
吕梁黑猪BACE2基因克隆及表达分析
2024-12-31 00:00:00杨梓杰张晋赵丹
安徽农学通报 2024年22期
关键词:生物信息学



摘要" 为探讨吕梁黑猪BACE2基因特点和结构,本研究采用PCR技术对该基因进行克隆,并对所编码的蛋白质开展生物信息学分析,分析内容包括蛋白质的理化性质、信号肽、亲疏水性、跨膜结构、序列比对、二、三级结构预测、亚细胞定位预测,以及互作关系;通过实时荧光定量PCR(qPCR)技术,研究BACE2基因在吕梁黑猪的心脏、肝脏、脾脏、肺脏、肾脏、肌肉和脂肪组织中的表达情况。生物信息学结果显示,该黑猪BACE2基因的编码序列大小(CDS)为1 545"bp,包含514个氨基酸;BACE2蛋白是一种疏水性蛋白,无信号肽,具有明显的跨膜作用,是一种非分泌型的蛋白质;序列比对发现吕梁黑猪同山羊、牛亲缘关系较近,同家鼠亲缘关系较远;蛋白的二级结构由无规则卷曲(49.03%)、α-螺旋(18.48%)、延伸连(27.24%)和β-转角(5.25%)构成,主要分布于内质网(55.6 %)。qPCR 结果显示,其肾脏组织中BACE2基因的表达水平最高,肌肉组织中的表达量最低。研究结果揭示了BACE2蛋白拥有的多种独特属性及BACE2基因的分布特点,为该蛋白更深层次的科学探究提供参考。
关键词" 吕梁黑猪;BACE2基因;生物信息学
中图分类号" S85"""""" 文献标识码" A"""""" 文章编号" 1007-7731(2024)22-0043-07
DOI号" 10.16377/j.cnki.issn1007-7731.2024.22.010
基金项目 山西省教育科学“十四五”规划课题“高质量教育体系下高校动医专业基础课程实验数字化教学探索与实践”(SZH-230078)。
作者简介 杨梓杰(2003—)男,山西太原人,从事动物医学研究。
通信作者 赵丹(1986—),男,山西太原人,硕士,讲师,从事动物生物化学研究。
收稿日期 2024-08-07
Cloning and expression analysis of BACE2 gene in Lyuliang black pig
YANG Zijie""" ZHANG Jin""" ZHAO Dan
(Jinzhong College of Information, Taigu 030800, China)
Abstract" In order to investigate the characteristics and structure of the BACE2 gene in Lyuliang black pigs, the BACE2 gene was cloned by PCR technology,and the encoded protein was analyzed by bioinformatics, including physical and chemical properties, signal