
不同有机替代处理对旱地麦田土壤微生物群落的影响
2024-12-31 00:00:00乔青青刘治国姜丽伟赵晓东李廷亮谢英荷
天津农业科学 2024年10期
摘" " 要:为探明不同有机替代处理对土壤功能微生物和参与氮循环功能酶基因的影响,在山西洪洞旱地麦田长期定位试验基地设定有机肥(OF)和腐殖酸(HF)2种替代处理,并对其进行研究。结果表明,不同有机替代处理对土壤真菌群落多样性和结构的改变明显高于细菌群落。Network分析发现,不同有机替代处理均增加了微生物的共现性网络边数、节点数和平均邻居数,并且细菌群落间互相作用更复杂。Streptomyces、Paenibacillus、Chaetomium和Talaromyces为植物病原菌的生物防治菌,OF处理的Streptomyces、Paenibacillus、Chaetomium和Talaromyces丰度较HF处理均有不同程度提高。植物病原菌Fusarium和Acremonium丰度在OF处理中为最低值,较HF处理分别降低33%和37%。11个参与氮循环的酶基因被发现,OF处理的总丰度较HF处理增加47%,特别是OF处理的酶基因EC 1.7.2.6和EC 1.7.99.-丰度较HF处理分别提升126%和95%,这将有利于氮素养分的供应。此外,潜在解磷菌的丰度在不同有机替代处理中呈增加趋势。综上所述,有机肥替代处理较腐殖酸替代处理更能为晋南旱地小麦生长提供优良的条件,适宜在该区域推广。
关键词:小麦;有机替代;微生物;氮循环;功能酶基因
中图分类号:S154.3" " " " " 文献标识码:A" " " " " DOI 编码:10.3969/j.issn.1006-6500.2024.10.011
Effects of Different Organic Substitutes on Soil Microbial Community in Dryland Wheat Fields
QIAO Qingqing1,2, LIU Zhiguo3, JIANG Liwei4, ZHAO Xiaodong1,2, LI Tingliang4, XIE Yinghe4
(1. College of Biological Sciences and Technology, Taiyuan Normal University, Yuci, Shanxi 030619, China; 2. Shanxi Key Laboratory of Earth Surface Processes and Resource Ecology Security in Fenhe River Basin, Taiyuan Normal University, Yuci, Shanxi 030619, China; 3. Agricultural Industry Development Center of Pianguan County, Pianguan, Shanxi 036400, China; 4. College of Resources and Environment, Shanxi Agricultural University, Taigu, Shanxi 030801, China)
Abstract: In order to investigate the effects of different organic substitutes on soil functional microorganisms and functional enzyme genes involved in nitrogen cycling, organic fertilizer (OF) and humic acid (HF) were set up in a long-term location experiment based in the dryland wheat field of Hongtong, Shanxi Province. The results revealed that the changes in the diversity and structure of soil fungal communities were notably more pronounced than those in bacterial communities resulting from various organic substitutes. Network analysis showed that different organic substitutes increased the number of edges, nodes, and average neighbors in the microbial co-occurrence network, making the interactions between bacterial communities more complex. The abundance of biological control