
异色瓢虫黑化突变体ml在两种不同食物条件下的抗紫外线能力测定
2025-02-19 00:00:00李媛秦亚龙孙元星郝亚楠
植物保护 2025年1期

摘要
紫外线辐射对昆虫具有重要影响,表皮黑化程度不同的个体对紫外线辐射的抵抗能力不同,而食物是影响昆虫表皮黑化程度的一个重要因素。本研究以豌豆蚜Acyrthosiphon pisum与人工饲料为食料,通过测定异色瓢虫Harmonia axyridis黑化突变体(ml)和野生型(wt)各龄幼虫(1~4龄)经紫外线辐照处理后的存活率、发育历期及发育至成虫的体重等,评价不同食物条件下两种表型的异色瓢虫抗紫外线能力的差异,以及不同发育阶段接受紫外线辐照对其抗紫外线能力的影响。结果表明:1)对两种食物饲喂的异色瓢虫1~4龄幼虫进行紫外线照射,ml在多个发育阶段较wt具有更高的存活率,且其总发育历期大多也明显缩短,且ml型1龄幼虫经紫外线照射后发育至成虫时体重更重。2)低龄幼虫(1龄和2龄)接受紫外线照射,其后续发育阶段的存活率较低,且在1龄幼虫紫外线处理中ml的适合度表现明显优于wt;4龄幼虫接受紫外线照射后后续阶段存活率最高,但2种表型间没有显著差异。3)2种表型的异色瓢虫均以取食豌豆蚜的个体抗紫外线能力整体优于取食人工饲料的个体,食物因素对两种表型存活率的影响存在差异。综上,异色瓢虫黑化突变体的抗紫外线能力强于野生型,且受处理幼虫阶段及食物类型的影响。本研究结果为进一步探究异色瓢虫的抗逆能力及筛选适应不同环境的生防资源奠定了理论基础。
关键词
异色瓢虫;" 黑化突变体;" 存活率;" 发育历期;" 体重
中图分类号:
S 476.2
文献标识码:" A
DOI:" 10.16688/j.zwbh.2024055
收稿日期:" 20240129""" 修订日期:" 20240415
基金项目:
甘肃省青年科技基金计划(21JR7RA848);甘肃农业大学伏羲青年英才培养计划(Gaufx-03Y05)
致" 谢:" 参加本试验部分工作的还有江代礼、谭翰杰、张能和纪烨斌等同学,特此一并致谢。
* 通信作者
E-mail:
haoya_nan@126.com
#
为并列第一作者
Determination of the UV-resistance of a melanism mutant ml of Harmonia axyridis under two different food conditions
LI Yuan," QIN Yalong," SUN Yuanxing," HAO Yanan*
(College of Plant Protection, Gansu Agricultural University, Biocontrol Engineering Laboratory of Crop
Diseases and Pests of Gansu Province, Lanzhou" 730070, China)
Abstract
UV radiation significantly affects insects, but individuals with different degrees of cuticle melanism vary in their resistance to UV radiation. Food is an important factor affecting the cuticle melanism of insects. In this study, the melanism mutant (ml) and the wild type (wt) of Harmonia axyridis were reared with two different foods, i.e., pea aphid (Acyrthosiphon pisum) or artificial diet, and the larvae at different stages (1st instar to 4th instar) were exposed to UV. After exposure, the cumulative survival rate in subsequent stages, developmental duration, and adult body weight were measured to evaluate the UV-resistance of the two phenotypes as well as the influences of UV-exposure stage. The results showed that: 1) Under both foods conditions, the ml type versus wt type had a relatively higher survival rate in most development stages and, in most cases, had shortened developmental time after their 1st to 4th instar larvae exposed to UV. Meanwhile, ml had significantly higher body weight when the 1st instar larvae exposed to UV; 2) UV exposure of early-stage larvae (1st instar and 2nd instar) resulted in the lowest subsequent survival rates, but ml had significantly better fitness than wt in UV-exposure treatment of the 1st instar larvae. The highest survival rates were observed in UV-exposure treatment of the 4th instar larvae, but no significant difference was detected between the two phenotypes; 3) For both phenotypes, pea aphid-fed individuals exhibited higher UV-resistance than those feeding on artificial diet. However, food factor exhibited different influences on survivals of the two phenotypes. In conclusion, the melanism mutant demonstrated stronger UV-resistance than wt, and their UV-resistance affected by the UV-exposure stage and the food type. This study provides a theoretical foundation for further research on the stress resistance of H.axyridis and the identification of biocontrol agents adapted to different environments.