peptide, hydrophobicity, transmembrane structure, sequence alignment, secondary and tertiary structure prediction, subcellular localization, and interaction. The expression of the BACE2 gene in the heart, liver, spleen, lung, kidney, muscle and adipose tissue of this pigs was measured by real-time quantitative PCR. The results of bioinformatics showed that the coding sequence (CDS) of BACE2 gene size was 1 545"bp, containing 514"amino acids. BACE2 protein is a hydrophobic protein, no signal peptide, has obvious transmembrane function, and is a non-secretory protein. Sequence alignment showed that the pig had a close genetic relationship with goat and cattle, and the furthest genetic relationship with Mus musculus. The secondary structure of the protein was composed of random curl (49.03%), α-helix (18.48%), extension link (27.24%) and β-corner (5.25%), mainly distributed in the endoplasmic reticulum (55.6%).qPCR results showed that the expression level of BACE2 gene was the higher in kidney tissue and the lower in muscle tissue of this pigs. The results revealed a variety of unique properties of BACE2 protein and the distribution characteristics of BACE2 gene, which provided a reference for further scientific exploration of this protein.
Keywords" Lyuliang black pigs; BACE2 gene; bioinformatics
猪是一种脊椎动物,也是杂食类哺乳动物。由于品种的不同,其体貌特征也各不相同,通常以耳大、头长、四肢短小、鼻直、身体肥壮和腰背窄为主要形体特征[1]。吕梁黑猪是引进生猪进行自繁自养得到的品种,其全身体毛黑色、结构匀称、四肢健壮,头型稍长,额部有横行皱纹,具有肉质鲜美、抗病力强等特点[2]。BACE2(Beta-site app-cleaving enzyme 2,BACE2)是BACE1的同源物,是一种θ分泌酶,也是一种条件性β分泌酶,其能够水解β淀粉样前体蛋白(β-amyloid precursor protein,APP)产生β淀粉样蛋白(Amyloid-β peptide,Aβ)[3]。在体外,发现BACE2是一种特别活跃的Aβ降解蛋白酶(Aβ-degrading proteases,AβDP),其催化效率超过了除胰岛素降解酶(Insulin degrading enzymes,IDE)以外的已知的 AβDPs[3],是治疗或预防阿尔兹海默症(Alzheimer’s disease,AD)的一种强有力的候选药物。谢建等[4]研究表明,敲除BACE2可抑制细胞侵袭、迁移和肿瘤生长,过表达BACE2可促进细胞间质转化和增殖。王会志[5]通过BACE2激活NF-κB通路促进胶质瘤恶性进展的机制研究,发现BACE2在胶质瘤的发生与发展中扮演关键角色,敲低BACE2可减弱胶质瘤侵袭迁移及增殖能力,是治疗该疾病的潜在靶点。
目前,将吕梁黑猪作为研究对象的相关研究相对较少,在其肌肉发育和脂肪沉积中,BACE2基因的作用机制有待进行充分研究。因此,对其BACE2基因的生物信息学进行分析,有助于深入理解该物种在遗传层面的特性,尤其是在肌肉发育和脂肪沉积等性状方面。本研究采用PCR技术对BACE2基因进行克隆,并对所编码的蛋白质开展生物信息学分析,通过实时荧光定量PCR(qPCR)技术,研究BACE2在吕梁黑猪的心脏、肝脏、脾脏、肺脏、肾脏、卵巢、肌肉和脂肪组织中的表达情况,为BACE2蛋白更深层次的科学探究提供参考,为该猪种的保种和改良工作提供分子生物学层面的数据参考。
1 材料与方法
1.1 试验材料
在山西吕梁莱芜黑猪养殖基地,挑选3头体质强健的吕梁黑猪作为试验对象。屠宰后,迅速采集其心脏、肝脏、脾脏、肺脏、肾脏、卵巢、肌肉以及脂肪8个组织。为确保组织的完整性和活性,采集后立即放入液氮中进行冷冻保存。随后,将组织样本运送回实验室,转移至-80"℃冰箱中,备用。
1.2 试验仪器与试剂
PCR仪(NEW VeritiPro)、电泳仪(GH600E)、超微量分光光度计(NanoDrop 2000),凝胶成像系统(Invitrogen E-Gel Power Snap G8100);DL2000 DNA Marker、LATaq DNA聚合酶以及M-MLV反转录测试盒,均购自宝日医生物技术(北京)有限公司。
1.3 试验方法
