
外源茉莉酸甲酯诱导脐橙果实抗青霉病与活性氧代谢的关系
2024-12-31 00:00:00陈明郑致远王印宝郭越蒋海燕付永琦曾教科陈金印向妙莲
果树学报 2024年12期







摘" " 要:【目的】通过探究外源茉莉酸甲酯(Methyl Jasmonate,MeJA)诱导脐橙果实采后抗青霉病与活性氧代谢的关系,为MeJA应用于控制果实采后病害提供理论依据。【方法】以纽荷尔和龙回红脐橙果实为试材,在温度(26±1) ℃、湿度85%~95%的环境下,使用浓度为50 μmol·L-1 MeJA密闭熏蒸处理24 h后,接种青霉病菌Penicillium italicum(1.25×106 spores·mL-1)20 µL,以无菌水处理为对照,另设100 μmol·L-1水杨苷异羟肟酸(salicyhydroxamic acid,SHAM)喷施处理,测定脐橙果实活性氧代谢相关指标并分析关键酶基因表达水平。【结果】50 μmol·L-1 MeJA熏蒸处理显著提高2个脐橙品种果实的谷胱甘肽还原酶(GR)、抗坏血酸过氧化物酶(APX)、超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、单脱氢抗坏血酸还原酶(MDHAR)和脱氢抗坏血酸还原酶(DHAR)活性以及对应酶基因CsGR、CsAPX、Cu-ZnSOD、CsCAT、CsMDHAR、CsDHAR3的相对表达量,在纽荷尔中MeJA处理分别为对照的1.10、1.16、1.12、1.69、1.21、1.43、1.16、1.42、1.56、1.36、1.58、2.32倍,在龙回红中MeJA处理分别为对照的1.13、1.49、1.02、1.41、1.26、1.79、2.98、1.59、1.10、1.79、1.57、1.31倍;同时,显著提高了还原型谷胱甘肽(GSH)、抗坏血酸(AsA)含量和DPPH自由基清除率,降低了过氧化氢(H2O2)含量和超氧阴离子(O2.-)产生速率,延缓了丙二醛(MDA)含量的积累。【结论】使用MeJA于接种前24 h熏蒸脐橙果实,能提高柑橘果实活性氧代谢相关酶活性以及关键酶基因的表达量,增强果实对青霉病的抗性。
关键词:脐橙果实;茉莉酸甲酯;青霉病抗性;活性氧代谢
中图分类号:S666.4 文献标志码:A 文章编号:1009-9980(2024)12-2555-12
Involvement of induced resistance by methyl jasmonate to blue mold and reactive oxygen species metabolism in navel orange fruit
CHEN Ming, ZHENG Zhiyuan#, WANG Yinbao, GUO Yue, JIANG Haiyan, FU Yongqi, ZENG Jiaoke, CHEN Jinyin, XIANG Miaolian*
(College of Agronomy, Jiangxi Agricultural University/Jiangxi Provincial Key Laboratory of Fruit and Vegetable Storage and Preservation/Collaborative Innovation Center of Postharvest Key Technology and Quality Safety of Fruits and Vegetables in Jiangxi Province, Nanchang 330045, Jiangxi, China)
Abstract: 【Objective】 Navel orange [Citrus sinensis (Linn.) Osbeck ], one of the most widely planted citrus fruit trees in the world, is mainly planted in Chongqing, Hubei, Jiangxi, Fujian and other places in southern China. However, navel orange fruits are susceptible to diseases caused by pathogens during postharvest transportation and storage, seriously affecting the quality and economic value of the fruits. The blue mold caused by Penicillium italicum is one of the most important postharvest diseases in navel orange fruits, which is currently mainly controlled by chemicals. Currently there is an urgent need to develop safer and more efficient preservation techniques to control post-harvest diseases of navel orange fruits. Methyl jasmonate (MeJA) is an important natural plant growth regulator, which can be used as an inducer of resistance to pathogen infestation to produce a defense response in plants. In recent years, the aspect of MeJA-induced disease resistance in plants