Key words
Harmonia axyridis;" melanism mutant;" survival rate;" developmental duration;" body weight
紫外线(ultraviolet,UV)是一种电磁辐射,可分为3种不同的波长类型:紫外线A(UVA: 320~400 nm)、紫外线B(UVB: 280~315 nm)和紫外线C(UVC: 200~280 nm),紫外线所携带的能量与波长成反比,即波长越短能量越高,对生物体造成的伤害也越严重[1]。UVB和UVC对生物的伤害较大,但部分UVB和全部UVC被大气层中的氧和臭氧吸收,因此进入地表环境的主要是UVA[2]。UVA对环境、植被、昆虫等产生诸多不良影响。
Rebollar等[3]的研究表明,紫外线能够引发生物体产生大量超氧阴离子、过氧化氢、羟自由基和单线态氧等对蛋白质和脂膜造成损伤的活性氧(reactive oxygen species, ROS),破坏昆虫正常的细胞结构和生理功能,从而带来存活率下降、发育历期延长、化蛹推迟及虫体质量减轻等诸多危害[1,4]。例如,低强度UVA辐射能明显延长果蝇的发育历期[5]。昆虫对紫外线辐射的抵抗能力受多重因素影响。首先,不同发育阶段昆虫对紫外线的抵抗能力不同。其中,一般未发育完全的幼龄昆虫更易受到紫外线直接或间接的影响[6]。此外,表皮黑色素含量对昆虫的紫外线抗性具有重要作用[78],昆虫黑色表皮相较于浅色表皮能更有效地抵御紫外线,如体色为白色的家蚕幼虫相比黑色家蚕幼虫对UVA照射更敏感[9]。在自然条件下,黑化是昆虫最为常见的体色变异现象[1011],但有关黑化突变体的抗紫外线能力仅有少量研究。
异色瓢虫Harmonia axyridis属鞘翅目Coleoptera瓢甲科Coccinellidae[12],可捕食多种害虫,尤其对各类蚜虫有较强的捕食能力,作为重要生防天敌在全球农业生产中被广泛应用[13]。异色瓢虫因其鞘翅的多样性而著称,是一种典型的多型性生物,其鞘翅底色大多由红黄色与黑色组成[14],其上分布黑色或黄色斑点以形成丰富的斑型变化。本实验室在室内饲养种群中发现了一体色异常黑化的突变体(ml)。该突变体各发育阶段的体色都呈现出明显的黑化特征,其典型表型如图1所示[15]。本研究以豌豆蚜Acyrthosiphon pisum与人工饲料为食料探究异色瓢虫ml各龄幼虫经紫外线辐照处理后后续阶段的存活率、发育历期及发育至成虫的体重的变化规律,并分析ml与野生型(wt)响应紫外线辐射的差异。旨在为进一步明确异色瓢虫的环境适应机制奠定基础。
1" 材料与方法
1.1" 材料
1.1.1" 两种食物及供试虫源
本试验所用异色瓢虫黑化型突变体(ml)为实验室内长期饲养种群,对照野生型(wt)为黄底型异色瓢虫,于2022年5月采自兰州市安宁区黄河北岸(103°34′E-103°47′E, 36°5′N-36°10′N)的黄刺玫Rosa xanthina上,在实验室长期饲养。2种表型的异色瓢虫均饲养于直径为9 cm的培养皿内,每皿4头,每天提供充足且新鲜的绿色型豌豆蚜Acyrthosiphon pisum,培养皿放置在人工气候箱[温度(25±1)℃,湿度60%~65%,光周期L∥D=14 h∥10 h]内。豌豆蚜饲养于盆栽蚕豆幼苗上,并置于人工气候箱[温度(20±1)℃,湿度60%~65%,光周期L∥D=14 h∥10 h]内繁殖种群以供后续试验使用。
1.1.2" 人工饲料的配制
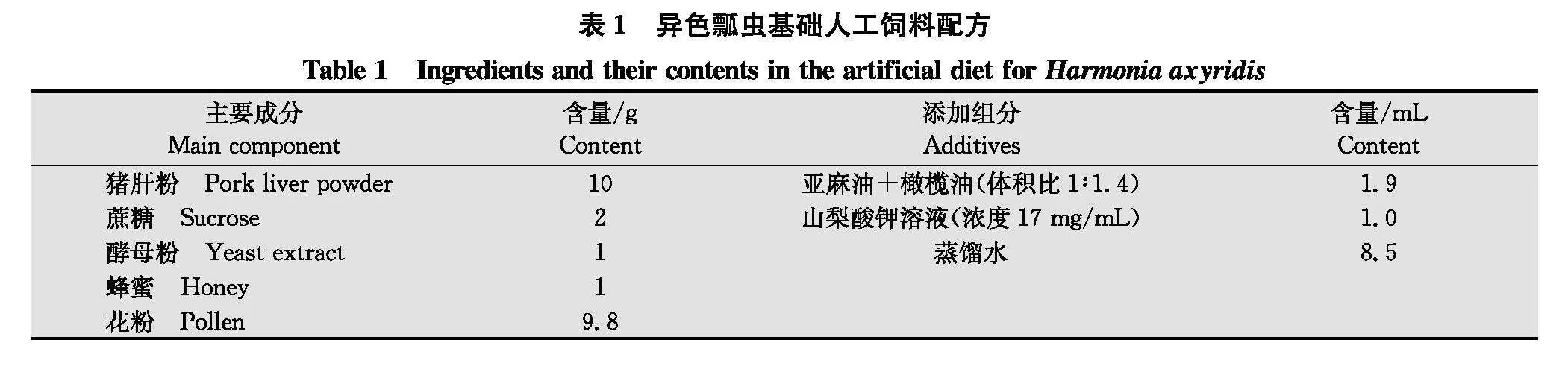