1.3.1 总RNA提取及反转录 采用Trizol法提取组织的总RNA。采用超微量分光光度计NanoDrop 2000检测RNA的质量和浓度,其OD260和OD280在1.8~2.0。通过以上处理,得到一个完备且有代表性的核酸序列数据,满足相关必要的标准后,按照M-MLV反转录测试盒的说明书进行反转录,反转录得到的cDNA样品存置于-20"℃的冰箱,备用。
1.3.2 BACE2基因引物设计与合成 基于GenBank提供的BACE2基因序列(登记编号:XP 020926661.1.1),设计用于克隆和实时荧光定量PCR(qPCR)的引物。引物序列送至西安擎科生物科技公司合成,详情如表1所示。
1.3.3 BACE2基因的克隆与测序 PCR试验以吕梁黑猪肝脏组织cDNA为模板进行特异性扩增。反应总体系为25 μL,具体组成:ddH2O 8.5 μL,2×TaqPCR Master Mix 12.5 μL,cDNA 2 μL,上下游引物各1 μL(浓度10 μmol/L)。扩增条件:首先94"℃预变性30"s;然后进行35个扩增循环,每个循环98"℃变性10"s,56.02"℃退火30"s,72"℃延长1"min,循环结束后72"℃延伸2"min。将产物保存于4"℃冰箱。为验证扩增结果的准确性,采用高达琼脂糖凝胶电泳及凝胶成像系统对目的条带进行检测,将符合预期的条带送至西安擎科生物科技公司进行测序分析。
1.3.4 BACE2基因的生物信息学分析 采用生物信息学方法对测序分析得到的氨基酸残基序列及其次级结构进行了深入研究,以揭示蛋白质的空间结构特点,为后续的实验研究提供了一定的数据支撑。使用Protparam工具,分析蛋白质理化性质。在https//prosite.expasy.org网站查找蛋白质结构的研究资料。采用基于密度泛函原理的策略,对蛋白质的分子构造与其绑定作用之间的联系进行初步研究。通过SignaIP软件预测蛋白质信号肽;通过https://web.expasy.org/protscale预测蛋白质的亲疏水性;利用TMHMM2.0软件进行蛋白质的跨膜结构预测;利用NCBI软件里的BLASTX工具进行蛋白质同源性比对,利用MEGA7软件绘制蛋白质演变树状结构。通过https://predictprotein.org预测蛋白质的次级结构;通过https://swissmodel.expasy.org预测蛋白质三级结构。通过PSORT 在线软件进行蛋白亚细胞定位。基于以上数据,应用机械学习和人工神经网络的技术,成功建立了一个以计算机模拟为核心的三维蛋白质模型。利用在线软件STRING对来自多个数据平台的蛋白质氨基酸组成及其之间的功能性关联进行详细的比较分析,构建BACE2蛋白互作网络。
1.3.5 实时荧光定量PCR 以8个组织的cDNA为模板,GAPDH为内参,进行qPCR检测。试验的反应总体系为20 µL,详细组分:10 µL的2× SYBR Green Pro Taq HS Premix,上游和下游引物(浓度为10 µmol/µL)各0.4 µL,cDNA模板1 µL,以及8.2 µL ddH2O。反应程序:首先进行95"℃预变性30"s;随后进行40个扩增循环,每个循环包括95"℃变性5"s,57"℃退火30"s,65"℃延伸5"s;循环结束后95"℃延伸5"min。