has become a hot issue, but there are fewer reports on MeJA-induced resistance to P. italicum in navel orange fruits. In this study, we investigated the relationship between exogenous MeJA-induced postharvest resistance to the blue mold and reactive oxygen metabolism in navel orange fruits to provide a theoretical basis for the potential of the application of MeJA to control postharvest fruit diseases. 【Methods】 In this study, Newhall and Longhuihong navel orange fruits were used as test materials. Firstly, the fruits were cleaned with 0.05% sodium hypochlorite and water, then after being dried at room temperature, one group fruits were treated with 50 μmol·L-1 MeJA for 24 h at (26±1) ℃ and 85%-95% relative humidity, another group fruits were sprayed with 100 μmol·L-1 SHAM, and the third group treated with sterile water was used as control. The conidia of P. italicum that had been cultured on PDA medium for one week were collected and made into a spore suspension with a concentration of 1.25×106 spores·mL-1. The surface of the fruits was sterilized with 75% ethanol and inoculation was carried out by puncturing the equatorial part of the fruit at equal distances using an inoculation needle with a wound size of 2 mm in diameter and 3 mm in depth and injecting 20 µL of spore suspension of P. italicum. Ten fruits were selected from each treatment for 7 consecutive days, and 1 cm of peel tissue was taken from the disease-health junction of navel oranges and quickly frozen with liquid nitrogen for further analysis. The activities of glutathione reductase (GR), ascorbate peroxidase (APX), superoxide dismutase (SOD), catalase (CAT), monodehydroascorbate reductase (MDHAR), dehydroascorbate reductase (DHAR), and the relative expression levels of related genes including CsGR, CsAPX, Cu-ZnSOD, CsCAT, CsMDHAR and CsDHAR3 were measured. All data were statistically organized and analyzed by one-way analysis of variance (ANOVA) using Excel 2018 and SPSS 20.0 software. The origin 2018 software was used for graphing, and all data in the graphs were expressed as mean ± standard error (SE). 【Results】 MeJA fumigation treatment significantly enhanced the activities of GR, APX, SOD, CAT, MDHAR, DHAR and the relative expression levels of CsGR, CsAPX, Cu-ZnSOD, CsCAT, CsMDHAR and CsDHAR3 in two navel orange varieties fruits. In the MeJA treatment of Newhall, they were 1.10, 1.16, 1.12, 1.69, 1.21, 1.43, 1.16, 1.42, 1.56, 1.36, 1.58 and 2.32 times as much as the control, respectively. In the MeJA treatment of Longhuihong, they were 1.13, 1.49, 1.02, 1.41, 1.26, 1.79, 2.98, 1.59, 1.10, 1.79, 1.57 and 1.31 times as much as the control, respectively. At the same time, the content of reduced glutathione (GSH), ascorbic acid (AsA) and DPPH free radical scavenging rate were significantly increased, the content of hydrogen peroxide (H2O2) and the production rate of superoxide anion (O2.