固体人工饲料参照前期研究所述配方及方法[1617]提前配制,并分装在2 mL离心管中置于-20℃冰箱内保存备用,饲料主要成分及含量见表1。
1.2" 方法
1.2.1" 幼虫饲养
2种表型的异色瓢虫成虫分别饲养在直径9 cm培养皿内,提供充足豌豆蚜,雌雄交配后将所产卵挑出置于直径3 cm培养皿内,并放置1个浸满蒸馏水的棉球用于保湿,2~3 d后卵开始孵化,将黑化型和野生型初孵幼虫分别置于直径为3 cm的培养皿内单头饲养,并分别使用人工饲料(AD)和豌豆蚜(AP)饲喂。用人工饲料喂养时,将人工饲料添加至边长为0.5 cm左右的硫酸纸上然后放入培养皿,并在培养皿中放入一浸湿蒸馏水的棉球保湿,2 d更换1次人工饲料,及时给棉球添水并更换培养皿以保持饲养环境干净。用豌豆蚜喂养时,每天提供充足且新鲜的豌豆蚜并及时清理培养皿。所用试验幼虫均饲养于人工气候箱内[温度(25±1)℃,湿度60%~65%,光周期L∥D=14 h∥10 h]。当幼虫发育至所需阶段后进行抗紫外线能力测定。
1.2.2" 紫外线处理不同龄期幼虫
本试验分别对1龄(L1)或2龄(L2)、3龄(L3)和4龄(L4)幼虫进行紫外线辅照处理,记为UV-L1、UV-L2、UV-L3及UV-L4。每个龄期共计处理25头幼虫。准备一方形泡沫塑料盒(28 cm × 16 cm × 18 cm),在其顶部按照手提式紫外线分析仪(365 nm,WFH-204B,杭州齐威仪器有限公司)滤光片的大小裁剪出一长方形开口(16.5 cm × 6.5 cm),并将手提式紫外线分析仪置于泡沫盒上。紫外线处理时,用橡皮筋将纱网固定在每个装有试验幼虫的培养皿(1头/皿)的外壁上,将培养皿转移至泡沫盒内,并将紫外线分析仪的光源对准纱网。紫外灯距培养皿8 cm,持续照射4 h后将供试幼虫挑出并转移至新的培养皿中继续单头饲喂。每天观察幼虫的存活及发育情况,统计经紫外线处理的幼虫发育至后续各阶段的累计存活率[发育至各阶段的存活数/初始幼虫数(25)×100%]、后续各阶段的发育历期以及新羽化成虫的体重(羽化后24 h)。
1.3" 数据分析
采用SPSS 26软件进行数据分析。其中,紫外线处理不同龄期幼虫后的累计存活率采用4×2列联表法进行卡方检验,显著水平均设置为0.05;当检测到显著差异时,各处理进一步通过2×2列联表法依次进行两两比较[18];发育历期、新羽化成虫体重采用独立样本t检验进行差异显著性比较,显著水平均设置为0.05。使用Excel作图。
2" 结果与分析
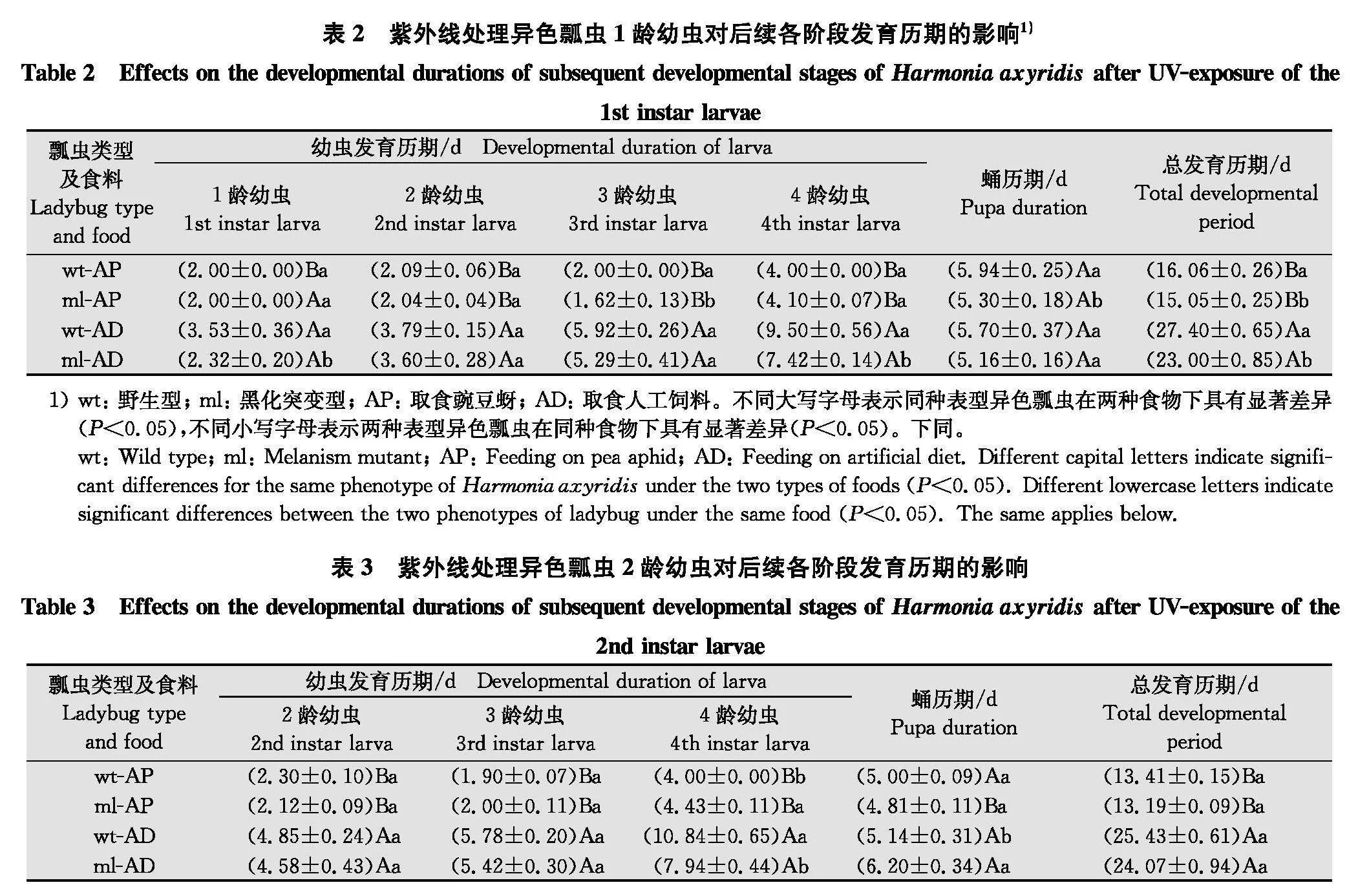
2.1" 紫外线处理不同龄期幼虫后后续不同发育阶段试虫的累计存活率
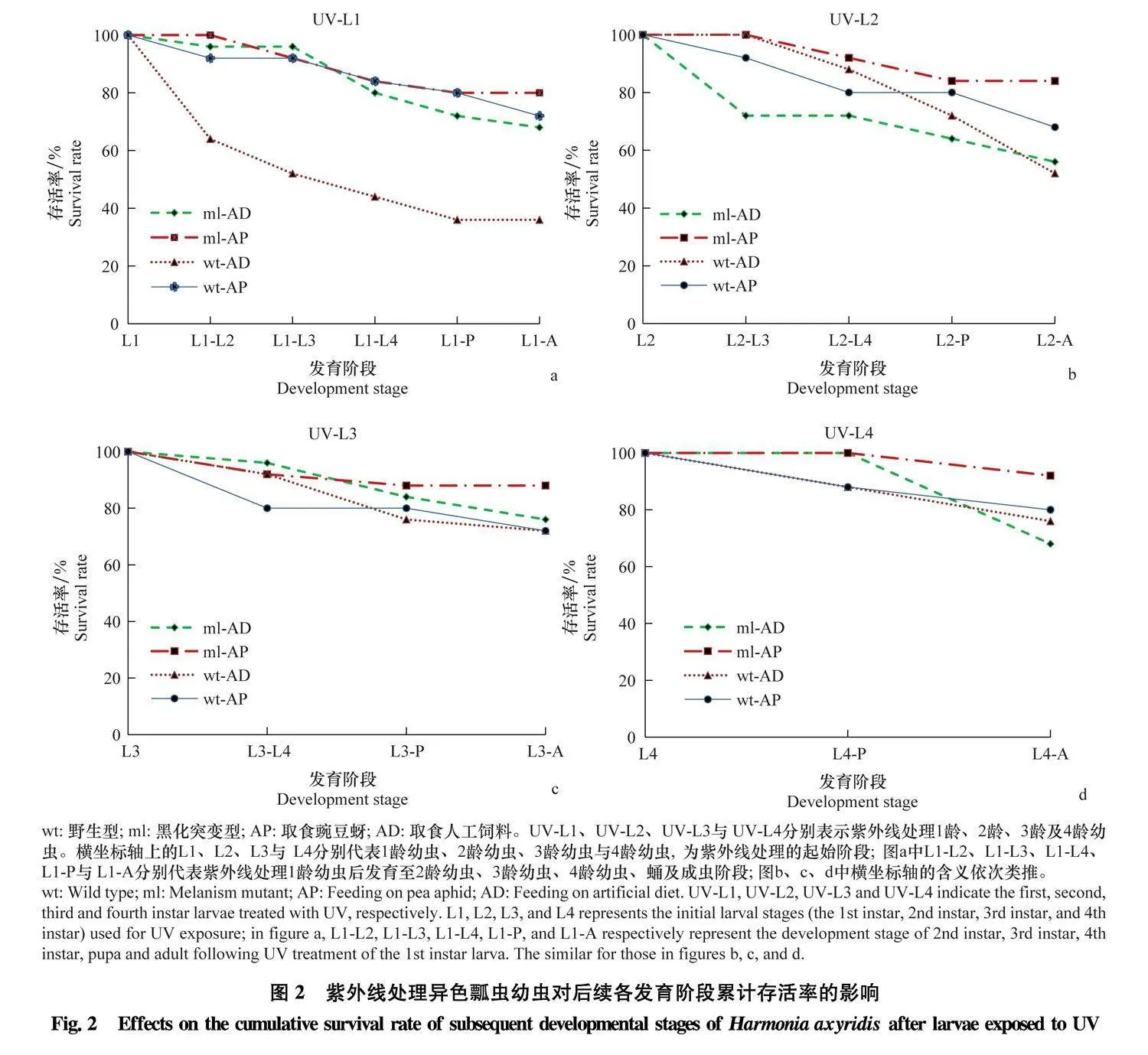
取食豌豆蚜的幼虫经紫外线处理后,其发育至后续阶段的累计存活率均逐渐降低,黑化突变体(ml)幼虫发育至各阶段的累计存活率均稍高于野生型(wt),但无显著差异(Pgt;0.05)。其中,ml型1龄幼虫和wt型2龄幼虫经紫外线照射后发育至成虫的存活率最低,分别为80%与68%(图2a,b);而ml型和wt型的4龄幼虫接受紫外线照射后发育至成虫的存活率最高,分别为92%和80%(图2d)。