1.4 数据处理
用Excel软件和2-ΔΔCt方法对qPCR结果进行统计分析。
2 结果与分析
2.1 BACE2基因克隆与测序
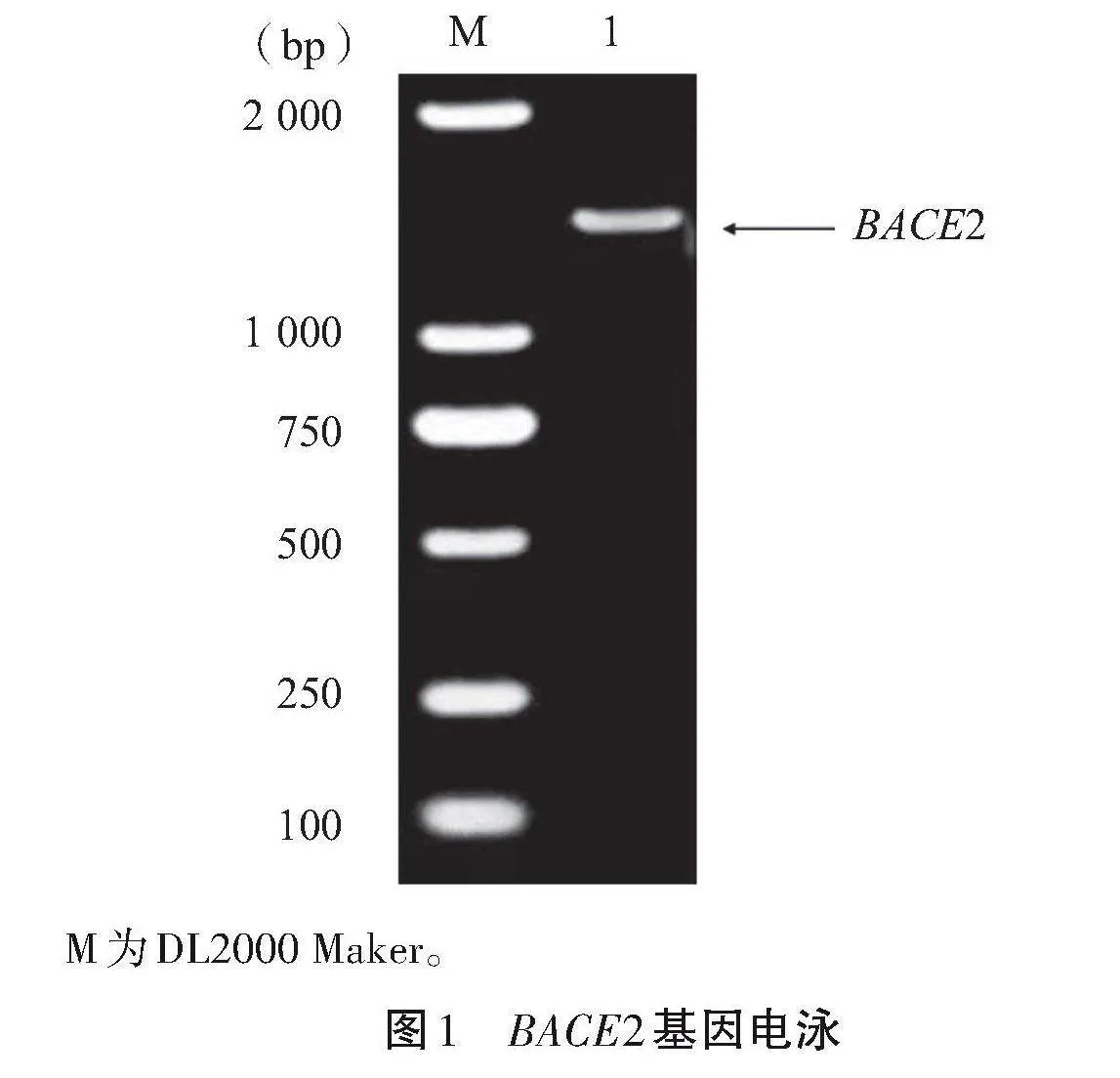
以cDNA为模板,利用特异性引物进行PCR扩增该基因片段,1%琼脂糖凝胶电泳检测结果显示,目的条带大小约为1 500"bp,符合预期(图1)。收集测序数据后,采用DNAstar软件中的Seqman子程序来完成序列的拼接工作。将拼接序列与NCBI数据库中BACE2基因的CDS区使用BLAST软件进行比对,结果表明,吕梁黑猪的BACE2基因编码区域长度达到了1 545"bp。基于开放阅读框(ORF Finder)的计算,这一序列被编码为514个氨基酸。
M为DL2000 Maker。
2.2 BACE2蛋白的理化性质
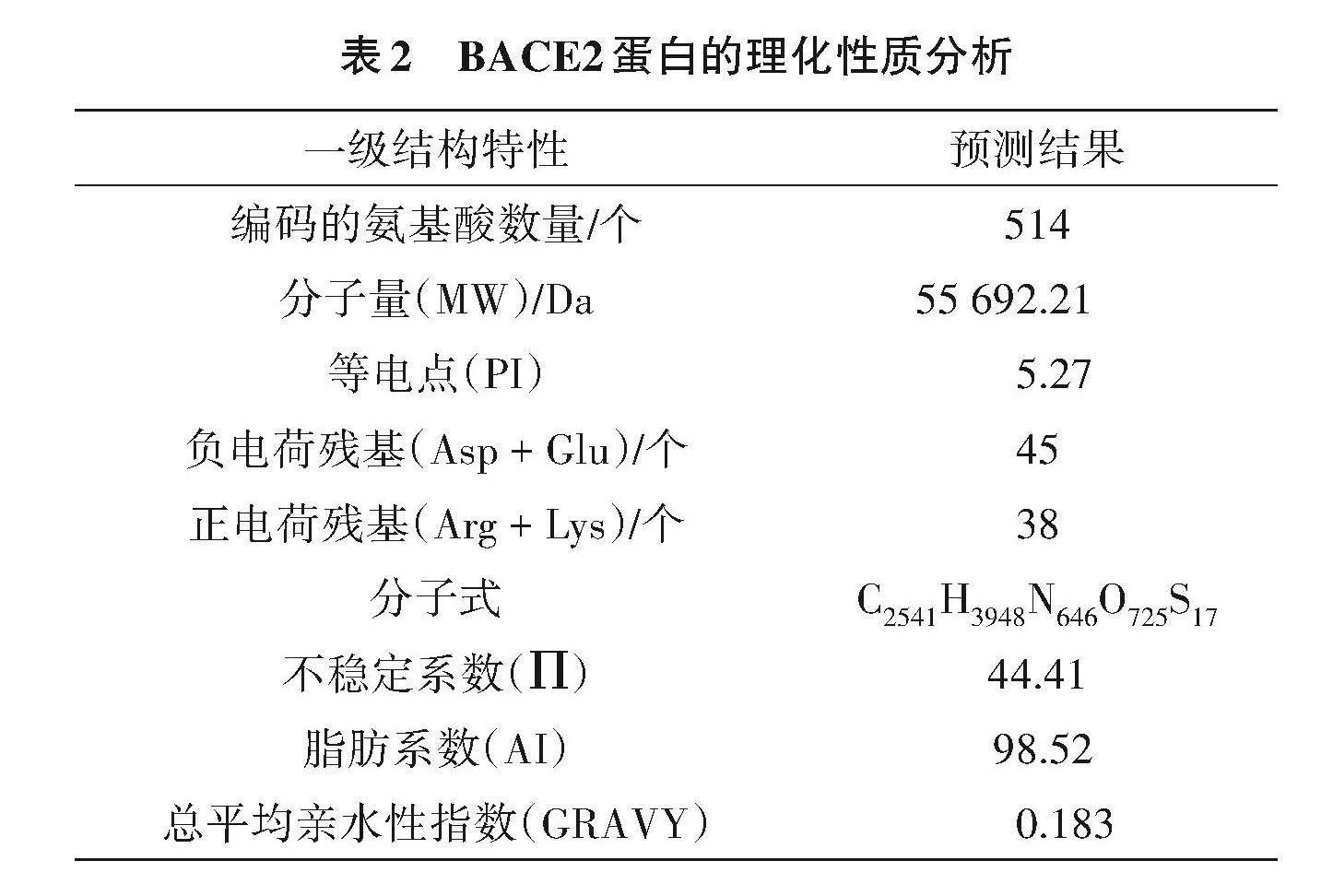
使用 Protparam 软件分析该蛋白理化性质,结果如表2所示,BACE2蛋白的分子式C2541H3948N646O725S17、氨基酸数量为514个,其分子量为55 692.21 Da、理论等电点(PI)5.27,在氨基酸组分中含量最高的是亮氨酸(Leu),占总氨基酸数的11.7%,组氨酸(Cys)含量最低,占总氨基酸数的0.6%。BACE2蛋白带正、负电荷残基总数分别为38、45个,脂肪系数(AI)98.52、不稳定系数()44.41、总平均亲水性指数(GRAVY)0.183。
2.3 BACE2蛋白的信号肽
信号肽的构造可细分为3个主要部分。首先是n区(n-region),是一个富含正电荷的氨基酸序列;紧接着是疏水区(h-region),由9个以上的中性氨基酸组成,共同构建了一个疏水性的界面;最后加工区(c-region),是信号肽酶作用的关键区域,负责切割信号肽,从而完成其生物学功能。
使用了SignaIP工具,对BACE2蛋白的信号肽状态进行预测,结果如图2所示。经过分析,该黑猪BACE2蛋白的信号肽实际上是不存在的,并且未形成跨膜的结构。这一发现有利于更深入地了解该基因编码产物在细胞中的位置和其调节机制。BACE2蛋白可能作为一种酶,直接与细胞质基质中的代谢底物发生相互作用,促进细胞内产生各种化学变化,被归类为非分泌型蛋白质。
2.4 BACE2蛋白的亲疏水性
通过在线分析平台研究BACE2蛋白的疏水特性,结果如图3所示。研究发现,黑猪的BACE2蛋白疏水性氨基酸的数量多于亲水性氨基酸,其中第133号氨基酸的位置展现了最高的疏水性(最小的亲水性);在第481号氨基酸的位置,其疏水性最低(亲水性最高)。表明BACE2蛋白实际上是一种疏水性质的蛋白质。
正值越高,代表疏水性越强;负值越高,代表其亲水性也越强。
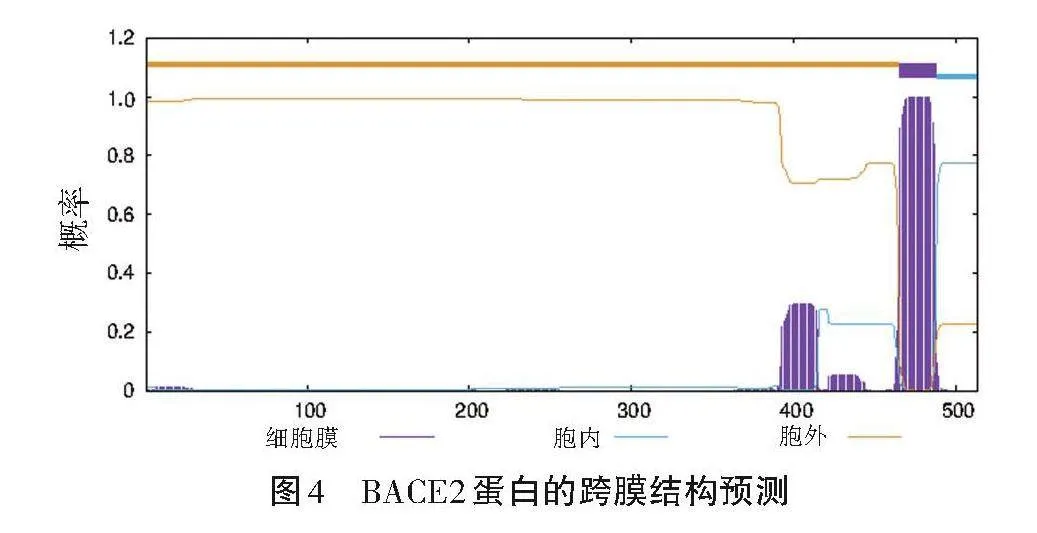