-) were decreased, and the accumulation of malondialdehyde (MDA) content was delayed. 【Conclusion】 In summary, MeJA fumigation of navel orange fruits for 24 h before inoculation could significantly enhance the resistance of fruits to P. italicum, increase the activity of enzymes related to active oxygen metabolism and the expression levels of key enzyme genes. The above results suggested that the enhancement of the resistance to the blue mold in navel orange fruits induced by the MeJA treatment might be related to its regulation of the reactive oxygen species metabolism. This study could provide theoretical basis and technical reference for the prevention and control of the blue mold disease during postharvest storage of the navel orange fruits.
Key words: Navel orange fruit; Methyl jasmonate; Resistance to blue mold; Reactive oxygen metabolism
在柑橘的贮运过程中,果实采后腐烂严重,烂果率为10%~30%,其中由意大利青霉(Penicillium italicum)侵染引起的青霉病是果实腐烂的主要致病原因之一[1],目前主要通过化学药剂控制该病害。由于使用防治的化学药剂在一定程度上也会抑制柑橘果实的生理活性,且长期使用化学试剂会降低植物对病原菌的抵御能力,使防治效果大大降低,以及公众对食品健康和环境污染的日益关注,因此,开发出更多绿色安全高效的方法来控制柑橘果实采后病害具有重要的生产意义。活性氧(ROS)的积累是由于植物遭受生物或非生物胁迫时,抗氧化防御系统不能及时消除其体内过量的ROS而产生的氧化应激反应;其后果是破坏植物代谢平衡,导致植物生理紊乱甚至衰老死亡[2-3]。
茉莉酸甲酯(methyl jasmonate,MeJA)是一种重要的天然植物生长调节剂,可以作为植物抵抗病原体侵染产生防御反应的诱抗剂[4]。王瀚博[5]在研究MeJA预处理采后蓝莓果实对灰霉病抗性的影响中发现,MeJA可通过增强超氧化物歧化酶(SOD)、过氧化氢酶(CAT)和抗坏血酸过氧化物酶(APX)等抗氧化酶活性,减轻活性氧损伤进而增强采后蓝莓果实对灰霉病的抗性。Wang等[6]的研究发现,MeJA能诱导杨梅抗氧化能力的增强,提高果实对橘青霉病菌(P. citrinum)的抗性。在外源MeJA诱导猕猴桃抗软腐病的研究中发现,MeJA诱导了果实中过氧化物酶(POD)、CAT和SOD等抗氧化酶活性的提高,同时也有效提高了相关酶基因AcPOD、AcSOD的表达量[7]。Cao等[8]的研究表明,10 μmol·L-1 MeJA处理采后枇杷果实可通过增加多胺的含量来抑制炭疽病的发生。王建伟等[9]研究表明外源施加MeJA后能够减轻盐胁迫对菜用甘薯光合系统的伤害,并且提高抗氧化酶活性和渗透调节物质的含量,促进菜用甘薯有机物质的积累,从而提高产量。以上研究表明,外源MeJA的应用能够诱导采后果蔬产生防御反应,激活相关防御酶活性,促进抗病物质积累,从而有效增强其对病原菌的抗性[4]。目前有关MeJA诱导脐橙果实抗青霉病的报道较少。笔者课题组在前期的试验中表明MeJA在调控脐橙果实抗青霉病中发挥重要作用,与其提高果实中防御酶POD、多酚氧化酶(PPO)以及病程相关蛋白几丁质酶(CHI)、β-1,3-葡聚糖酶(GLU)活性密切相关,但MeJA诱导抗病与脐橙果实活性氧代谢之间的关系仍需要进一步探究。本试验中,以纽荷尔和龙回红脐橙果实为试材,探究外源MeJA熏蒸处理对采后脐橙果实活性氧代谢相关酶活性及其酶基因表达的影响,旨在进一步探究MeJA调控脐橙果实抗采后青霉病的作用机制,为MeJA应用于果实贮藏保鲜提供理论依据。
1 材料和方法
1.1 试验材料
试验用果:纽荷尔、龙回红脐橙果实,于2021年11月12日采自江西省赣州市南康县龙回镇俊萍果业示范园(25.66° N,114.76° E),翌日运回实验室。选出大小均匀、成熟度一致、无病虫害和机械损伤的果实作为试验用果。先用自来水清洗,再用0.05%次氯酸钠溶液浸洗2 min,最后使用自来水冲洗干净。室温下晾干后使用保鲜袋单果套袋,入库备用(冷库温度:7~8 ℃)。