取食人工饲料的幼虫经紫外线照射后,其发育至后续阶段的累计存活率也均逐渐降低。其中,1龄幼虫接受紫外线照射后,ml型发育至2龄(χ2=8.000, P=0.005)、3龄(χ2=12.578, Plt;0.001)、4龄(χ2=6.522, P=0.011)、蛹(χ2=6.876, P=0.009)及成虫(χ2=5.128, P=0.024)的存活率均显著高于wt型(图2a)。但2龄幼虫接受紫外线照射后,ml型发育至3龄的累计存活率显著低于wt型(χ2=8.140, P=0.004)(图2b)。3龄及4龄幼虫接受紫外线处理后,ml型发育至后续各阶段的累计存活率均略高于wt,但两者间无显著差异(Pgt;0.05)。就发育至成虫的累计存活率来说,ml型2龄幼虫及wt型1龄幼虫接受紫外线照射后存活率最低,分别为56%与36%(图2a,b);ml型3龄幼虫及wt型4龄幼虫接受紫外线照射后存活率最高,均为76%(图2c,d)。
比较取食豌豆蚜和人工饲料的异色瓢虫的抗紫外能力发现,ml型1龄幼虫接受紫外线照射后发育至后续各阶段的累计存活率均没有显著差异(Pgt;0.05),但取食豌豆蚜的ml型2龄幼虫接受紫外线照射后发育至3龄(χ2=8.140, P=0.004)和成虫的累计存活率(χ2=4.667, P=0.031)及其4龄幼虫接受紫外线照射后发育至成虫的累计存活率(χ2=4.500, P=0.034)显著高于取食人工饲料的个体;取食豌豆蚜的wt型1龄幼虫接受紫外线照射后发育至2龄幼虫(χ2=5.711, P=0.017)、3龄幼虫(χ2=9.921, P=0.002)、4龄幼虫(χ2=8.681, P=0.003)、蛹(χ2=9.934, P=0.002)及成虫(χ2=6.522, P=0.011)的累计存活率均显著高于取食人工饲料的个体(图2a)。在其他龄期幼虫接受紫外线照射,同种表型异色瓢虫经2种食物饲养,个体的存活率均无显著差异(Pgt;0.05)(图2b~d)。
2.2" 紫外线处理不同龄期幼虫后后续发育阶段的发育历期
当紫外线处理1龄幼虫时,2种食物条件下ml发育至成虫的总发育历期均显著短于wt(Plt;0.05),且取食豌豆蚜时ml型3龄幼虫与蛹的发育历期显著短于wt型(Plt;0.05);取食人工饲料时ml型1龄与4龄幼虫的发育历期也显著短于wt型(Plt;0.05)。此外,比较取食两种食物的幼虫发育历期,发现取食豌豆蚜的ml型和wt型幼虫总发育历期均显著短于取食人工饲料的个体(Plt;0.05)(表2)。
当紫外线处理2龄幼虫时,2种食物条件下ml发育至成虫的总发育历期与wt型个体均无显著差异(Pgt;0.05)。取食人工饲料的ml型个体的4龄幼虫发育历期显著短于wt型个体(Plt;0.05),但豌豆蚜饲喂处理的4龄幼虫历期以及人工饲料饲喂处理的蛹期显著长于wt(Plt;0.05)。此外,取食豌豆蚜的ml型和wt型异色瓢虫总发育历期显著短于取食人工饲料的个体(Plt;0.05)。取食2种食物的wt型异色瓢虫蛹期无显著差异(Pgt;0.05)(表3)。
当紫外线处理3龄幼虫时,ml取食豌豆蚜时发育至成虫的总发育历期与wt间无显著差异(Pgt;0.05),而取食豌豆蚜时的4龄及取食人工饲料时的3龄幼虫发育历期及发育至成虫的总历期均显著短于wt(Plt;0.05);而当紫外线处理4龄幼虫时,ml取食豌豆蚜与人工饲料时各阶段的发育历期及发育至成虫的总历期均与wt间无显著差异(Pgt;0.05)。此外,除UV-L3处理中wt的蛹期外,2种表型异色瓢虫取食豌豆蚜后各阶段发育历期及总发育历期均显著短于取食人工饲料的个体(Plt;0.05)(表4)。
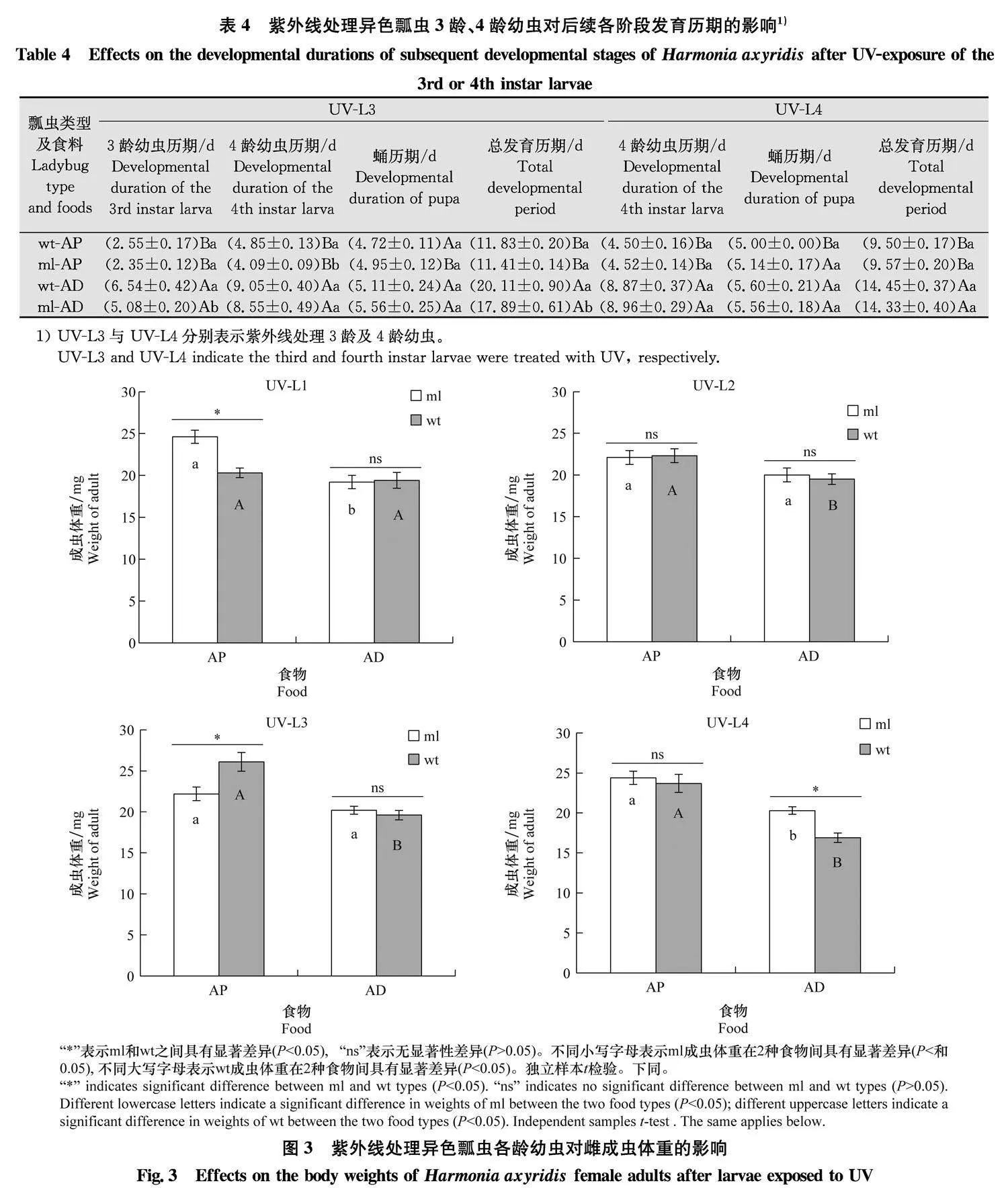
2.3" 紫外线处理不同龄期幼虫后新羽化成虫的体重