2.5 BACE2蛋白的跨膜结构
运用在线技术平台对BACE2蛋白的跨膜结构进行研究,结果如图4所示。图4横轴呈现了氨基酸在蛋白质中的位置,而纵轴揭示了可能存在的膜上跨越结构。与现存生物材料进行比较,认为该理论模型可精确地解释BACE2蛋白的膜层跨接结构的形成。结果表明,吕梁黑猪体内的BACE2蛋白中有12个区域具备穿透细胞膜的能力。基于这些数据,可以假设该蛋白质具有3个膜间结构。
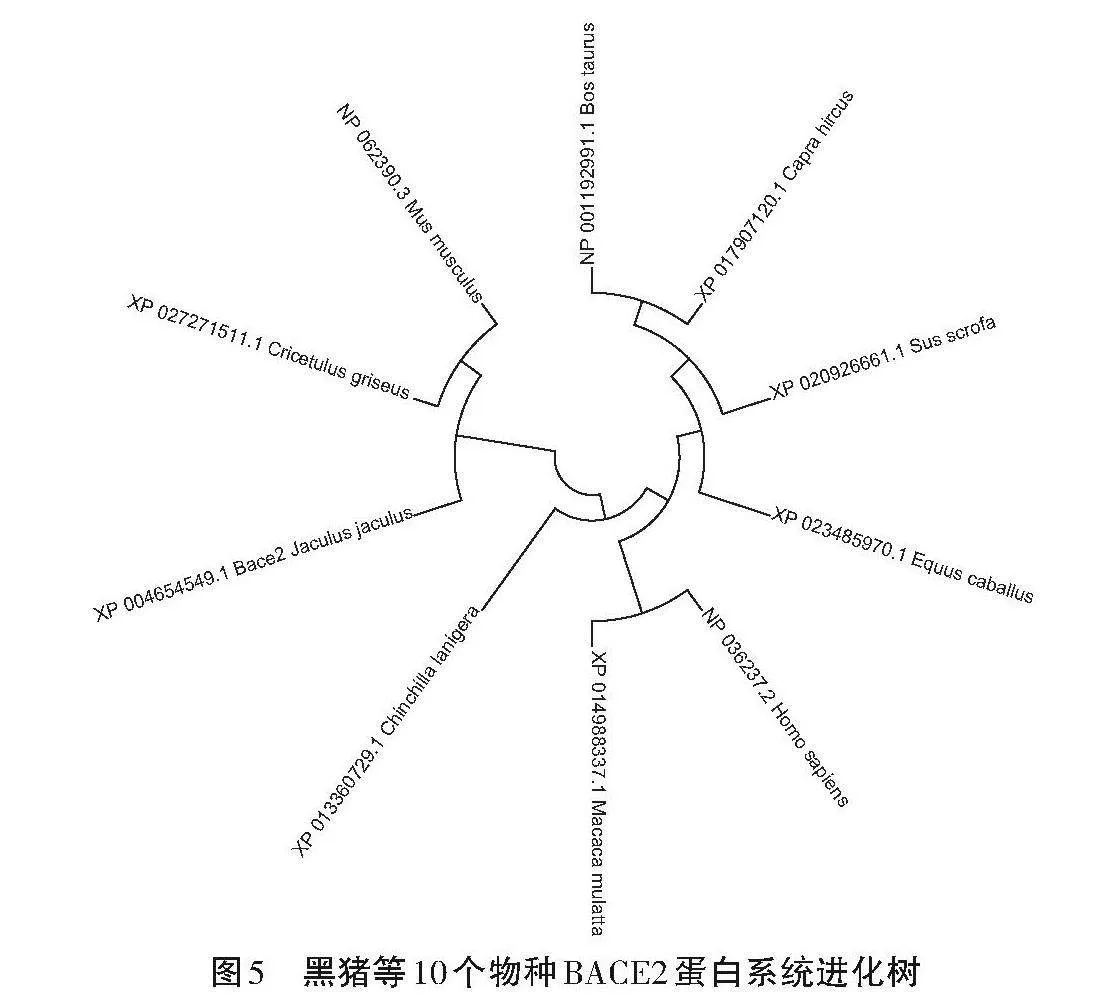
2.6 BACE2蛋白序列比对与进化
使用DNA MAN软件将吕梁黑猪BACE2蛋白的氨基酸序列与牛(NP 001192991.1)、山羊(XP 017907120.1)、马(XP 023485970.1)、智人(NP 036237.2)、猕猴(XP 014988337.1)、长尾龙猫(XP 013360729.1)、小埃及跳鼠(XP 004654549.1)、中国仓鼠(XP 027271511.1)和家鼠(NP 062390.3)的BACE2序列进行比对,绘制系统进化树,如图5所示。吕梁黑猪与山羊和牛的亲缘关系较近,与家鼠的较远。不同物种的BACE2蛋白序列之间存在一定程度的相似性。
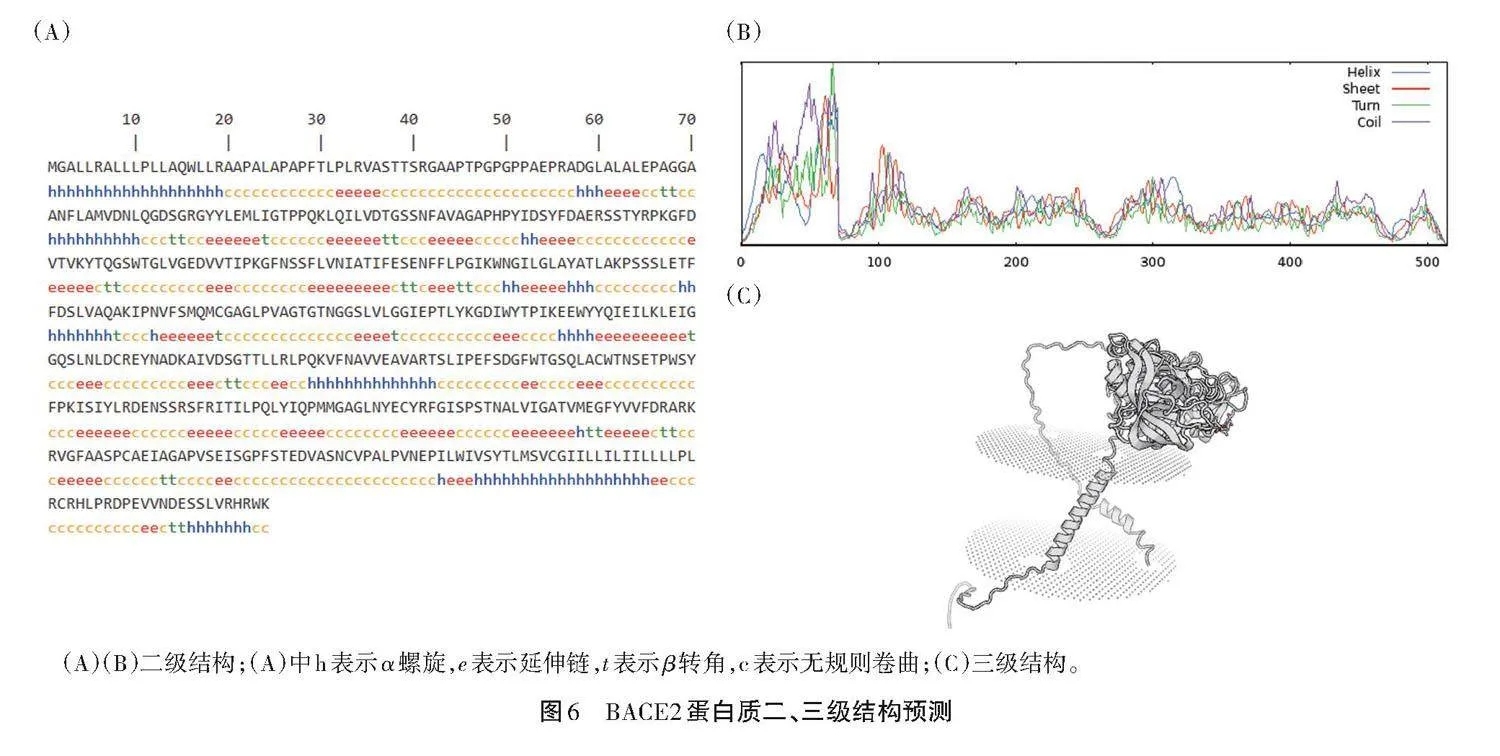
2.7 BACE2蛋白的二级、三级结构预测
蛋白质二级结构作为一级结构和三级结构的纽带,对其预测可为三级结构和功能提供大量信息,如设计蛋白质突变体或确定蛋白质空间结构及功能。使用SWISS-MODEL软件进行BACE2蛋白的二级、三级结构模拟和预测,结果如图6所示。
由图6可知,BACE2蛋白的二级结构主要表现为不规则的卷曲形态,包含252个氨基酸,占总氨基酸的49.03%。α-螺旋结构包含95种氨基酸,占总氨基酸的18.48%。扩展部分包含140种氨基酸,占总氨基酸的27.24%;除此之外,这些也是由不包含肽键和不规则连接点的短链组成的。在β-转角中,存在27种氨基酸,占总氨基酸的5.25%;还存在其他蛋白质中的一些小片段,这些片段呈现了一定的相似性,这些特征表明其是一个具有巨大开发潜力的动物资源库。
在对吕梁黑猪BACE2蛋白的结构特性进行深入研究的过程中,发现其三级结构主要是基于二级结构来构建的,并且通过多肽链进行进一步的盘绕和折叠,最终形成了一种复杂的三维形态(图6C)。在这些元素中,存在许多具有不同构型空间的非共价键连接模式的二级结构单元。这些次级结构单元具有多样的形状,包括α-螺旋、β-折叠、β-转角,以及各种不规则的卷曲感。β-折叠与其他3个类别之间有一定的联系。在次级键的驱动下,这些结构在侧链基团间的相互作用中共同转变为蛋白质的三级结构。由于氨基酸残基间的相互作用力不同,以及受分子内氢和氧形成共价键等各种因素影响,其空间结构呈现明显的异质性。分析结果表明,该蛋白达到了三级结构的标准,但仍然呈现出不规则的卷缩形态,这进一步突显了其结构成分的多样性和高度的复杂性。