供试青霉病菌(P. italicum):由果蔬贮藏与保鲜江西省重点实验室提供。从典型脐橙发病果实中单孢分离,用马铃薯葡萄糖琼脂培养基(PDA)进行纯化培养。试验前于恒温箱[(25±1) ℃]培养5~7 d,用无菌水洗脱孢子,经灭菌脱脂棉过滤后,配置浓度为1.25×106 spores·mL-1孢子悬浮液,备用。
试验试剂:茉莉酸甲酯(MeJA):纯度95%,水杨苷异羟肟酸(salicyhydroxamic acid,SHAM,茉莉酸生物合成抑制剂)购自美国Sigma-Aldrich公司,配制浓度为100 μmol·L-1,待用。
1.2 脐橙果实处理
预试验筛选出最适浓度50 μmol·L-1和最适熏蒸时间24 h的MeJA处理,在(26±1) ℃、相对湿度85%~95%环境下密闭熏蒸处理纽荷尔和龙回红脐橙果实;以无菌水处理为对照,另设100 μmol·L-1 SHAM喷施处理,共3组处理,24 h后于超净工作台上通风1 h后接种。脐橙果实表面经75%乙醇消毒后,用接种针在其赤道部等距离刺伤,伤口大小为直径2 mm,深度3 mm;注入浓度为1.25×106 spores·mL-1的P. italicum孢子悬浮液20 µL。每组处理3次重复,每次重复30个果实,每处理共90个果实,处理后置于温度(26±1) ℃、相对湿度85%~95%培养箱。接种后0~7 d逐日从各处理中随机选取10个果实,取3组处理脐橙果实病健交界处1 cm果皮组织,用液氮迅速冷冻研磨装袋,于-80 ℃超低温保存。
1.3 活性氧代谢相关酶活性和物质含量的测定
谷胱甘肽还原酶(GR)、APX活性,还原型谷胱甘肽(GSH)、抗坏血酸(AsA)含量和超氧阴离子(O2.-)产生速率和丙二醛(MDA)含量参照曹建康等[10]方法测定,SOD活性、过氧化氢(H2O2)含量采用试剂盒(南京建成生物技术有限公司,南京,中国)测定。CAT活性参考李合生[11]的方法测定。DPPH自由基清除率参考张昭等[12]的方法测定。单脱氢抗坏血酸还原酶(MDHAR)和脱氢抗坏血酸还原酶(DHAR)活性参考Wei等[13]的方法测定。
1.4 活性氧代谢途径关键基因表达分析
参考马巧利[14]的方法,采用改良Trizol法提取果皮总RNA,使用微量核酸分析仪和1%琼脂糖凝胶电泳检测RNA质量。使用Hifair®Ⅲ试剂盒(翌圣,上海)反转录RNA合成cDNA链。制备的cDNA保存-80 ℃超低温冰箱用于后续qRT-PCR试验。基因表达测定:以甜橙CsActin基因为定量的内参基因,通过qRT-PCR方法检测6种活性氧代谢抗病相关基因的表达量。引物序列如表1所示。PCR反应程序为:95 ℃预变性30 s,95 ℃持续5 s,60 ℃退火持续30 s,72 ℃延伸30 s,共反应40个循环。反应体系为10 μL,包含1 μL cDNA + 0.3 μL F primer (10 mmol·L-1)+0.3 μL R primer (10 mmol·L-1) + 5 μL TB Green® Premix Ex Taq™ + 3.4 μL ddH2O。每个样品3次重复测定,基因表达水平计算采用2-ΔΔCt方法[15]。
1.5 数据处理与分析
试验按照完全随机试验设计,全部试验指标设3次生物学重复。数据采用Excel 2018和SPSS 20.0软件进行统计整理和单因素方差分析(ANOVA)。采用Origin 2018软件作图,图中所有数据以平均值(mean)±标准误差(SE)表示。
2 结果与分析
2.1 MeJA处理对脐橙果实GR活性和GSH含量的影响
从图1-A可知,除接种后第1天和第6天外,纽荷尔MeJA处理组GR活性在接种后7 d内均显著高于对照(CK)和SHAM处理组(p<0.05),其中在接种后第5天到达峰值,分别是对照和SHAM处理组的1.10倍和1.13倍。从图1-B可知,龙回红MeJA处理组GR活性在除接种后第4天和第6天外均显著高于CK和SHAM处理组(p<0.05);MeJA处理组的GR活性在接种后第5天最高,分别是CK和SHAM处理组的1.13和1.16倍。
在接种后第3天和第5~6天,纽荷尔MeJA处理组GSH含量显著高于CK和SHAM处理组(p<0.05);且在接种后第6天最高,分别是CK和SHAM处理组的1.17和1.32倍(图1-C)。在接种后第1、3天和第6~7天,龙回红MeJA处理组GSH含量显著高于CK和SHAM处理组(p<0.05);且在接种后第7天最高,分别是CK和SHAM处理组的1.17和1.20倍(图1-D)。
2.2 MeJA处理对脐橙果实APX活性和AsA含量的影响
纽荷尔脐橙MeJA和CK处理组APX活性呈先上升后下降、然后再上升最后又下降的趋势,在接种后第3天最高,MeJA处理组显著高于CK和SHAM处理组(p<0.05),分别是CK和SHAM处理组的1.16、1.22倍(图2-A);龙回红MeJA和CK处理组APX活性总体呈先上升后下降趋势,CK和处理组APX活性均在接种后第4天最高,MeJA处理组显著高于CK和SHAM处理组(p<0.05),分别是CK和SHAM处理组的1.49、1.45倍(图2-B)。随着接种时间的延长,纽荷尔MeJA和SHAM处理组均呈双峰趋势。在接种后除第4天外,纽荷尔MeJA处理组AsA含量均显著高于CK和SHAM处理组(p<0.05)(图2-C);MeJA处理组AsA含量在接种后第6天最高,分别是CK和SHAM处理组的1.12和1.16倍。于接种后第1~7天,龙回红3组处理均大致呈先上升后下降趋势。在接种后第1天和第3~6天,龙回红MeJA处理组AsA含量显著高于CK和SHAM处理组(p<0.05)(图2-D);MeJA处理组AsA含量在接种后第4天最高,为CK和SHAM处理组的1.20和1.14倍。
2.3 MeJA处理对脐橙果实SOD和CAT活性的影响