取食不同食物情况下,取食豌豆蚜的ml型异色瓢虫的1龄幼虫(t=-4.726, Plt;0.001)与4龄幼虫(t=-3.625, P=0.002),wt型的2龄幼虫(t=-2.627, P=0.015)、3龄幼虫(t=-5.676, Plt;0.001)与4龄幼虫(t=-8.727, Plt;0.001)经紫外线处理后发育至成虫的雌虫体重均显著高于取食人工饲料的个体。当取食豌豆蚜时,1龄幼虫接受紫外线照射后ml型个体发育至成虫的体重显著高于wt(t=4.369, Plt;0.001),但3龄幼虫接受紫外线照射后ml型发育至成虫的体重显著低于wt(t=-2.819, P=0.011)。当取食人工饲料时,ml型4龄幼虫接受紫外线照射后发育至成虫的体重显著高于wt(t=3.120, P=0.005),但其他龄期的幼虫紫外线处理后2个表型发育至成虫的体重均无显著差异(Pgt;0.05)(图3)。
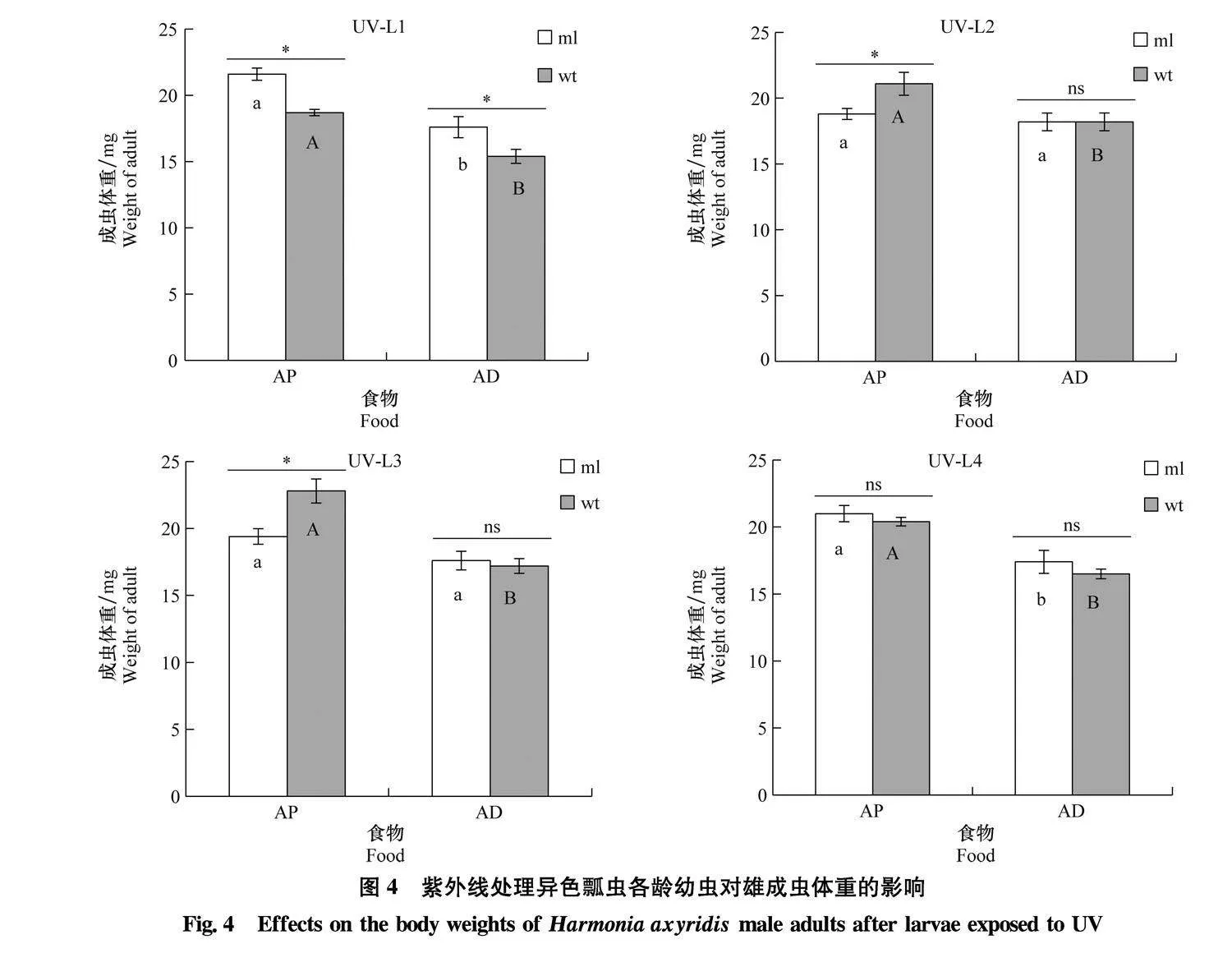
取食不同食物情况下,取食豌豆蚜的ml型异色瓢虫的1龄幼虫(t=-4.390,Plt;0.001)与4龄幼虫(t=-3.534,P=0.003),wt型1龄幼虫(t=-5.758,Plt;0.001)、2龄幼虫(t=-2.584,P=0.017)、3龄幼虫(t=-5.652,Plt;0.001)与4龄幼虫(t=-7.503,Plt;0.001)经紫外线照射后发育得到的雄成虫体重显著高于取食人工饲料的个体。当取食豌豆蚜时,ml型1龄幼虫接受紫外线照射的处理中发育至雄成虫的体重显著高于wt(t=5.384,Plt;0.001),但在2龄幼虫(t=-2.587,P=0.019)与3龄幼虫(t=-3.250,P=0.004)接受紫外线照射的处理中雄成虫体重显著低于wt。当取食人工饲料时,ml型1龄幼虫接受紫外线照射的处理中雄成虫体重显著高于wt(t=2.340,P=0.028),但2个表型在其他龄期幼虫紫外线暴露处理后获得的雄成虫体重均无显著差异(Pgt;0.05)(图4)。
3" 结论与讨论
紫外线处理2种表型幼虫后,异色瓢虫黑化突变体(ml)较野生型(wt)的存活率升高,总发育历期缩短,1龄幼虫接受紫外线照射后发育至成虫的体重更高。前期研究发现,ml与wt在正常条件下的主要生物学表现无显著差异[15],由此说明,ml抗紫外线的能力强于wt。对不同表型果蝇的研究也获得了类似的结果,正常型果蝇Drosophila melanogaste相比黄化型果蝇的抗紫外线能力更强[1920]。两种表型抗紫外线能力差异可能是由于ml型个体体内的黑色素含量较wt高或表皮厚度不同,对其进行了有效保护。紫外线照射通常会引起昆虫外骨骼黑色素含量增高及角质层增厚[1],从而提高其免疫力,达到光保护作用。相关功能已在部分昆虫中证实,如天蓝细蟌Coenagrion puella被紫外线照射后体内的黑色素合成能力相对增强[21],而赤拟谷盗Tribolium castaneum被紫外线照射后外表皮变厚[22]。此外,紫外线辐射会引起昆虫产生大量氧自由基,但体内的黑色素可通过螯合自由基来抵挡紫外线辐射对昆虫造成的生理和外部形态上的损伤[2324]。