agents for plant pathogens, such as Streptomyces, Paenibacillus, Chaetomium, and Talaromyces, was higher in OF compared to the HF. The abundance of plant pathogenic fungi, Fusarium and Acremonium, showed the lowest abundance in the OF, which was 33% and 37% lower than that in the HF. A total of 11 enzyme genes involved in the nitrogen cycle showed a 47% increase in abundance in the OF compared to the HF. In particular, the abundance of enzyme genes EC 1.7.2.6 and EC 1.7.99.- increased by 126% and 95% in the OF compared to the HF, potentially enhancing nitrogen nutrient availability. In addition, the abundance of potential phosphate-solubilizing microbes also showed an increasing trend in different organic substitute treatments. In conclusion, organic fertilizer substitute provides more favorable conditions for dryland wheat growth in southern Shanxi compared to humic acid substitution, making it suitable for regional promotion.
Key words: wheat; organic substitute; microorganism; nitrogen cycling; functional enzyme gene
中国是世界上较早种植小麦的国家之一,也是世界上最大的小麦生产国,其小麦产量约占全球总产量的17%[1],自给率已超过100%[2]。然而,过度依赖或单一使用化肥会导致土壤酸化和肥力下降,降低土壤微生物的多样性[3],进而威胁着粮食产量的提升,这明显有悖于绿色发展以及可持续发展理念。因此,有机替代正逐步成为一种可持续的农业实践方式。多项研究表明,有机肥与化肥配合施用可以增加土壤有机质和养分含量,改善土壤肥力和结构,从而保证农业的可持续性和作物产量[4-6]。Zhou等[5]研究发现,有机氮肥和无机氮肥按1∶1和2∶1的比例混合施用可显著提高蔬菜产量和氮素吸收率。Ren等[6]研究表明,有机肥代替化肥可提高作物产量。同时,腐殖酸通过调节土壤微生物群落,改善土壤结构,增强土壤肥力,促进作物生长[7-9],提高小麦对养分的吸收与利用,进一步提升小麦产量[10]。前人研究表明,相较于同等养分配方肥,添加腐殖酸的复合肥可促进作物生长,改善土壤健康和肥力[11]。因此,为进一步提升土壤质量和作物产量,本研究选择有机肥和腐殖酸2种有机替代方式,并对其进行深入研究,为推动可持续农业实践提供科学依据。
氮元素作为作物生长繁殖的重要养分之一,既是合成含氮代谢产物的源泉,也是保证土壤肥效和质量的关键因素[12]。Liu等[13]研究发现,与单独使用化肥相比,牛粪与化肥的混合施用显著增加了土壤氮含量,改善了土壤细菌和真菌群落。王伟华等[14]研究显示,化肥配合秸秆还田或有机肥配施显著改善了土壤养分含量及微生物活性。先前研究发现,有机肥的添加能够将寡营养型微生物(在纯矿物肥料或不施肥土壤中茁壮成长的微生物)转变为能够分解复杂有机化合物的微生物,引起参与养分循环的关键物种变化,从而影响微生物群落,增强土壤肥力[15]。笔者先前的研究表明,有机替代方式对可溶性有机氮、微生物有机氮和轻组有机氮的提高均有促进作用,并且有机肥替代部分化肥较腐殖酸替代部分化肥更有利于作物产量的提高[16]。相比于腐殖酸替代方式,有机肥替代对方式土壤微生物群落结构及其对山西晋南旱地小麦生长的影响还有待进一步研究。
本研究选取农民常用的有机肥和腐殖酸肥料,基于本课题组先前的研究结果,本研究设置有机肥替代处理和腐殖酸替代处理,研究功能微生物及参与氮循环的功能基因在不同有机替代处理中的响应特征,以期为土壤可持续发展、科学施肥和作物增产提供科学依据。
1 材料与方法
1.1 试验地概况
试验地点位于山西省临汾市洪洞县刘家垣镇东梁村旱地冬麦区试验基地(111°35′ E,36°22′ N)。该试验基地自2012年开始作为长期定位试验基地,属于暖温带大陆性季风气候,年平均气温12.7℃,年平均日照2 450 h,无霜期180~210 d,年均降水量328~699 mm,土壤类型为石灰性褐土,耕层土壤容重为1.23 g·cm-3,全量氮、磷、钾和有机质的含量分别为0.07%、0.01%、4.0%和1.4%。