由图3-A可知,纽荷尔3组处理SOD活性出现双峰趋势。在除接种后第6天外,MeJA处理SOD活性均显著高于CK组(p<0.05);且在接种后第5天最高,分别是CK和SHAM处理组的1.12和1.27倍。如图3-B可知,龙回红MeJA处理SOD活性在接种后第3、7天显著高于CK组(p<0.05);且在接种后第6天最高,分别是CK和SHAM处理组的1.02和1.10倍。
纽荷尔3组处理呈先上升后下降趋势(图3-C)。在接种后第1、6、7天,MeJA处理CAT活性显著高于CK和SHAM处理组(p<0.05);在接种后第6天最高,分别是CK和SHAM处理组的1.69和2.10倍。龙回红MeJA处理组CAT活性在接种后除第6天外均显著高于CK和SHAM处理组(p<0.05);MeJA处理组CAT活性于接种后第4天最高,分别是CK和SHAM处理组的1.41和1.96倍(图3-D)。
2.4 MeJA处理对脐橙果实H2O2含量和O2.-产生速率的影响
如图4-A所示,纽荷尔3组处理H2O2含量呈先上升后下降趋势,于接种后第6天最高。MeJA处理H2O2含量在接种后第1~4天和第7天显著低于CK和SHAM处理组(p<0.05);且在接种后第4天,仅为CK和SHAM处理组的51.21%和44.37%。如图4-B所示,龙回红3组处理H2O2含量大致呈上升趋势。在接种后第3~5天,MeJA处理H2O2含量显著低于CK组(p<0.05);于接种后第4天,分别为CK和SHAM处理组的84.67%和90.93%。
从图4-C和图4-D可知,MeJA和SHAM处理组O2.-产生速率呈双峰趋势。在除接种后第5天外,纽荷尔MeJA处理O2.-产生速率均显著低于CK组(p<0.05);在接种后第3天最低,仅为CK和SHAM处理组的24.56%和31.84%。龙回红MeJA处理O2.-产生速率在接种后第1天和第4~7天显著低于CK组(p<0.05);于接种后第3天最低,分别为CK和SHAM处理组的95.33%和66.56%。
2.5 MeJA处理对脐橙果实MDA含量和DPPH自由基清除率的影响
如图5-A和图5-B所示,纽荷尔和龙回红3组处理MDA含量均随接种时间的延长呈大致上升趋势。纽荷尔MeJA处理在接种后除第5天外显著低于CK组(p<0.05);在接种后第3天最低,分别为CK和SHAM处理组的64.08%和67.46%。龙回红MeJA处理组MDA含量在接种后第2~5天显著低于CK组(p<0.05);于接种后第5天,含量是CK和SHAM处理组的74.93%和71.89%。
在接种后第3~7天,纽荷尔MeJA处理DPPH自由基清除率显著高于CK组(p<0.05);在接种后第6天,MeJA处理DPPH自由基清除率最高,分别是CK和SHAM处理组的1.29和1.19倍(图5-C)。在除接种后第2、5天外,龙回红MeJA处理DPPH自由基清除率均显著高于CK组(p<0.05);于接种后第7天,MeJA处理DPPH自由基清除率最高,分别是CK和SHAM处理组的1.18和1.11倍(图5-D)。
2.6 MeJA处理对脐橙果实MDHAR和DHAR活性的影响
由图6-A可知,纽荷尔MeJA处理MDHAR活性在接种后第3~7天显著高于CK和SHAM处理组(p<0.05);于接种后第3天达到峰值,MeJA处理MDHAR活性分别是CK和SHAM处理组的1.21和1.78倍。由图6-B可知,除在接种后第6天龙回红MeJA处理MDHAR活性与SHAM处理无显著差异外,在接种后第2~7天显著高于CK和SHAM处理组(p<0.05);于接种后第5天最高,MDHAR活性分别是CK和SHAM处理组的1.26和1.53倍。
随着接种时间的延长,纽荷尔MeJA和SHAM处理组DHAR活性出现双峰趋势(图6-C)。于接种后第1天和第4~6天,MeJA处理DHAR活性显著高于CK和SHAM处理组(p<0.05);在接种后第5天最高,分别是CK和SHAM处理组的1.43和1.47倍。从图6-D可知,龙回红3组处理DHAR活性均大致呈先上升后下降的趋势。于接种后第4天,MeJA处理DHAR活性最高,分别是CK和SHAM处理组的1.79和1.91倍。
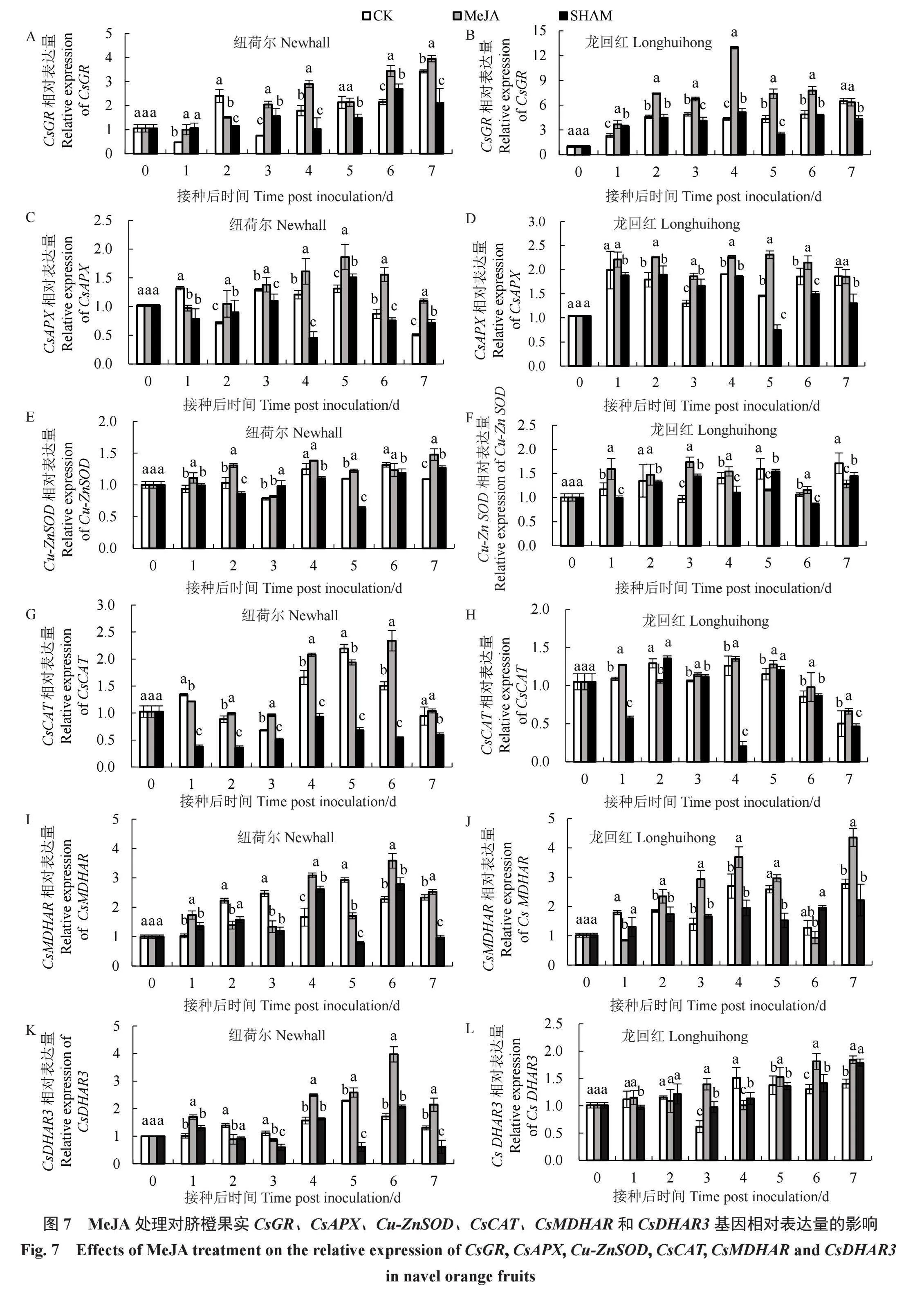
2.7 MeJA处理对脐橙果实活性氧代谢途径关键酶基因表达的影响
由图7-A所示,于接种后第3~4天和第6~7天,纽荷尔MeJA处理组CsGR相对表达量显著高于CK和SHAM处理组(p<0.05);在接种后第7天,MeJA处理组CsGR相对表达量最高,分别是CK和SHAM处理组的1.16和1.86倍。由图7-B所示,除接种后第7天外,龙回红MeJA处理组CsGR相对表达量显著高于CK和SHAM处理组(p<0.05);于接种后第4天,MeJA处理组CsGR相对表达量最高,分别是CK和SHAM处理组的2.98和2.52倍。
纽荷尔MeJA处理组CsAPX相对表达量除接种后第1天外均显著高于CK和SHAM处理组(p<0.05)(图7-C);MeJA处理组CsAPX相对表达量在接种后第5天最高,分别是CK和SHAM处理组的1.42和1.24倍。龙回红MeJA处理组CsAPX相对表达量在接种后第2~6天显著高于CK和SHAM处理组(p<0.05)(图7-D);MeJA处理组CsAPX相对表达量于接种后第5天最高,分别是CK和SHAM处理组的1.59和3.01倍。
如图7-E所示,在接种后除第3、4和6天外,纽荷尔MeJA处理组Cu-ZnSOD相对表达量显著高于CK和SHAM处理组(p<0.05);于接种后第7天,MeJA处理组Cu-ZnSOD相对表达量最高,分别是CK和SHAM处理组的1.36和1.17倍。如图7-F所示,在接种后第3天,龙回红MeJA处理组Cu-ZnSOD相对表达量达到峰值,分别是CK和SHAM处理组的1.79和1.20倍。