本研究进一步发现,2种表型的异色瓢虫均在紫外线处理低龄幼虫时(1龄和2龄幼虫)引起的后续阶段存活率最低,而紫外线处理高龄幼虫(4龄幼虫)引起的后续阶段存活率最高。同时,ml型1龄幼虫经紫外线处理后,后续阶段的适合度表现明显优于wt(存活率及发育至成虫的体重显著增高,而发育历期显著缩短);而4龄幼虫经紫外线处理,ml与wt间的生物学特性无显著差异。上述结果表明,紫外线照射处理不同龄期幼虫对2种表型异色瓢虫的生物学表现具有不同的影响。通常,紫外线对昆虫卵的伤害最大,幼虫次之,成虫最小[1]。Ali等[25]在东方黏虫Mythimna separata抗紫外线研究中也发现UVA辐射对低龄幼虫的危害大于高龄幼虫。这种差异主要是由于高龄幼虫体内蛋白质和脂质等含量较高,可在免疫防御紫外线辐射等不利环境中发挥重要作用[2627]。本研究还发现紫外线处理某一龄期幼虫后,2个表型在该阶段的生物学表现无差异,但在后续的某些发育阶段表现出明显差异,如紫外线处理1龄幼虫后,其1龄幼虫发育历期在2种表型间无差异,但在3龄及蛹期却表现出明显差异。上述结果表明,紫外线处理造成的不利影响是随昆虫生长发育逐渐表现出来的,且可能存在累加效应。Ali等[6]研究发现UVA辐射东方黏虫后导致其成虫寿命缩短但产卵率升高。此外,张雅君等[28]发现紫外线辐射雄性果蝇可能会引发基因突变,即亲代受到紫外线辐射后造成子代表现异常的现象。因此,后续研究还需测定紫外线辐射处理不同龄期幼虫对异色瓢虫黑化突变体ml成虫生殖力与子代生长发育的影响及与野生型wt的差异。
同时,研究发现喂饲不同食物的异色瓢虫幼虫经紫外线辐射后后续阶段的生物学特征表现出明显差异。从整体来看,取食豌豆蚜的ml和wt型个体较取食人工饲料的个体具有更高的存活率,更短的发育历期。与此同时,当取食人工饲料时,ml型在部分紫外线处理中的表现优于野生型wt(1龄幼虫处理中的存活率及4龄幼虫处理中的雌成虫体重)。造成上述差异的原因可能是与人工饲料相比豌豆蚜营养更为丰富[29],更有利于异色瓢虫生长发育[30]。特别是,与抗紫外线有关的黑色素的合成需要从食物中获取蛋白质等原料[31],而2种食物中蛋白质的含量可能存在较大差异。同时,本试验研究结果也表明在人工饲料条件下ml较wt对食物的利用效率更高,以利于其提升保护性酶的含量等[32]。昆虫体色影响其食物利用效率的研究已有相关报道,如Ethier等[33]发现天幕毛虫Malacosoma neustria取食相同食物后黑化型较正常型的雄虫体型变小。
本试验研究了UVA对2种表型异色瓢虫幼虫生长发育的影响,结果表明黑化突变体较野生型具有更强的抗紫外线能力且取食人工饲料可进一步提升黑化突变体的抗紫外线优势。此外,本实验室前期已有研究发现,取食豌豆蚜后黑化突变体的抗干旱能力也明显优于野生型(未发表)。由上可知,黑化突变体已表现出良好的自然环境适应能力,但对其他环境因子如极端温度[3435]的适应力还有待进一步评价。同时,本试验也发现黑化突变体对蚜虫的日捕食量很高(成虫体重与野生型相当或稍高),且前期研究发现其幼虫取食人工饲料后的适合度表现优于野生型,具有利用人工饲料饲养的重要优势[15]。综上,黑化突变体是具有重要应用潜能的生防资源,可依据不同地区的紫外线辐照、干旱程度等条件合理使用。同时,本文基于食物因子影响抗紫外线能力的结果也可为科学饲养适于不同环境的天敌昆虫产品提供有益指导。参考文献
[1]" 王姿烨, 王飞凤, 何万财, 等. UV对昆虫的生物学效应及昆虫适应机制[J]. 应用昆虫学报, 2022, 59(1): 1628.
[2]" WANG Shaobin, SU Weihan, WEI Dingwen. Biologically effective radiation of solar ultraviolet radiatin and the depletion of ozone layer [J]. Acta Scientiae Circumstantiae, 1993, 13(1): 114119.