1.2 试验设计
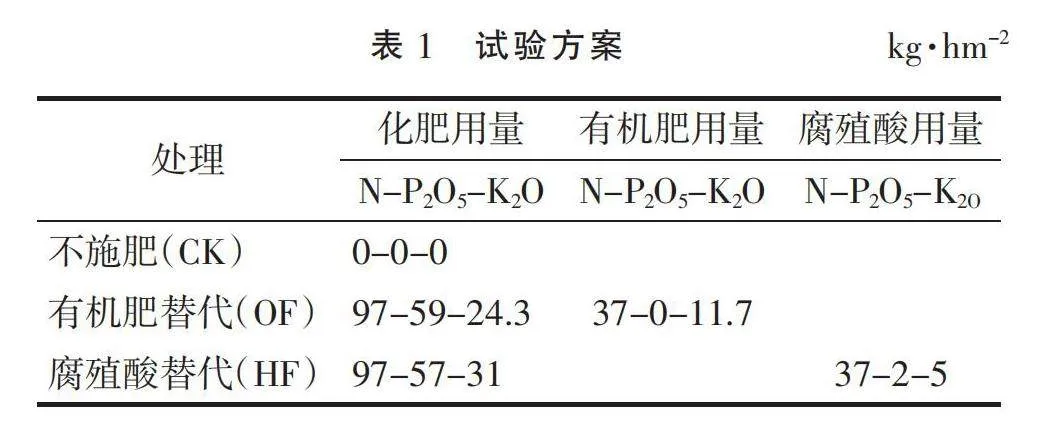
本研究于2020年9月在试验基地进行,试验年限为1个冬小麦生育期,种植冬小麦品种为临丰3号,播种量为150 kg·hm-2。试验共设3个处理,处理1为不施肥(CK),处理2和处理3均在施化肥(尿素:含N 46%、过磷酸钙:含P2O5 16%、氯化钾:含K2O 60%)的基础上分别用有机肥(商品有机肥“地爱沃2号”,OF)和腐殖酸(腐殖酸水溶肥粉剂,HF)替代部分化肥,并遵循N、P、K 等养分量原则,具体施肥量见表1。同时,各处理均采用垄膜沟播的栽培方式,即垄上覆膜、沟内膜侧播种,播种2行,行距20 cm,垄宽35 cm,沟宽30 cm,全生育期保持地膜覆盖,不进行灌溉。供试土壤采集时间为2020年12月,将采集的土壤去除杂物,保存于-80 ℃冰箱中,用于高通量测序。
1.3 测定方法
1.3.1 微生物群落的测定 使用Power Soil 试剂盒采集土壤DNA样本,并参照Power Soil试剂盒的技术要求对其进行分析。采用琼脂糖凝胶电泳法测定DNA 的纯化程度和浓度,将适量试样放入离心管内,用无菌水将待测样品稀释到1 kg·m-3。对稀释后的基因组DNA进行PCR扩增,PCR扩增步骤如下:(1)98 °C预变性1 min;(2)进行30个循环(98 ℃变性10 s,50 ℃退火30 s,72 ℃延伸30 s);(3)在72 ℃条件下,延伸5 min。细菌的扩增引物为338F(5'-ACTCCTACGGGAGGCAGCAG-3')和806R(5'- GGACTACHVGGGTWTCTAAT-3'),真菌的扩增引物为ITS1F(5'-CTTGGTCATTTAGAGGAAGTAA-3')和ITS2R(5'-GCTGCGTTCTTCATCGATGC-3')。基于PCR产物的浓度进行质量混样,并充分混匀样品。然后,用1×TAE缓冲液和2%琼脂糖凝胶电泳提纯PCR目标物质。纯化步骤如下:先在紫外灯下切下含有目的DNA部分的琼脂糖凝胶,称量胶块质量,向胶块中加入Buffer溶液,上述混合液经过离心、洗脱等步骤后,再对PCR产物进行检测定量。采用Thermofisher公司购买的试剂盒进行文库构建,之后检测文库是否合格(Qubit定量),通过后进行样品上机测序(上海美吉生物医药科技有限公司),测序完成后进行数据处理(原始序列的拼接、质控和过滤,OTUs聚类分析,测序深度的检测,群落组成的统计分析)。
1.3.2 功能酶基因的测定 采用宏基因组测序技术鉴定土壤功能酶基因,具体方法如下:利用Soil DNA Kit试剂盒提取土壤DNA,并对其进行浓度和纯度的检测。使用Covaris超声波破碎仪将DNA随机打断为400 bp左右长度的片段,然后进行末端修复、产物纯化、PCR扩增等一系列步骤,完成文库的构建。对文库的Insert size进行初步的定量检测,以确保Insert size符合下一步要求。为了保证文库的可靠度,需准确定量文库的有效浓度。根据文库实际浓度以及最终数据量需求Pooling,通过NovaSeq PE150平台测序。数据下机后,先对原始数据进行数据质控,以获取到高质量的数据。然后对获得的Clean Data进行Metagenome组装。最后,对组装后的数据进行基因预测,从而获得功能基因信息。
1.4 数据统计分析
本研究采用Microsoft Excel 2007软件对数据进行分析,采用IBM SPSS Statistics v22.0软件进行单因素方差分析(One-way ANOVA),利用美吉生物云平台、Origin 2019、R语言进行绘图(Network图除外),运用Cytoscape 3.8.0对微生物群落种间关系进行可视化分析,借助IBM SPSS Statistics v22.0软件对微生物群落进行Spearman相关性分析,筛选出显著相关(Plt;0.05)的两两关系进行绘图。
2 结果与分析
2.1 不同有机替代处理对微生物群落多样性的影响
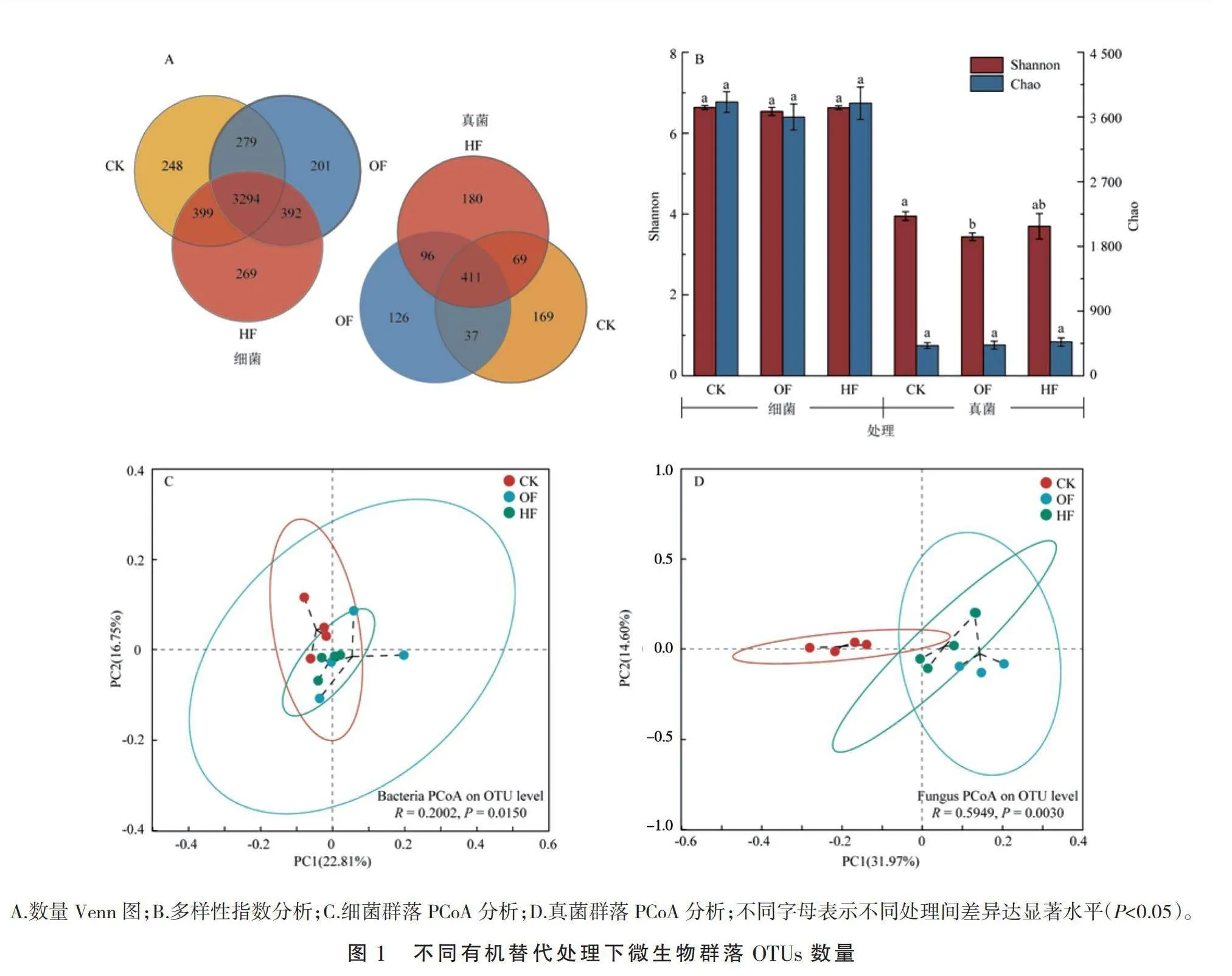
本研究中,细菌和真菌分别获得604 014条和818 403条有效序列,分别平均99 404条和53 200条reads,平均长度分别为417 bp和243 bp。测序序列在97%相似水平下进行聚类操作,细菌和真菌分别获得5 076个和1 084个操作分类单元(OTUs)。各处理的覆盖度均高于99%,并且稀释曲线最终走势趋于平缓,表明测序结果能够很好地反映各样本中微生物群落的真实情况。由维恩图(图1-A)结果可知,HF处理特有的OTUs数最多,细菌和真菌数量分别较CK处理增加8.5%和6.5%;而OF处理特有的OTUs数最少,较CK处理分别降低19%和25%。由α多样性结果可知(图1-B),不同有机替代对真菌群落多样性的影响高于细菌群落。例如,与CK处理相比,HF处理的真菌群落Chao指数增加12%,HF和OF处理的Shannon指数分别降低6%和13%,说明不同有机替代处理可以抑制土壤中真菌群落的多样性,并且OF处理的抑制效果更为明显。然而,不同有机替代处理的细菌群落α多样性差异不显著。在OTUs分类水平下,采用Bray-Curtis距离算法对不同有机替代下的土壤微生物群落进行主坐标分析(PCoA),细菌和真菌坐标轴的总解释度分别为39.56%和46.57%。聚类结果显示,不同处理间的微生物群落各自聚集在一起,其中真菌群落的聚类效果尤为明显。ANOSIM检验发现,细菌和真菌的组间差异性均达显著水平(Plt;0.05),并且真菌的空间变异明显高于细菌群落(图1-C、图1-D)。