在接种后第2~4天和第6天,纽荷尔MeJA处理组CsCAT相对表达量显著高于CK和SHAM处理组(p<0.05);MeJA处理组CsCAT相对表达量于接种后第6天最高,分别是CK和SHAM处理组的1.56和4.41倍(图7-G)。在接种后除第2天外,龙回红MeJA处理组CsCAT相对表达量均显著高于CK和SHAM处理组(p<0.05);在接种后第4天,MeJA处理组CsCAT相对表达量最高,分别是CK和SHAM处理组的1.10和6.75倍(图7-H)。
从图7-I可知,纽荷尔MeJA处理组CsMDHAR相对表达量在接种后第1、4天和第6~7天显著高于CK和SHAM处理组(p<0.05);其中在接种后第6天,MeJA处理组CsMDHAR相对表达量最高,分别是CK和SHAM处理组的1.58和1.29倍。从图7-J可知,除接种后第1、6天外,龙回红MeJA处理组CsMDHAR相对表达量均显著高于CK和SHAM处理组(p<0.05);MeJA处理组CsMDHAR相对表达量在接种后第7天最高,分别是CK和SHAM处理组的1.57和1.96倍。
由图7-K所示,在接种后除第2、3天外,纽荷尔MeJA处理组CsDHAR3相对表达量显著高于CK和SHAM处理组(p<0.05);于接种后第6天,MeJA处理组CsDHAR3相对表达量最高,分别是CK和SHAM处理组的2.32和1.91倍。由图7-L所示,在接种后第3天和第5~6天,龙回红MeJA处理组CsDHAR3相对表达量显著高于CK和SHAM处理组(p<0.05);MeJA处理组CsDHAR3相对表达量在接种后第7天达到峰值,分别是CK和SHAM处理组的1.31和1.03倍。
3 讨 论
MeJA作为一种抗病信号分子,可以诱导提高多种采后果蔬对病原菌侵染的抵抗能力。前人研究表明使用MeJA处理提高了果实抗氧化酶CAT、APX和SOD活性且促进了GSH、AsA含量增加,并降低了O2.-产生速率、抑制H2O2含量积累[16-18]。笔者在本试验结果中显示,与对照组相比,50 μmol·L-1 MeJA熏蒸处理纽荷尔和龙回红脐橙果实,均显著提高了GR、APX、SOD和CAT活性,降低了H2O2含量和O2.-产生速率,并促进了GSH和AsA积累,表明外源MeJA处理激发了活性氧代谢途径,显著增强了SOD活性,将不稳定的O2.-分解成为更稳定的H2O2;同时H2O2又在CAT和APX的作用下被分解为H2O和O2,以及在GR的催化下GSSG被还原形成GSH。在MeJA处理下,脐橙果实体内活性氧过度积累被清除,恢复了活性氧动态平衡,从而减轻了果实采后青霉病的发生。可见在外源激发因子的诱导下果蔬抗氧化酶活性增强同时抗氧化合物增加,清除了过度积累的活性氧,从而增强了果蔬抵抗病原菌侵染的能力。MDA是膜脂质过氧化的重要产物,其含量可以直接反映对植物细胞膜的破坏程度,是寄主抗病能力的重要指标[19]。DPPH自由基清除率是植物组织内的自由基清除率的判定指标。ROS清除系统中包含AsA-GSH循环,GR、APX、MDHAR和DHAR是AsA-GSH循环中的关键酶。APX催化AsA产生MDHA,并消除H2O2;MDHA在MDHAR催化下可形成AsA,GSH为DHAR提供电子将DHA催化为AsA。笔者在本研究中发现,与CK组和SHAM组比较,MeJA熏蒸处理影响两品种脐橙果实活性氧代谢,果实内MDHAR和DHAR活性得到显著提高,DPPH自由基清除率显著提升,进而有效延缓了MDA的积累。试验结果与前人用MeJA与氯化钠联合处理提高了玉米芽中抗氧化酶活性和清除自由基能力的结论相一致[20]。同时,为应对冷害、盐胁迫和病害等生物或非生物胁迫,前人在研究MeJA处理对番茄[21]、玉米[22]和枇杷[23]的影响中发现,MeJA能够不同程度地提高抗氧化物质GSH、AsA的含量和AsA-GSH循环中关键酶MDHAR、DHAR的活性,进而诱导果蔬增强对外界环境的抗性。由此推测MeJA处理诱导脐橙果实对青霉病的抗性与其调控活性氧代谢相关酶活性及物质含量有关,且笔者前期研究表明,龙回红脐橙果实病斑直径在接种第3天显著小于纽荷尔脐橙果实病斑直径,说明龙回红比纽荷尔脐橙果实抗青霉病,可能导致了MeJA处理调控活性氧代谢相关酶活性及物质含量在脐橙不同品种间存在差异,但其原因还需要进一步深入研究。
植物在遭受病原微生物侵染时,不同外源诱导剂可调控植物次生代谢,激活多种相关防御基因的表达,诱导植物产生防御反应。前人研究表明外源MeJA处理后均能增加果实体内活性氧代谢关键酶基因的表达量从而增强果实对病原菌的抗性[24-26]。此外,其他诱抗剂如苯并噻重氮(acibenzolar-S-methyl,ASM)和褪黑素(melatonin,MT)在对梨果实抗采后黑斑病的研究中指出,ASM处理激活了果实中抗病相关基因PcSOD、PcCAT、PcAPX和PcDHAR显著表达;以及MT预处理显著提高了果实中防御酶基因PpCAT、PpSOD、PpCHI和PpGLU的表达且在后期维持高水平表达,进而有效提高梨果实抗采后黑斑病的能力[27-28]。本试验结果表明,于接种前用MeJA熏蒸处理脐橙果实后,能诱导脐橙果实活性氧代谢相关防御酶基因(CsGR、CsAPX、Cu-ZnSOD、CsCAT、CsMDHAR和CsDHAR3)不同程度地显著表达。由此推测MeJA诱导采后脐橙对青霉病的抗性可能与其活性氧代谢中相关防御酶基因的高表达密切相关。
4 结 论
在接种P. italicum前采用外源50 μmol·L-1 MeJA熏蒸处理纽荷尔和龙回红脐橙果实24 h,显著提高了活性氧代谢相关酶(GR、APX、SOD、CAT、MDHAR和DHAR)活性,促进了AsA、GSH含量的增加,降低了O2.-产生速率和H2O2含量、抑制了MDA的积累并提高了果实内DPPH自由清除率水平;同时激活了相关防御酶基因(CsGR、CsAPX、Cu-ZnSOD、CsCAT、CsMDHAR和CsDHAR3)的显著表达,进而提升了脐橙果实抵抗青霉病菌侵染的能力。
参考文献References:
[1] 解淑慧,邵兴锋,王可,张兴龙,王鸿飞. 柑橘采后腐烂主要致病菌的分离鉴定及丁香精油对其抑制作用研究[J]. 果树学报,2013,30(1):134-139.