[3]" REBOLLAR E, BOUNOS G, OUJJA M, et al. Influence of polymer molecular weight on the chemical modifications induced by UV laser ablation [J]. Journal of Physical Chemistry B, 2006, 110(29): 1421514220.
[4]" ZHANG Changyu, MENG Jianyu. Identification of differentially expressed proteins in Ostrinia furnacalis adults after exposure to ultraviolet A [J]. Environmental Science and Pollution Research, 2018, 25: 2507125079.
[5]" 郑俊丽. UVA照射对黑腹果蝇生物学特性和抗氧化反应的影响[D]. 武汉: 华中农业大学, 2011.
[6]" ALI A, RASHID M A, HUANG Qiuying, et al. Effect of UV-A radiation as an environmental stress on the development, longevity, and reproduction of the oriental armyworm, Mythimna separata (Lepidoptera: Noctuidae) [J]. Environmental Science and Pollution Research International, 2016, 23(17): 1700217007.
[7]" FUTAHASHI R, FUJIWARA H. Juvenile hormone regulates butterfly larval pattern switches [J/OL]. Science, 2008, 319(5866): 1061. DOI: 10.1126/science.1149786.
[8]" TALLOEN W, DYCK H V, LENS L. The cost of melanization: butterfly wing coloration under environmental stress [J]. Evolution, 2004, 58(2): 360366.
[9]" HU Yonggang, SHEN Yihong, ZHANG Ze, et al. Melanin and urate act to prevent ultraviolet damage in the integument of the silkworm, Bombyx mori [J]. Archives of Insect Biochemistry and Physiology, 2013, 83(1): 4155.
[10]WITTKOPP P J, CARROLL S B, KOPP A. Evolution in black and white: genetic control of pigment patterns in Drosophila [J]. Trends in Genetics, 2003, 19(9): 495504.