2.8 BACE2蛋白亚细胞定位
通过PSORT Ⅱ在线软件分析发现,吕梁黑猪的BACE2蛋白主要分布于内质网(55.6%),同时还分布于高尔基(22.2%)、细胞外(11.1%)、细胞壁和质膜,占比11.1%。
2.9 BACE2蛋白互作网络
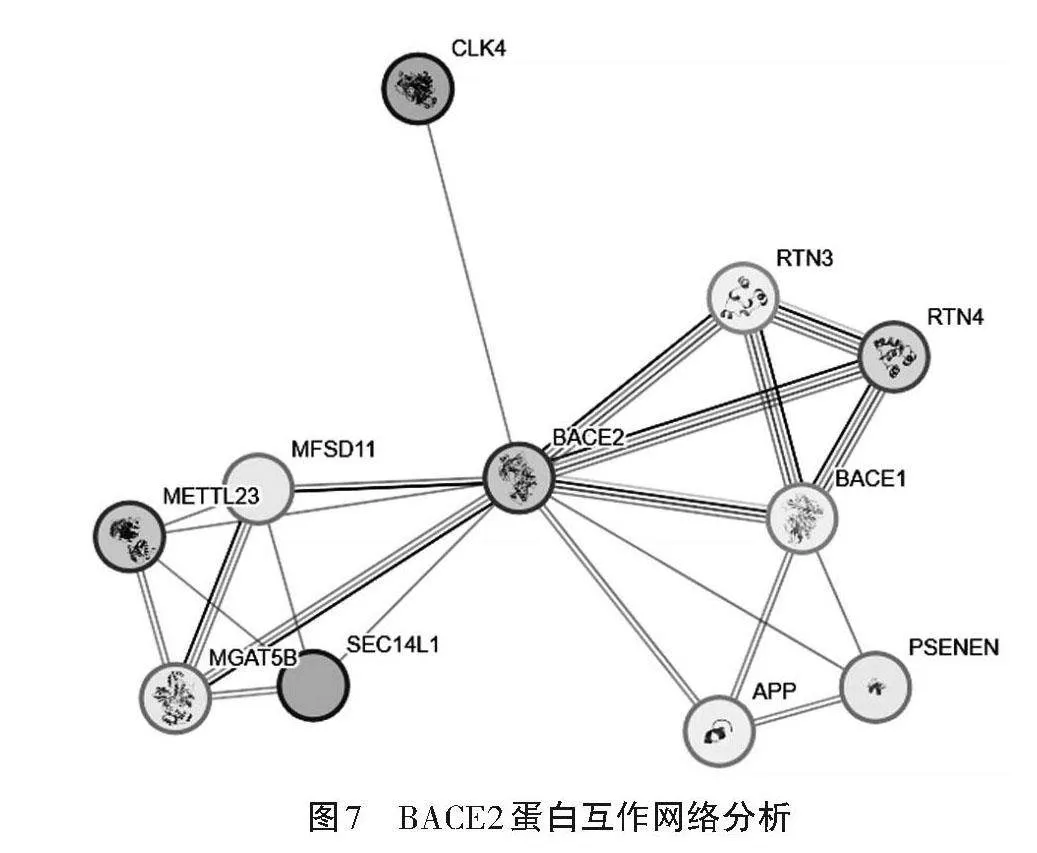
利用在线软件STRING构建BACE2蛋白互作网络,结果如图7所示,BACE2蛋白与CLK4、RTN3、RTN4、AKT3、MFSD11、BACE1、METTL23、PSENEN、MGAT5B和SEC14L1蛋白存在相互作用。网络互作节点11个,边数22个,平均局部聚类系数0.897。
2.10 BACE2基因在不同组织中的表达量
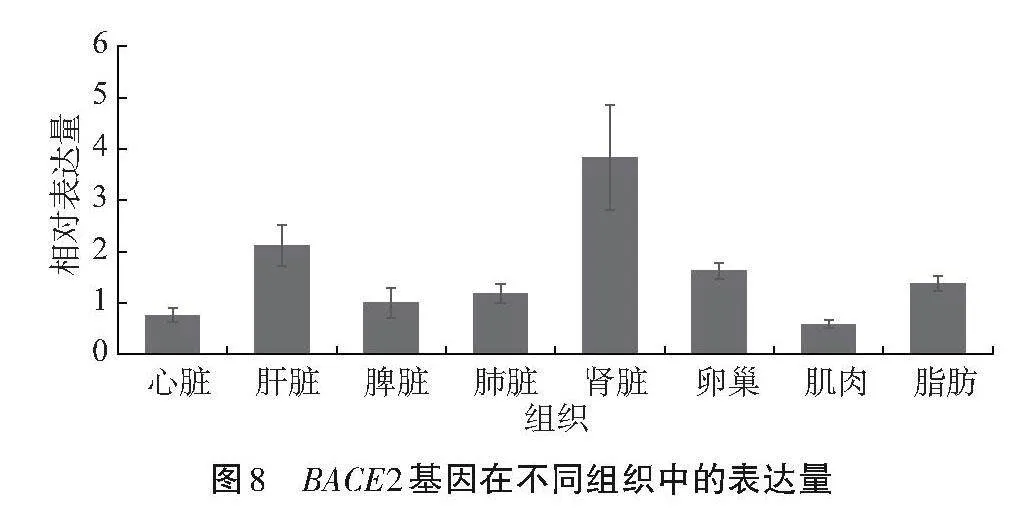
由图8可知,BACE2基因在吕梁黑猪心脏、肝脏、脾脏、肺脏、肾脏、肌肉、卵巢和脂肪组织中均有表达,其中在肾脏中的表达量最高,其次是肝脏,肌肉组织中的表达量最低。
3 结论与讨论
BACE2与BACE1具有同源性,在APP代谢途径中扮演类似的角色,可以水解APP。BACE2的重要功能之一是调节Aβ的生成,而Aβ是AD中神经纤维斑块的主要成分[6]。目前,已有研究证明BACE2在其他生理和病理过程中同样发挥重要作用。Sun等[7]和虞永峰[8]在非小细胞肺癌细胞中敲减BACE2基因,发现其能明显影响癌细胞的迁移、侵袭能力以及骨转移。刘希等[9]研究发现,引入人源性BACE2基因可以降低小鼠脑组织内APP的表达,减少海马区Aβ淀粉样斑块沉积,明显改善了小鼠的学习和记忆能力。鉴于BACE2基因在动物生理和病理过程中的重要作用[10-11] ,本研究在吕梁黑猪中开展了相关研究,成功克隆了黑猪BACE2基因的CDS区序列,全长1 545"bp,编码514个氨基酸。BACE2蛋白是碱性的,缺少信号肽,表明其合成后会直接在核糖体中发挥作用,不需要经历分泌过程,因此被归类为非分泌型蛋白。这类蛋白质不仅展现了出色的疏水性和稳定性,而且还表现出明显的跨膜作用。此外,从蛋白质二级结构看,吕梁黑猪BACE2蛋白以无规则卷曲为主,并伴有α-螺旋及片状结构分散分布。三级结构预测结果对二级结构成分预测精度有进一步验证的作用,两者结论高度吻合。Maloney等[12]和Sáez-valero等[13]研究表明,BACE2蛋白在同系同物种的进化过程中发挥了至关重要的作用。本研究将黑猪BACE2基因序列与牛、山羊等9个物种进行比对,结果表明,吕梁黑猪与山羊和牛的亲缘关系较近,与家鼠的较远,表明哺乳动物的BACE2基因序列相似性较高。
Sáez-valero等[13]分析表明,BACE2"mRNA在人体外周组织中表达水平较低,而在结肠、肾脏、胰腺、胎盘、前列腺、胃和气管中表达水平较高;成人和胎儿的全脑和大多数成人脑亚区表达很低或检测不到,成年大鼠脑原位杂交显示,BACE2"mRNA在大部分脑区以极低水平表达。本研究检测BACE2基因在吕梁黑猪心脏、肝脏、脾脏、肺脏、肾脏、肌肉、卵巢、脂肪组织中均有表达,以在肾脏中的表达量最高,其次是肝脏,肌肉组织中的表达量最低。结合亚细胞定位结果,BACE2蛋白主要分布于内质网上,亚细胞定位结果可能与其作为酶的功能有关。