XIE Shuhui,SHAO Xingfeng,WANG Ke,ZHANG Xinglong,WANG Hongfei. Isolation and identification of dominant pathogen on citrus fruit and the antifungal effect of clove oil on these fungi[J]. Journal of Fruit Science,2013,30(1):134-139.
[2] HASANUZZAMAN M,NAHAR K,HOSSAIN M S,MAHMUD J A,RAHMAN A,INAFUKU M,OKU H,FUJITA M. Coordinated actions of glyoxalase and antioxidant defense systems in conferring abiotic stress tolerance in plants[J]. International Journal of Molecular Sciences,2017,18(1):200.
[3] TORRES M A. ROS in biotic interactions[J]. Physiologia Plantarum,2010,138(4):414-429.
[4] ASGHARI M. Impact of jasmonates on safety,productivity and physiology of food crops[J]. Trends in Food Science amp; Technology,2019,91:169-183.
[5] 王瀚博. 茉莉酸甲酯对蓝莓果实采后品质及抗病性调控机制研究[D]. 南京:南京林业大学,2021.
WANG Hanbo. The regulatory mechanism of physiology,biochemistry and disease resistance mediated by methyl jasmonate in postharvest blueberries[D]. Nanjing:Nanjing Forestry University,2021.
[6] WANG K T,JIN P,HAN L,SHANG H T,TANG S S,RUI H J,DUAN Y F,KONG F Y,KAI X,ZHENG Y H. Methyl jasmonate induces resistance against Penicillium citrinum in Chinese bayberry by priming of defense responses[J]. Postharvest Biology and Technology,2014,98:90-97.
[7] PAN L Y,ZHAO X Y,CHEN M,FU Y Q,XIANG M L,CHEN J Y. Effect of exogenous methyl jasmonate treatment on disease resistance of postharvest kiwifruit[J]. Food Chemistry,2020,305:125483.
[8] CAO S F,CAI Y T,YANG Z F,JOYCE D C,ZHENG Y H. Effect of MeJA treatment on polyamine,energy status and anthracnose rot of loquat fruit[J]. Food Chemistry,2014,145:86-89.
[9] 王建伟,许光龄,陈艳丽,朱国鹏,陈玥,刘金伟. 外源茉莉酸甲酯对盐胁迫下菜用甘薯生长生理的影响[J]. 中国瓜菜,2022,35(6):69-75.
WANG Jianwei,XU Guangling,CHEN Yanli,ZHU Guopeng,CHEN Yue,LIU Jinwei. Exogenous methyl jasmonate affects growth and physiology of leafy sweet potato under salt stress[J]. China Cucurbits and Vegetables,2022,35(6):69-75.
[10] 曹建康,姜微波,赵玉梅. 果蔬采后生理生化实验指导[M]. 北京:中国轻工业出版社,2007.
CAO Jiankang,JIANG Weibo,ZHAO Yumei. Experimental guidance of postharvest physiology and biochemistry of fruits and vegetables[M]. Beijing:China Light Industry Press,2007.
[11] 李合生. 植物生理生化实验原理和技术[M]. 北京:高等教育出版社,2000.
LI Hesheng. Principles and techniques of plant physiological biochemical experiment[M]. Beijing:Higher Education Press,2000.
[12] 张昭,吕泽芳,吴洪梅,周志钦,于杰. 重庆市10大名柚果实酚类物质质量分数及抗氧化活性研究[J]. 西南大学学报(自然科学版),2015,37(5):58-65.
ZHANG Zhao,LÜ Zefang,WU Hongmei,ZHOU Zhiqin,YU Jie. Phenolic contents of ten famous Chongqing pummelo (Citrus grandis) cultivars and their antioxidant capacities[J]. Journal of Southwest University (Natural Science Edition),2015,37(5):58-65.
[13] WEI M L,GE Y H,LI C Y,HAN X,QIN S C,CHEN Y R,TANG Q,LI J R. G6PDH regulated NADPH production and reactive oxygen species metabolism to enhance disease resistance against blue mold in apple fruit by acibenzolar-S-methyl[J]. Postharvest Biology and Technology,2019,148:228-235.
