ± 0. 5)dB 的低幅度版本。ZZ 分别与另外3种刺激的低幅度版本组合形成组合刺激ZMD、ZZD和ZWN。由于在自然状态下,同种广告鸣叫和噪声一般来自空间的不同位置,而空间分离有助于信号感知[40];为更好地模拟自然状态,使用双音箱呈现组合鸣叫,即一只音箱播放ZZ,另一只音箱放置其后并抬高15 cm播放组合刺激的另一种声音。因为伪重复可能影响动物行为数据的统计分析结果[41],因此对每种鸣叫采用多个样本声音,从而消除伪重复的不良影响[39]。具体而言,对每种广告鸣叫(ZZ、MD和ZD),从数据集中分别选取来自4个不同个体且音节数量相同的鸣叫,从而形成4个刺激集,每个刺激集均包含ZZ、MD、ZD、WN、ZMD、ZZD 和ZWN 七种刺激(每种刺激各1个)。本研究对32只雄性进行回放试验,每个刺激集分别用于8只雄性。对于雌性趋声性试验,ZZ分别与其余6种刺激配对,形成6个刺激对;由于仅捕获54只雌性,因而随机选择4个刺激集中的2个刺激集分别用于14只雌性,另外2个刺激集分别用于13只雌性。
1. 3 雌性趋声性试验
实验箱(270 cm × 95 cm × 100 cm)内贴吸音棉,底部铺厚约5 cm的潮湿土壤。两只音箱(SME-AFS,Saul Mineroff Electronics,Elmont,NY,USA)等距放置于实验箱两端内侧,播放68 dB低幅度版本声音的音箱放置于某一侧音箱后上方。试验前,使用1 000 Hz纯音校准每个音箱的输出强度为80 dB。试验开始前,将雌性置于实验箱正中央半圆形透明塑料罩下,随机选择某个刺激对,以3. 5 s 刺激间隔(interstimulusinterval,ISI,安徽树蛙平均鸣叫间隔)轮唱;对于组合刺激而言,低幅度版本声音以1. 3 s的ISI(约等于MD和ZD两物种的平均鸣叫间隔)重复播放。刺激回放3 min后,通过滑轮拉起透明塑料罩,使用安装于实验箱两端的2 台红外摄像机(X4-IPC100WM,HawkSpee,China)观察并记录雌性的行为响应。若雌性移动至某音箱10 cm内的范围,视为其对该音箱所播放声音刺激的选择;若20 min内雌性未做出选择,中止试验。重复上述试验流程,直至所有刺激对均被随机选择1次。
1. 4 雄性回放试验
将音箱放置于距雄性洞穴1 m处并朝向洞穴。首先记录雄性在无声音刺激条件下的自发广告鸣叫6 min(对照条件control condition,CC);然后随机选取7种刺激之一回放6 min(ISI为3. 5 s,低幅度版本声音的ISI为1. 3 s)。使用麦克风(ME66,Sennheiser,Wedemark,Germany)记录回放刺激和雄性响应鸣叫。重复上述试验流程,直至所有刺激均被随机选择1次。
1. 5 数据处理与统计分析
对于雄性响应鸣叫,使用Adobe Audition 3. 0软件(San Jose,CA,USA)测量鸣叫的个数、每个鸣叫的音节数和持续时间;对每只雄性和每种声音刺激而言,求得每个参数的平均值。对于雌性试验,记录每种声学刺激被选择的次数和选择潜伏期(从拉开塑料罩到雌性做出选择之间的时间间隔)。