2.2 不同有机替代处理对微生物群落组成的影响
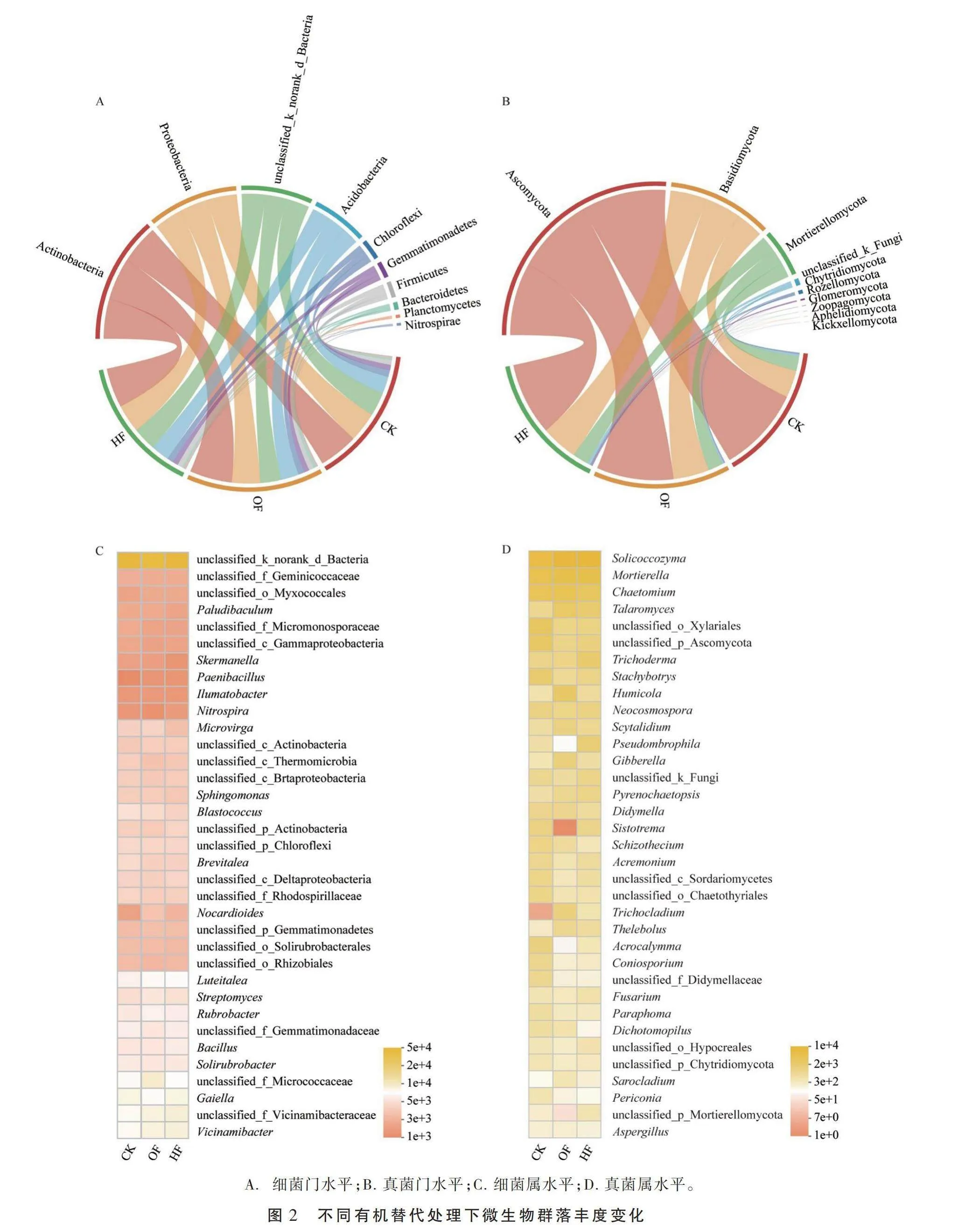
2.2.1 门水平微生物 在门分类水平下,Actino bacteria(30%~32%)、Proteobacteria(21%~23%)、unclassi fied_
k_norank_d_Bacteria(16%~18%)和Acidobacteria(13%~14%)为细菌的优势菌门(图2-A)。与CK处理相比,不同有机替代处理的Actinobacteria和Acidobacteria丰度呈增加趋势,而OF和HF处理的Proteobacteria丰度分别降低4.0%和5.6%。unclassified_k_norank丰度在OF处理中降低12%,而在HF处理中不变。Ascomycota是真菌群落门水平分类下丰度最高的物种,占总体的53%~71%, OF和HF处理的丰度较CK处理分别降低9.4%和15%(图2-B)。Basidiomycota和Mortierellomycota分别为丰度排名第2和第3的真菌门,其丰度在不同有机替代处理中均呈增加趋势。例如,与CK处理相比,OF和HF处理的Basidiomycota丰度分别增加31%和33%。
2.2.2 属水平微生物 选取相对丰度排名前35的属水平细菌(图2-C)(丰度值均在1e+3以上)和真菌(图2-D)(丰度值均在1e+0以上)进行研究。丰度排名前5的细菌分别依次为unclassified_k_norank_d
_Bacteria(17%~19%)、unclassified_f_Vicinamibac-teraceae(3.3%~3.8%)、Vicinamibacter(3.3%~3.8%)、Gaiella(3.2%~3.6%)和unclassified_f_Micrococcaceae(2.9%~4.1%),占总体的41%~43%。与CK处理相比,OF和HF处理的Vicinamibacter丰度分别升高10%和13%。Luteitalea、Bacillus、Streptomyces、Nocardioides和Paenibacillus的丰度变化趋势与Vicinamibacter相同。例如,OF和HF处理的Nocardioides丰度较CK处理分别增加79%和41%。同时,与CK处理相比,OF处理的Nitrospira丰度降低8.5%,而HF处理的Nitrospira丰度增加7.4%。而Microvirga、Paludibaculum、Ilumatobacter在不同处理中的丰度变化趋势与Nitrospira相反。
真菌中,以Solicoccozyma、Mortierella、Chaetomium、Talaromyces和unclassified_o_Xylariales为优势属,占总体的49%~60%。与CK处理相比,各有机替代处理的Solicoccozyma、Mortierella和Talaromyces丰度均呈升高趋势,尤其是OF和HF处理的Talaromyces丰度分别提升210%和137%。同时,Trichoderma、Humicola和Scytalidium丰度在不同处理中的变化趋势与Talaromyces相同。然而,与CK处理相比,OF和HF处理的Stachybotrys、Didymella、Acremonium和Dichotomopilus丰度均呈下降趋势,其平均降幅分别为35%和45%。此外,与CK处理相比,OF处理的 Chaetomium丰度提高17%,而HF处理的 Chaetomium丰度下降21%。与之相反,与CK处理相比,HF处理的Pseudombrophila和Fusarium丰度分别增加244%和30%,而OF处理Pseudombrophila和Fusarium丰度分别下降82%和13%。
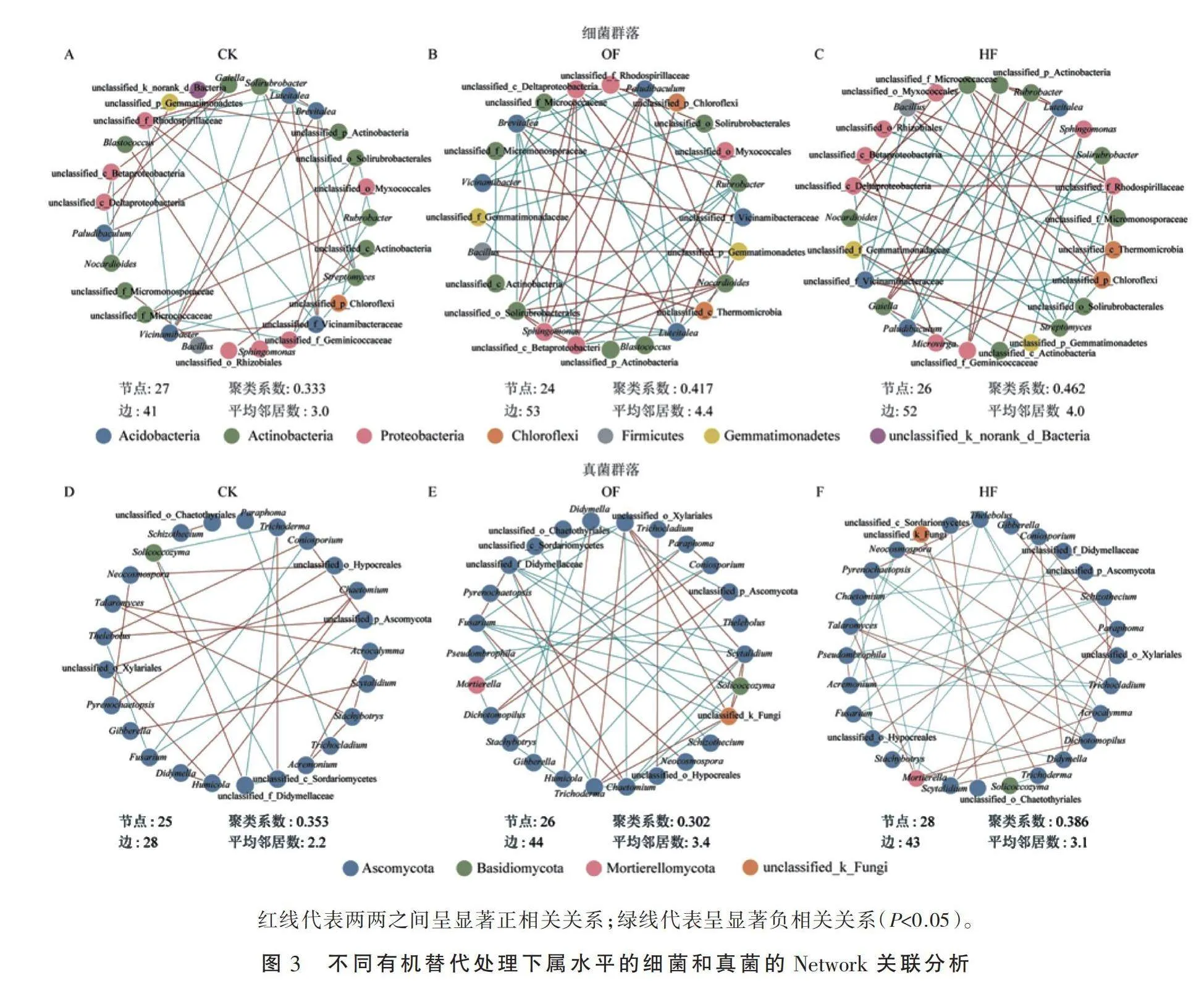
2.3 不同有机替代处理对微生物种间关系的影响