[11]WITTKOPP P J, BELDADE P. Development and evolution of insect pigmentation: Genetic mechanisms and the potential consequences of pleiotropy [J]. Seminars in Cell and Developmental Biology, 2009, 20(1): 6571.
[12]ROBERTSON J A, SLIPINSKI A, MOULTON M, et al. Phylogeny and classification of Cucujoidea and the recognition of a new superfamily Coccinelloidea (Coleoptera: Cucujiformia) [J]. Systematic Entomology, 2015, 40(4): 745778.
[13]王甦, 张润志, 张帆. 异色瓢虫生物生态学研究进展[J]. 应用生态学报, 2007, 18(9): 21172126.
[14]BEZZERIDES A L, LOOFBOURROW S A. Melanic facial patterns and their significance in the multicolored Asian lady beetle (Harmonia axyridis) [J]. The American Midland Naturalist, 2014, 172(2): 366371.
[15]李明凌, 王森山, 郝亚楠, 等. 不同突变体异色瓢虫取食人工饲料的适合度研究[J]. 西北农业学报, 2022, 31(11): 15071513.
[16]孙元星, 陈明娟, 李明凌, 等. 人工饲料添加花粉对异色瓢虫幼虫生长发育的影响[J]. 中国生物防治学报, 2023, 39(5): 10581065.
[17]SUN Yuanxing, HAO Yanan, LIU Tongxian. A β-carotene-amended artificial diet increases larval survival and be applicable in mass rearing of Harmonia axyridis [J]. Biological Control, 2018, 123(1): 105110.
[18]SIGHINOLFI L, FEBVAY G, DINDO M L, et al. Biochemical content in fatty acids and biological parameters of Harmonia axyridis reared on artificial diet [J]. Bulletin of Insectology, 2013, 66(2): 283290.
[19]DRAPEAU M D, RADOVIC A, WITTKOPP P J, et al. A gene necessary for normal male courtship, yellow, acts downstream of fruitless in the Drosophila melanogaster larval brain [J]. Journal of Neurobiology, 2010, 55(1): 5372.
[20]DRAPEAU M D, CYRAN S A, VIERING M M, et al. A cis-regulatory sequence within the yellow locus of Drosophila melanogaster required for normal male mating success [J]. Genetics Society of America, 2005, 172(2): 10091030.
[21]DEBECKER S, SOMMARUGA R, MAES T, et al. Larval UV exposure impairs adult immune function through a trade-off with larval investment in cuticular melanin [J]. Functional Ecology, 2015, 29(10): 12921299.
[22]LIU Yanmei, YU Lin, ALI S, et al. Effects of UVA radiation on cuticle microstructure of Tribolium castaneum [J]. 广东农业科学, 2018, 45(6): 8489.
[23]RIEDEL F, VORKEL D, EATON S. Megalin-dependent yellow endocytosis restricts melanization in the Drosophila cuticle [J]. Development, 2011, 138(1): 149158.
[24]杨松. 昆虫黑化反应的分子机制研究进展[J]. 国外医学:寄生虫病分册, 2003, 30(4): 164168.
[25]ALI A, RASHID M A, HUANG Qiuying, et al. Influence of UV-A radiation on oxidative stress and antioxidant enzymes in Mythimna separata (Lepidoptera: Noctuidae) [J]. Environmental Science and Pollution Research, 2017, 24(9): 83928398.
[26]RAVANAT J L, DOUKI T, CADET J. Direct and indirect effects of UV radiation on DNA and its components [J]. Journal of Photochemistry and Photobiology B: Biology, 2001, 63(1/2/3): 88102.
[27]CADET J, SAGE E, DOUKI T. Ultraviolet radiation-mediated damage to cellular DNA [J]. Mutation Research, 2005, 571(1/2): 317.
[28]张雅君, 梁佳勇, 曾慕衡. 紫外线对雄果蝇寿命及子代生理的影响[J]. 天津农业科学, 2013, 19(7): 7578.
[29]WOLF S, ROMEIS J, COLLATZ J. Utilization of plant-derived food sources from annual flower strips by the invasive harlequin ladybird Harmonia axyridis [J]. Biological Control, 2018, 122: 118126.
[30]张屾.非昆虫源人工饲料对异色瓢虫生物学、生化特性及捕食行为影响的研究[D]. 北京: 中国农业科学院, 2014.
[31]QIAO Liang, LI Yuanhao, XIONG Gao, et al. Effects of altered catecholamine metabolism on pigmentation and physical properties of sclerotized regions in the silkworm melanism mutant [J/OL]. PLoS ONE, 2012, 7(8): e42968. DOI: 10.1371/journal.pone.0042968.
[32]周冬, 杜一民, 杨杰, 等. 连续多代UV-B照射对麦长管蚜保护酶活性的影响[J]. 昆虫学报, 2014, 57(7): 762768.
[33]ETHIER J, DESPLAND E. Effects of polymorphic melanism and larval diet on life history traits of Malacosoma disstria moths [J]. Journal of Insect Physiology, 2012, 58(1): 6774.
[34]伍兴隆, 余佳敏, 刘国, 等. 高温处理对七星瓢虫生长发育及繁殖的影响[J]. 昆虫学报, 2024, 67(2): 223234.
[35]赵锋, 卜亚莉, 韩慧, 等. 阶段性极端高温对七星瓢虫生长发育和繁殖的影响[J]. 植物保护学报, 2023, 50(1): 129135.
(责任编辑:杨明丽)
猜你喜欢
戏曲研究(2022年4期)2022-06-27 07:06:08
湖北农业科学(2021年22期)2021-12-13 13:00:36
河北果树(2020年4期)2020-11-26 06:04:48
红蜻蜓·低年级(2020年9期)2020-09-26 11:49:22
小学生作文(低年级适用)(2018年4期)2018-11-29 02:24:31
中国稻米(2017年2期)2017-04-28 08:00:06
中国中西医结合皮肤性病学杂志(2016年4期)2016-07-18 10:59:56
中国稻米(2016年2期)2016-06-29 09:53:29
中国茶叶(2016年7期)2016-01-18 00:34:28
小朋友·快乐手工(2015年10期)2015-11-02 00:34:08