对所有数据进行正态性检验(Shapiro-Wilk W 检验)和方差齐性检验(Levene’s检验)。采用双因素(4个“刺激集”和8种“刺激类型”)重复测量ANOVA对雄性响应鸣叫的声学参数进行统计分析;因“刺激集”无显著主效应,表明伪重复不影响雄性试验的相关统计分析;采用单因素(“刺激类型”)重复测量ANOVA对雄性发声参数进行统计分析,并用Bonferroni事后检验进行多重比较分析。对于雌性选择次数,采用二项式检验(binomial test)评估不同刺激集或不同刺激之间的差异;前者无显著性差异,表明伪重复也不影响雌性试验的相关统计分析。对于每个刺激对,采用独立样本t 检验分析雌性选择潜伏期。所有统计分析使用SPSS 26(Illinois,USA)完成,以P lt; 0. 05为显著性水平。
2 结果
2. 1 雌性选择结果
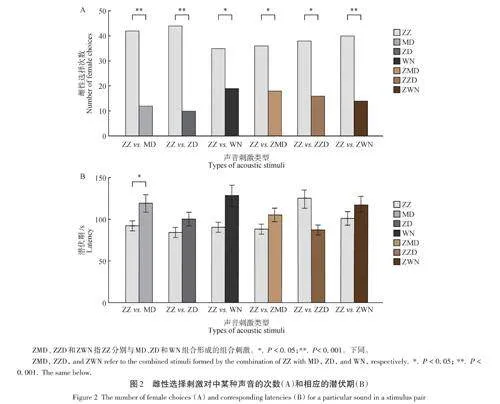
对所有刺激对而言,雌性更偏爱单纯的同种广告鸣叫。当ZZ与MD轮唱时,雌性显著偏爱ZZ(42 vs.12,P lt; 0. 001);当ZZ与ZD轮唱时,雌性显著偏爱于ZZ(44 vs.10,P lt; 0. 001);当ZZ与WN轮唱时,雌性显著偏爱前者(35 vs.19,P = 0. 040);当ZZ 与ZMD 轮唱时,雌性显著偏爱ZZ(36 vs.18,P = 0. 020);当ZZ与ZZD 轮唱时,雌性显著偏爱前者(38 vs.16,P =0. 004);当ZZ 与ZWN 轮唱时,雌性显著偏爱于ZZ(40 vs.14,P = 0. 001)(图2A)。当ZZ与MD轮唱时,雌性选择前者的潜伏期显著短于选择后者的潜伏期(P lt; 0. 05);对其他刺激对而言,雌性选择的潜伏期无显著差异(P gt; 0. 05)(图2B)。
2. 2 雄性响应鸣叫的声学参数
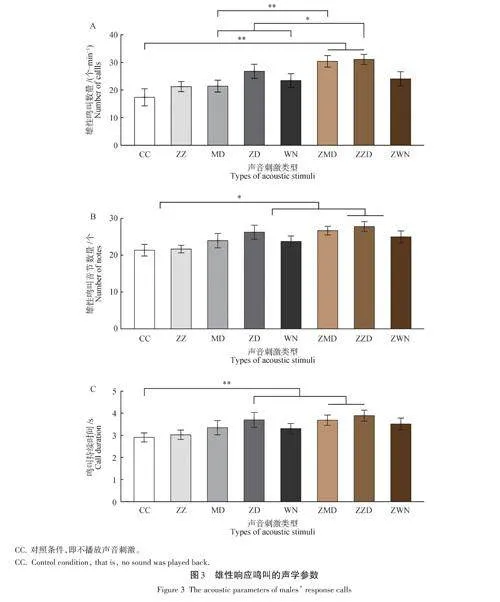
对于雄性响应鸣叫的数量而言,因素“刺激类型”的主效应显著(F(7,217) = 8. 624,P lt; 0. 001,partialη2 = 0. 218)。多重比较分析显示,ZMD和ZZD条件下的响应鸣叫数量显著多于对照条件(P lt; 0. 001),ZZD条件的响应鸣叫数量显著多于MD和WN条件(P lt; 0. 05),ZMD条件的响应鸣叫数量显著多于MD条件(P lt; 0. 001)(图3A)。对于雄性响应鸣叫的音节个数而言,因素“ 刺激类型”的主效应显著(F(7,217) = 6. 824,P lt; 0. 001,ε = 0. 738,partial η2 =0. 180)。多重比较显示,ZD、ZMD和ZZD条件下的音节个数显著多于对照条件(P lt; 0. 05),但ZD、ZMD和ZZD之间无显著差异(图3B)。对于雄性响应鸣叫的持续时间而言,因素“刺激类型”的主效应显著(F(7,217) = 8. 257,P lt; 0. 001,partial η2 = 0. 216)。多重比较显示,ZD、ZMD和ZZD条件下的鸣叫持续时间显著长于对照条件(P lt; 0. 001),但ZD、ZMD 和ZZD之间无显著差异(图3C)。
3 讨论
3. 1 雌性安徽树蛙偏爱信噪比高的同种广告鸣叫
本研究结果表明,与同域异种广告鸣叫、异域异种广告鸣叫、白噪声以及组合声音等刺激相比,雌性通常选择无噪声重叠的同种广告鸣叫。这一结果与既往研究发现的雌性通常偏爱信噪比更高的同种信号[39,42]一致。首先,在无尾两栖类合唱中,特定个体的广告鸣叫无法避免地与其他同种个体鸣声、异种个体鸣声或非生物环境噪声重叠,从而导致鸣叫精细成分损失[43−44],进而导致声音识别难度上升,即高水平的背景噪声可能会导致听觉掩蔽,降低雌性区分和识别鸣叫精细信息的能力,最终影响雌性选择和繁殖成功率[45]。