为进一步探明不同有机替代对微生物种间关系的影响,本研究选取各处理排名前30的属水平微生物进行Network关联分析(图3)。由网络拓扑参数可知,与CK处理相比,OF处理和HF处理的细菌群落间的相关关系呈增加趋势,并且HF处理的正相关关系占比最高。同时,HF处理的聚类系数远高于其他处理。这进一步表明HF处理更能增强物种之间的关联度。与CK处理相比,HF处理的 Streptomyces与unclassified_f_Vicinamibacteraceae、Rubrobacter与unclassified_f_Geminicoccaceae种间关系从负相关转变为正相关,而Streptomyces与unclassified_f_Rhodospirillaceae、 Paludibaculum与unclassified_p_Actinobacteria种间关系由正相关转变为负相关。OF处理下,仅有Nocardioides与unclassified_c_Deltaproteobacteria种间关系由负相关转变为正相关。值得注意的是Nocardioides与unclassified_o_Solirubrobacterales在OF处理中表现为正相关关系,在HF处理中呈负相关关系。真菌群落的种间关系与细菌群落类似,即有机替代处理的相关关系较CK处理有所增加(图3-D、图3-E、图3-F)。然而,与CK处理相比,其正相关关系较OF处理(44%)和HF处理(48%)分别下降39%和33%。经OF处理后,unclassified_f_Didymellaceae与Solicoccozy种间关系均从正相关转变为负相关,Trichoderma与Solicoccozyma种间关系由负相关转变为正相关。OF处理下,Fusarium、Chaetomium和Mortierella分别与多个属水平真菌展现出负相关关系,如Fusarium与Solicoccozyma、unclassified_k_Fungi
和Trichoderma呈现负相关。HF处理下,Trichocladium与Acremonium和Pseudombrophila、Pyrenochaetopsis与Acrocalymma和Talaromyces均表现为负相关关系。
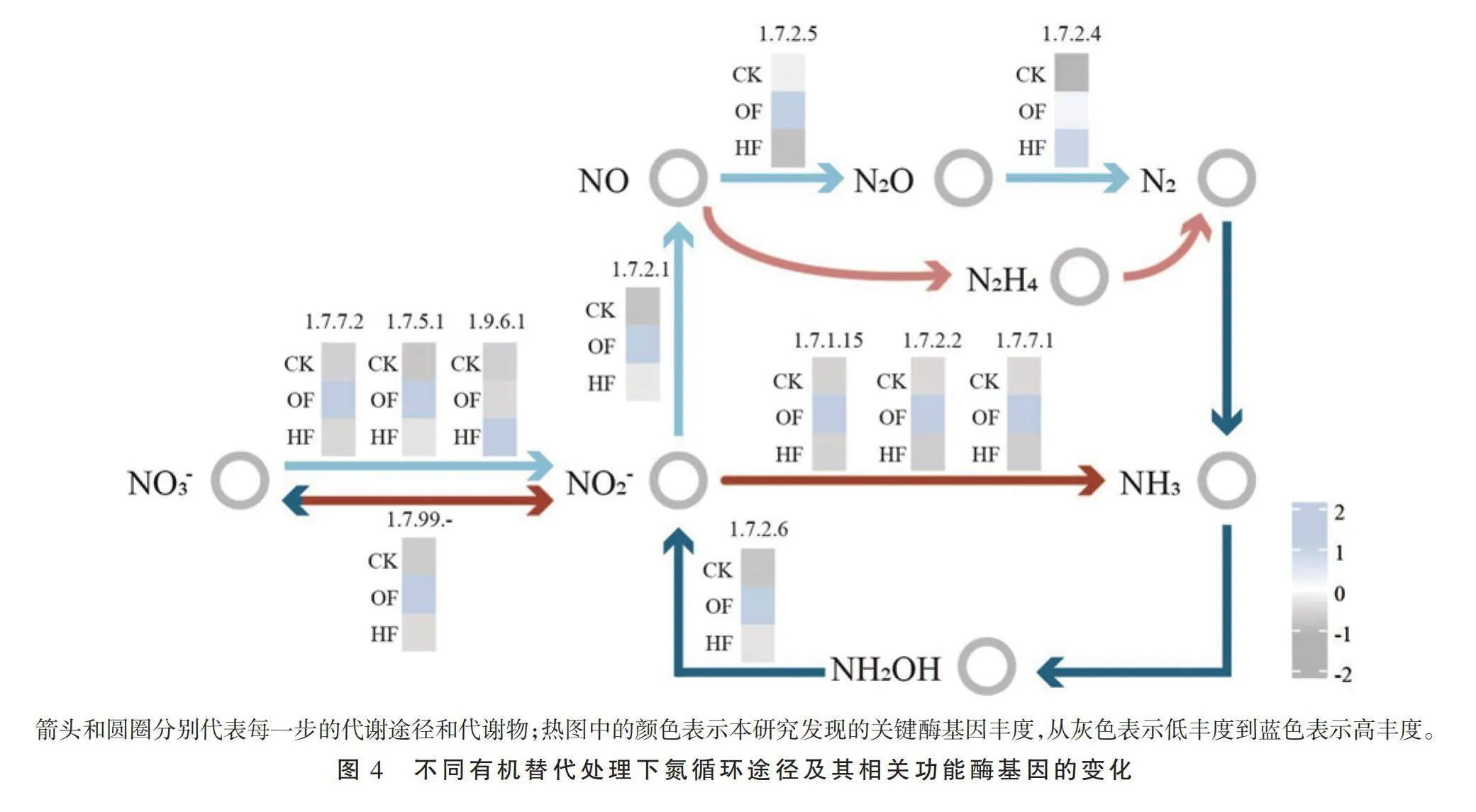
2.4 不同有机替代处理对氮循环功能酶基因的影响
本研究共发现11个参与氮循环的功能酶基因(图4),包括2个参与硝化作用的酶基因EC 1.7.2.6和EC 1.7.99.-,3个参与异化性硝酸盐还原作用的酶基因EC 1.7.7.2、EC 1.7.5.1和EC 1.9.6.1,3个参与亚硝酸盐氨化的酶基因EC 1.7.1.15、EC 1.7.7.1和EC 1.7.2.2,以及3个参与反硝化作用的酶基因EC 1.7.2.1、EC 1.7.2.5和EC 1.7.2.4。与CK处理相比,HF处理的功能酶基因总丰度变化不大,但OF处理的功能酶基因总丰度增加57%。尤其是OF处理的EC 1.7.2.6丰度变化最明显,较CK处理增加305%。3 讨论与结论
3.1 不同有机替代对功能微生物的影响
植物病原菌是造成农作物大幅减产的重要因素[17]。Didymella sp.、Fusarium sp.和Acremonium sp.作为常见的植物病原菌在本研究中被发现,Didymella sp.会引发玉米[18]和开心果[19]叶枯病,Fusarium sp.富集会导致小麦和玉米等粮食作物根系病害的发生(根腐、茎腐)和根表面的损害(萎蔫)[20-21],Acremonium sp.会引发夏威夷果速衰病[22]以及高粱、玉米、草莓坏死及枯萎[23]。本研究中,较CK处理相比,不同有机替代处理均降低了潜在植物病原菌Didymella和Acremonium丰度,而Fusarium丰度在有机肥替代处理中降低,在腐殖酸替代处理中升高。这表明各有机替代处理具有抑制植物病原菌的潜力,并且有机肥替代较腐殖酸的抑制效果相对更好。
生物防治技术是利用一种或多种微生物来抑制病原菌生命活力和繁殖能力的方法[24-25]。Streptomyces sp.和Paenibacillus sp.作为常见的生物防治菌在本研究中被发现,Streptomyces sp.是Actinobacteria中的一种生物防治菌,可以抵制多种病菌对植物的侵染。前人研究发现,对浙麦冬黑斑病病原菌链格孢菌[26]、水稻白叶枯病菌[27]、马铃薯枯萎病镰刀菌[28]等病原菌能产生明显的抑制效果。Paenibacillus sp.广泛存在于植物根际土壤中,对植物病原微生物具有拮抗作用,可以被应用于生物防治,保护植物免受枯萎病和青枯病的侵害[29],可通过调节根际微生物进而抑制黄瓜枯萎病[30]。上述生物防治细菌在各有机替代处理中的丰度均呈增加趋势,并且在有机肥替代处理中增幅最大。
Chaetomium sp.、Trichoderma sp.和Talaromyces sp.是常见的生物防治真菌。Zhang等[31]研究表明,Chaetomium sp.可以防治枯萎病,如抑制谷物幼苗和甘蔗枯萎,降低番茄枯萎病和尖孢镰刀菌引起的枯萎病、苹果斑病引起的枯萎病、杨树枯萎病引起的根腐病的发病率。Trichoderma sp.可以产生或释放多种化合物,激活植物免疫力,以防止病原体感染[32],对早疫病、晚疫病、枯萎病、黑胫病、纹枯病和促倒病均有较好的生物防治作用[29]。Talaromyces sp.代谢产生的有机物质对控制土传病害的发生具有重要作用,可以促进玉米和水稻生长[33],保护水稻作物免受水稻鞘枯病的危害[34],具有较好的防治效果。Naraghi等[35]研究证实,Talaromyces sp.对于由大丽轮枝孢引起的棉花和马铃薯枯萎病的防治具有较好的效果。本研究中,Talaromyces和Trichoderma的丰度在不同有机替代处理中均升高;Chaetomium的丰度在有机肥替代处理中增加,而在腐殖酸处理降低。不难发现,相较于腐殖酸替代处理,有机肥替代处理更有利于潜在生物防治菌的生长,这将有利于对土传病害的生物防治。