[14] 马巧利. 2,4-二氯苯氧乙酸保鲜柑橘果实采后品质机理[D]. 武汉:华中农业大学,2014.
MA Qiaoli. Mechanism underlying 2,4-dichlorophenoxyacetic acid maintaining citrus fruit postharvest quality[D]. Wuhan:Huazhong Agricultural University,2014.
[15] LIVAK K J,SCHMITTGEN T D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCt Method[J]. Methods,2001,25(4):402-408.
[16] HABIBI F,RAMEZANIAN A,GUILLÉN F,SERRANO M,VALERO D. Blood oranges maintain bioactive compounds and nutritional quality by postharvest treatments with γ-aminobutyric acid,methyl jasmonate or methyl salicylate during cold storage[J]. Food Chemistry,2020,306:125634.
[17] DONG Y,ZHI H H,XU J,ZHANG L H,LIU M P,ZONG W. Effect of methyl jasmonate on reactive oxygen species,antioxidant systems,and microstructure of Chinese winter jujube at two major ripening stages during shelf life[J]. Journal of Horticultural Science and Biotechnology,2016,91(3):316-323.
[18] 王英珍,程瑞,张绍铃,白彬,何子顺,张虎平. 采前茉莉酸甲酯(MeJA)处理对梨果实抗病性的影响[J]. 果树学报,2016,33(6):694-700.
WANG Yingzhen,CHENG Rui,ZHANG Shaoling,BAI Bin,HE Zishun,ZHANG Huping. Effect of pre-harvest methyl jasmonate treatment on disease resistance in pear fruit[J]. Journal of Fruit Science,2016,33(6):694-700.
[19] DIETZ K J,MITTLER R,NOCTOR G. Recent progress in understanding the role of reactive oxygen species in plant cell signaling[J]. Plant Physiology,2016,171(3):1535-1539.
[20] HE W W,LUO H,XU H,ZHOU Z Y,LI D J,BAO Y H,FU Q,SONG J F,JIAO Y,ZHANG Z Y. Effect of exogenous methyl jasmonate on physiological and carotenoid composition of yellow maize sprouts under NaCl stress[J]. Food Chemistry,2021,361:130177.
[21] 李子龙. SlMYC2调控茉莉酸甲酯介导AsA-GSH循环提高番茄果实抗冷性的机制研究[D]. 淄博:山东理工大学,2022.
LI Zilong. The roles of SlMYC2 in regulating ascorbate-glutathione cycle mediated by methyl jasmonate in postharvest tomato fruits under cold stress[D]. Zibo:Shandong University of Technology,2022.
[22] 陈芳,杨双龙,张莉,佟祎鑫,张志华,虞凡枫,贾凌云. 外源茉莉酸甲酯对盐胁迫下玉米幼苗AsA-GSH循环的影响[J]. 生物学通报,2021,56(11):44-48.
CHEN Fang,YANG Shuanglong,ZHANG Li,TONG Yixin,ZHANG Zhihua,YU Fanfeng,JIA Lingyun. Effects of exogenous methyl jasmonate on ascorbate-glutathione cycle in Zea mays seedlings under salt stress[J]. Bulletin of Biology,2021,56(11):44-48.
[23] 蔡玉婷. MeJA处理减轻枇杷采后冷害和病害的机理研究[D]. 南京:南京农业大学,2012.
CAI Yuting. Study on mechanism of MeJA treatment on chilling injury and disease of loquat after harvest[D]. Nanjing:Nanjing Agricultural University,2012.
[24] PAN L Y,CHEN X R,XU W,FAN S S,WAN T,ZHANG J,CAI Y L. Methyl jasmonate induces postharvest disease resistance to decay caused by Alternaria alternata in sweet cherry fruit[J]. Scientia Horticulturae,2022,292:110624.
[25] SAAVEDRA G M,SANFUENTES E,FIGUEROA P M,FIGUEROA C R. Independent preharvest applications of methyl jasmonate and chitosan elicit differential upregulation of defense-related genes with reduced incidence of gray mold decay during postharvest storage of Fragaria chiloensis fruit[J]. International Journal of Molecular Sciences,2017,18(7):1420.
[26] 赵显阳. 外源茉莉酸甲酯(MeJA)对梨果实抗青霉病及其保鲜作用的研究[D]. 南昌:江西农业大学,2020.
ZHAO Xianyang. Effects of exogenous methyl jasmonate (MeJA) induced resistance to blue mold and storage preservation in postharvest pear fruit[D]. Nanchang:Jiangxi Agricultural University,2020.
[27] LI X,LI C Y,CHENG Y,HOU J B,ZHANG J H,GE Y H. Postharvest application of acibenzolar- S-methyl delays the senescence of pear fruit by regulating reactive oxygen species and fatty acid metabolism[J]. Journal of Agricultural and Food Chemistry,2020,68(17):4991-4999.
[28] 向妙莲,吴帆,李树成,马巧利,王印宝,肖刘华,陈金印,陈明. 外源褪黑素调控活性氧代谢诱导梨果实抗采后黑斑病[J]. 园艺学报,2022,49(5):1102-1110.
XIANG Miaolian,WU Fan,LI Shucheng,MA Qiaoli,WANG Yinbao,XIAO Liuhua,CHEN Jinyin,CHEN Ming. Exogenous melatonin regulates reactive oxygen metabolism to induce resistance of postharvest pear fruit to black spot[J]. Acta Horticulturae Sinica,2022,49(5):1102-1110.