因此雌性倾向选择非重叠的、信噪比较高的同种广告鸣叫[39,42,46−47]。其次,雌性在繁殖过程中的投入成本远高于雄性,在选择时更加谨慎[25,48]。若雌性因背景噪声干扰未能选择最适配偶,可能会退一步与低质量雄性交配,最终导致繁殖质量下降,因此雌性偏爱高信噪比的同种广告鸣叫可更好地确保选择更为合适的配偶。最后,雌性的发声器官欠发达,且无声囊,几乎不发出鸣叫[49],因而面临发声所致的被捕食风险远低于雄性,故无背景噪声重叠的同种广告鸣叫对雌性而言可能是最优选择。与此一致,当前结果发现雌性安徽树蛙始终偏爱信噪比高的同种广告鸣叫,而且整体而言选择该声音的潜伏期小于其他声音。此外,雌性选择同种广告鸣叫的潜伏期显著短于选择大别山姬蛙广告鸣叫的潜伏期,这可能是两者在同域分布,因此雌性能识别出相对熟悉的背景噪声从而更快地做出选择。
3. 2 雄性安徽树蛙偏爱响应被同域分布蛙类鸣声重叠的同种广告鸣叫
本研究结果表明,与无声音刺激相比,所用的所有声音刺激均能提高雄性安徽树蛙鸣叫数量。这一发现与既往研究[38−39]一致,表明雄性鸣叫行为具有高度可塑性,即可根据社会和环境因素的变化,动态优化竞争策略[50];亦表明雄性响应声音信号的容忍度高于雌性[25]。当前结果还发现,雄性安徽树蛙对包含同种广告鸣叫和异种广告鸣叫的组合声音有更强响应,组合声音可显著诱发雄性发声、增加鸣叫音节个数并延长鸣叫持续时间。一方面,大多数无尾两栖类雌性偏爱更长、更复杂的同种广告鸣叫,从而迫使雄性通过增加鸣叫数量和延长鸣叫长度等方式吸引雌性以确保繁殖成功[51−52];另一方面,雄性鸣叫在吸引同种雌性的同时,亦会导致雄性承受更高的被捕食压力[53−54],但高水平背景噪声可降低雄性所面临的被捕食风险[55]。本研究所用的组合声音(特别是包含同种广告鸣叫和异种广告鸣叫的组合声音)包含着两个在空间上分离的听觉目标,可以分散天敌注意力和降低天敌对特定个体的定位精度,从而降低被捕食风险。同种广告鸣叫和异种广告鸣叫的组合声音可能具备吸引雌性并能降低被捕食风险的双重功能,所以雄性安徽树蛙更偏爱与其竞争。此外,虽然对背景噪声的熟悉程度可能影响某些蛙类雄性个体的行为响应,但在本研究中雄性对于同域及异域异种广告鸣叫的行为响应之间无显著差异,即并未发现在其他蛙类中所发现的“dear enemy”现象,这可能是本试验所用的熟悉刺激并非是特定个体附近的雄性大别山姬蛙广告鸣叫。
总之,当前结果表明,在安徽树蛙中,雌性更偏爱响应信噪比高的同种广告鸣叫,雄性更偏爱响应被其他蛙类鸣声重叠的同种广告鸣叫;说明雌雄两性行为响应存在差异,这种差异可能源自自然选择和性选择双重压力的综合作用。
猜你喜欢
通信电源技术(2024年2期)2024-04-07 14:35:26
视野(2022年19期)2022-10-21 09:08:54
噪声与振动控制(2022年3期)2022-07-04 02:25:28
地震研究(2021年4期)2021-09-23 01:31:07
地震研究(2021年1期)2021-04-13 01:05:04
科技创新与应用(2020年32期)2020-11-09 03:14:14
地震研究(2017年3期)2017-11-06 23:41:55
地震研究(2017年3期)2017-11-06 23:38:05
电子技术与软件工程(2016年22期)2016-12-26 10:09:57
中国学术期刊文摘(2016年1期)2016-02-13 14:05:23