土壤微生物是陆地生态系统的重要组成部分,其多样性和群落组成不仅反映了土壤生态系统的健康状况和功能,也反映了土壤肥力状况[36-37]。Nocardioides sp.不仅可参与有机物的降解,而且具有一定的固氮能力[38]。Bacillus sp.具有解磷的能力,可分解土壤中的有机物(纤维素和木质素),并通过分泌次生代谢物诱导植物产生抗病性[39],抑制土壤病原体传播,促进植物生长[30]。Chaetomium sp.参与了土壤氮循环过程[31],Trichoderma sp.促进植物根系生长,提高氮磷含量[32],Talaromyces sp.有着高效的有机磷矿化能力[33]。本研究中,上述微生物的丰度在有机肥替代处理中较腐殖酸替代处理均有较大幅的升高。这说明有机肥替代在削弱潜在土传病害菌和增强土壤肥力方面展示出更强的协同效益[40-41]。
3.2 不同有机替代处理对微生物种间关系的影响
通过Network关联分析发现,除OF处理的真菌群落聚类系数比CK处理降低外,不同有机替代处理的平均邻居数、网络密度、聚类系数均明显升高。这表明经不同有机替代处理后土壤微生物群落形成了一个更为复杂的网络结构。前人研究发现,土壤养分对微生物群落有很大的影响[42]。本研究中,有机肥和腐殖酸替代处理后,土壤微生物群落形成了更为复杂的网络结构。例如,优势细菌Streptomyces与unclassified_f_Vicinamibacteraceae、 Rubrobacter与unclassified_f_Geminicoccaceae由拮抗作用变为协同作用,从而增加了土壤微生物间的协作水平,进而强化了土壤微生物抵抗环境干扰的能力[43]。生物防治菌与病原菌的拮抗效应可以有效防治土传病害的发生[44-45]。本研究中,有机肥替代处理下,潜在植物病原菌Fusarium、潜在生物防治菌Trichoderma和具有土壤溶磷能力的真菌Solicoccozyma呈拮抗作用。腐殖酸替代处理下,Trichocladium与植物致病菌Acremonium表现为负相关关系。这进一步表明不同有机替代对土传病害有一定的抑制潜力。同时,潜在生物防治菌Trichoderma和潜在解磷菌Solicoccozyma在有机肥替代处理中呈正相关关系。这说明微生物在不同有机替代处理中形成了更复杂的共存网络,并增强了功能菌的互作关系。
3.3 不同有机替代对氮循环功能酶基因的影响
氮循环过程包括异化硝酸盐还原、硝化、反硝化和固氮[46],而酶是氮转化过程的重要驱动因素[47]。研究发现,氮循环功能基因在不同有机替代中呈增长趋势,其中有机肥替代处理的增幅得更明显。亚硝酸盐还原酶EC 1.7.2.2与铵态氮的生成有关[48]。本研究中,EC 1.7.2.2的丰度在有机肥替代处理中升高,而在腐殖酸替代处理中降低。这说明有机肥替代具有增强铵态氮的潜力。同时,硝化过程中羟胺脱氢酶EC 1.7.2.6催化羟胺生成亚硝酸盐,在酶基因EC 1.7.5.1的驱动下,亚硝酸盐进一步转化为硝酸盐。硝酸盐是植物所需的主要氮源之一,促使植物合成氨基酸、蛋白质和叶绿素,对植物的生长发育至关重要[49]。本研究中,不同有机替代处理下,EC 1.7.2.6和EC 1.7.5.1的丰度较CK明显增加,并且有机肥替代处理的增幅远高于腐殖酸替代处理。这说明有机肥替代具有较高的供氮潜力。
3.4 结论
(1)不同有机替代处理改变了旱地麦田土壤微生物群落的多样性和结构,并且对真菌群落的影响更为明显。
(2)不同有机肥替代处理调控了微生物的种间关系,增强了潜在生物防治菌(如Streptomyces、Paenibacillus、Chaetomium和Talaromyces)丰度,有利于提高土壤健康和作物抵抗力;抑制了潜在植物病原菌(如Fusarium和Acremonium)生长,降低了病害发生风险,并且有机肥替代处理效果更明显。
(3)不同有机替代(尤其是有机肥替代)处理增强了潜在参与养分转化的功能微生物(EC 1.7.2.6和EC 1.7.99.-)活性,有利于氮素养分供应,提高了参与氮循环过程酶基因丰度。
参考文献:
[1] ZHOU Y, CHEN Z X, CHENG M P, et al. Uncovering the dispersion history, adaptive evolution and selection of wheat in China[J]. Plant Biotechnology Journal, 2018, 16(1): 280-291.
[2] LIU H, XIONG W, MOTTALEB K A, et al. Contrasting contributions of five factors to wheat yield growth in China by process-based and statistical models[J]. European Journal of Agronomy, 2021, 130: 126370.
[3] PAN H, CHEN M M, FENG H J, et al. Organic and inorganic fertilizers respectively drive bacterial and fungal community compositions in a fluvo-aquic soil in northern China[J]. Soil and Tillage Research, 2020, 198: 104540.
[4] FERNANDEZ A L, SHEAFFER C C, WYSE D L, et al. Associations between soil bacterial community structure and nutrient cycling functions in long-term organic farm soils following cover crop and organic fertilizer amendment[J]. Science of the Total Environment, 2016, 566/567: 949-959.
[5] ZHOU J, LI B, XIA L L, et al. Organic-substitute strategies reduced carbon and reactive nitrogen footprints and gained net ecosystem economic benefit for intensive vegetable production[J]. Journal of Cleaner Production, 2019, 225: 984-994.
[6] REN J H, LIU X L, YANG W P, et al. Rhizosphere soil properties, microbial community, and enzyme activities: short-term responses to partial substitution of chemical fertilizer with organic manure[J]. Journal of Environmental Management, 2021, 299: 113650.
[7] EKIN Z. Integrated use of humic acid and plant growth promoting rhizobacteria to ensure higher potato productivity in sustainable agriculture[J]. Sustainability, 2019, 11(12): 3417.
[8] REN H Y, ISLAM M S, WANG H Y, et al. Effect of humic acid on soil physical and chemical properties, microbial community structure, and metabolites of decline diseased bayberry[J]. International Journal of Molecular Sciences, 2022, 23(23): 14707.
[9] DA CUNHA LEME FILHO J F, THOMASON W E, EVANYLO G K, et al. The synergistic effects of humic substances and biofertilizers on plant development and microbial activity: a review[J]. International Journal of Plant amp; Soil Science, 2020, 32(7): 56-75.
[10] KHAN R U, KHAN M Z, KHAN A, et al. Effect of humic acid on growth and crop nutrient status of wheat on two different soils[J]. Journal of Plant Nutrition, 2018, 41(4): 453-460.
[11] TIWARI J, RAMANATHAN A L, BAUDDH K, et al. Humic substances: structure, function and benefits for agroecosystems—a review[J]. Pedosphere, 2023, 33(2): 237-249.
[12] LI Y L, TREMBLAY J, BAINARD L D, et al. Long-term effects of nitrogen and phosphorus fertilization on soil microbial community structure and function under continuous wheat production[J]. Environmental Microbiology, 2020, 22(3): 1066-1088.
[13] LIU H Y, XU W, LI J Y, et al. Short-term effect of manure and straw application on bacterial and fungal community compositions and abundances in an acidic paddy soil[J]. Journal of Soils and Sediments, 2021, 21(9): 3057-3071.
[14] 王伟华, 刘毅, 唐海明, 等. 长期施肥对稻田土壤微生物量、 群落结构和活性的影响[J]. 环境科学, 2018, 39(1): 430-437.
[15] HARTMANN M, FREY B, MAYER J, et al. Distinct soil microbial diversity under long-term organic and conventional farming[J]. The ISME Journal, 2015, 9(5): 1177-1194.
[16] 姜丽伟, 谢英荷, 李廷亮, 等. 不同有机替代对黄土旱塬土壤碳、氮组分及冬小麦产量的影响[J]. 水土保持学报, 2022, 36(3): 286-293.
[17] PANTH M, HASSLER S C, BAYSAL-GUREL F. Methods for management of soilborne diseases in crop production[J]. Agriculture, 2020, 10(1): 16.
[18] MA W D, YANG J, DING J Q, et al. Gapless reference genome assembly of Didymella glomerata, a new fungal pathogen of maize causing Didymella leaf blight[J]. Frontiers in Plant Science, 2022, 13: 1022819.
[19] MORAL J, LICHTEMBERG P S F, PAPAGELIS A, et al. Didymella glomerata causing leaf blight on pistachio[J]. European Journal of Plant Pathology, 2018, 151(4): 1095-1099.
[20] WINTER M, SAMUELS P L, DONG Y, et al. Trichothecene production is detrimental to early root colonization by Fusarium culmorum and F. graminearum in fusarium crown and root rot of wheat[J]. Plant Pathology, 2019, 68(1): 185-195.
[21] FERRIGO D, MONDIN M, LADURNER E, et al. Effect of seed biopriming with Trichoderma harzianum strain INAT11 on Fusarium ear rot and Gibberella ear rot diseases[J]. Biological Control, 2020, 147: 104286.
[22] YAO X F, LIU Q M, LIU Y X, et al. Managing macadamia decline: a review and proposed biological control strategies[J]. Agronomy, 2024, 14(2): 308.
[23] RACEDO J, SALAZAR S M, CASTAGNARO A P, et al. A strawberry disease caused by Acremonium strictum[J]. European Journal of Plant Pathology, 2013, 137(4): 649-654.
[24] GULLINO M L, ALBAJES R, NICOT P C. Integrated pest and disease management in greenhouse crops[M]. 2nd ed. Cham: Springer, 2020.
[25] CONDE-CID M, ÁLVAREZ-ESMORÍS C, PARADELO-NUÑEZ R, et al. Occurrence of tetracyclines and sulfonamides in manures, agricultural soils and crops from different areas in Galicia (NW Spain)[J]. Journal of Cleaner Production, 2018, 197(Part 1): 491-500.
[26] 徐亦雯, 张诺, 曹瑱艳, 等. 浙麦冬黑斑病病原菌的分离鉴定及淀粉酶产色链霉菌对其生物防治效果[J]. 中国生物防治学报, 2022, 38(5): 1280-1287.
[27] 鲍莹, 严梦圆, 吴萌, 等. 腐殖酸配施壳聚糖调控设施番茄土壤细菌群落[J]. 农业环境科学学报, 2022, 41(12): 2772-2778.
[28] CASTAÑO R, BORRERO C, TRILLAS M I, et al. Selection of biological control agents against tomato Fusarium wilt and evaluation in greenhouse conditions of two selected agents in three growing media[J]. BioControl, 2013, 58(1): 105-116.
[29] 曹坳程, 张大琪, 方文生, 等. 土传病害防治技术进展及面临的挑战[J]. 植物保护, 2023, 49(5): 260-269.
[30] SHI L, DU N S, SHU S, et al. Paenibacillus polymyxa NSY50 suppresses Fusarium wilt in cucumbers by regulating the rhizospheric microbial community[J]. Scientific Reports, 2017, 7(1): 41234.
[31] ZHANG S S, CHEN Y L, TUO Y Y, et al. Effects of lime nitrogen on the fungi community of tobacco soil and its correlation analysis[J]. IOP Conference Series: Earth and Environmental Science, 2022, 1087(1): 012059.
[32] SALWAN R, SHARMA A, KAUR R, et al. The riddles of Trichoderma induced plant immunity[J]. Biological Control, 2022, 174: 105037.
[33] SUN X F, LIU F, JIANG W, et al. Talaromyces purpurogenus isolated from rhizosphere soil of maize has efficient organic phosphate-mineralizing and plant growth-promoting abilities[J]. Sustainability, 2023, 15(7): 5961.
[34] ABBAS A, FU Y P, QU Z, et al. Isolation and evaluation of the biocontrol potential of Talaromyces spp. against rice sheath blight guided by soil microbiome[J]. Environmental Microbiology, 2021, 23(10): 5946-5961.
[35] NARAGHI L, HEYDARI A, REZAEE S, et al. Biocontrol agent Talaromyces flavus stimulates the growth of cotton and potato[J]. Journal of Plant Growth Regulation, 2012, 31(4): 471-477.
[36] MOREIRA H, PEREIRA S I A, VEGA A, et al. Synergistic effects of arbuscular mycorrhizal fungi and plant growth-promoting bacteria benefit maize growth under increasing soil salinity[J]. Journal of Environmental Management, 2020, 257: 109982.
[37] MANASA M R K, KATUKURI N R, DARVEEKARAN NAIR S S, et al. Role of biochar and organic substrates in enhancing the functional characteristics and microbial community in a saline soil[J]. Journal of Environmental Management, 2020, 269: 110737.
[38] LIU S L, LI H M, XIE X Y, et al. Long-term moderate fertilization increases the complexity of soil microbial community and promotes regulation of phosphorus cycling genes to improve the availability of phosphorus in acid soil[J]. Applied Soil Ecology, 2024, 194: 105178.
[39] MAJHI B, SEMWAL P, MISHRA S K, et al. Arsenic stress management through arsenite and arsenate-tolerant growth-promoting bacteria in rice[J/OL]. International Microbiology: 1-15[2024-09-10]. https://doi.org/10.1007/
s10123-023-00447-w.
[40] URKURKAR J S, TIWARI A, CHITALE S, et al. Influence of long-term use of inorganic and organic manures on soil fertility and sustainable productivity of rice (Oryza sativa) and wheat (Triticum aestivum) in Inceptisols[J]. The Indian Journal of Agricultural Sciences, 2010, 80(3): 208-212.
[41] WANG Y J, XU Y, JIANG L H, et al. Effect of mild organic substitution on soil quality and microbial community[J]. Agronomy, 2024, 14(5): 888.
[42] FERNANDEZ C W, KENNEDY P G. Revisiting the ‘Gadgil effect’: do interguild fungal interactions control carbon cycling in forest soils?[J]. New Phytologist, 2016, 209(4): 1382-1394.
[43] SANTOLINI M, BARABÁSI A L. Predicting perturbation patterns from the topology of biological networks[J]. Proceedings of the National Academy of Sciences of the United States of America, 2018, 115(27): E6375-E6383.
[44] MANJUNATHA S V, NAIK M K, KHAN M F R, et al. Evaluation of bio-control agents for management of dry root rot of chickpea caused by Macrophomina phaseolina[J]. Crop Protection, 2013, 45: 147-150.
[45] SINGH S, KUMAR R, YADAV S, et al. Effect of bio-control agents on soil borne pathogens: a review[J]. Journal of Pharmacognosy and Phytochemistry, 2018, 7(3): 406-411.
[46] WANG S S, NIU Q Q, ZHU P F, et al. Metagenomics analysis unraveled the influence of sulfate radical-mediated compost nitrogen transformation process[J]. Journal of Environmental Management, 2022, 317: 115436.
[47] SUN X D, YE Y Q, MA Q X, et al. Variation in enzyme activities involved in carbon and nitrogen cycling in rhizosphere and bulk soil after organic mulching[J]. Rhizosphere, 2021, 19: 100376.
[48] GUO H H, GU J, WANG X J, et al. Beneficial effects of bacterial agent/bentonite on nitrogen transformation and microbial community dynamics during aerobic composting of pig manure[J]. Bioresource Technology, 2020, 298: 122384.
[49] ISLAM S, ISLAM R, KANDWAL P, et al. Nitrate transport and assimilation in plants: a potential review[J]. Archives of Agronomy and soil Science, 2022, 68(1): 133-150.
猜你喜欢
今日农业(2022年16期)2022-11-09 23:18:44
今日农业(2021年12期)2021-11-28 15:49:26
今日农业(2021年8期)2021-11-28 05:07:50
金桥(2021年10期)2021-11-05 07:23:28
今日农业(2021年13期)2021-08-14 01:38:00
作文小学中年级(2020年4期)2020-06-11 12:47:08
中小企业管理与科技·下旬刊(2016年11期)2016-11-18 08:19:35
中国科技博览(2016年19期)2016-10-19 11:47:05
中国实用医药(2016年15期)2016-05-24 13:57:34
科技视界(2016年9期)2016-04-26 12:23:48