本研究成功地克隆了黑猪BACE2基因,并对该基因编码的BACE2蛋白使用生物信息学软件进行详细的预测与分析,包括其是否含有信号肽、亲水和疏水属性、膜之间的结构特征,以及蛋白质的二级和三级结构等。结果表明,黑猪BACE2基因的CDS序列长度1 545"bp,编码了514种氨基酸;BACE2蛋白是碱性的,缺少信号肽,展现了出色的疏水性和稳定性,而且还表现出了明显的跨膜作用;其二级和三级结构高度吻合,主要存在与内质网,与多种蛋白具有互作作用;在不同的哺乳物种之间表现出高度的同源性。本研究通过检测不同组织的BACE2基因表达量,发现其在肾脏中的表达量最高,肌肉中的表达量最低。本研究为BACE2蛋白未知生物学功能的研究提供一个新的角度,为了解动物细胞侵袭机制及色素沉着预防与控制奠定基础,为相关方面的研究工作提供参考。未来将综合运用生物信息学研究手段,对黑猪BACE2基因表达机制进行进一步的探索。
参考文献
[1] 周亭,吕月琴. 憨态敦厚的猪[J]. 少儿美术,2023(11):24-27.
[2] 崔海红. 丹麦养猪业生态建设对中国养猪业可持续发展的启示[J]. 世界农业,2020(6):98-103.
[3] WANG Z,XU Q,CAI F,et al. BACE2,a conditional β-secretase,contributes to Alzheimer’s disease pathogenesis[J]. JCI insight,2019,4(1):e123431.
[4] 谢建,刘胜春. BACE2在人体胰腺和胰岛素瘤中的表达[J]. 重庆医科大学学报,2012,37(1):6-9.
[5] 王会志. BACE2激活NF-κB通路促进胶质瘤恶性进展的机制研究[D]. 济南:山东大学,2022.
[6] 王瑞山,张云武,张弦,等. HIF-1调控小鼠bace1基因的转录表达[J]. 厦门大学学报(自然科学版),2007,46(4):564-568.
[7] SUN X L,HE G Q,SONG W H. BACE2,as a novel APP θ-secretase,is not responsible for the pathogenesis of Alzheimer’s disease in Down syndrome[J]. The FASEB journal,2006,20(9):1369-1376.
[8] 虞永峰. BACE2在非小细胞肺癌骨转移中的作用及机制[D]. 上海:上海交通大学,2018.
[9] 刘希,郑娜,卢宏. 人源性BACE2对阿尔茨海默病小鼠海马区淀粉样斑块形成及空间学习记忆能力的影响[J]. 郑州大学学报(医学版),2021,56(2):195-199.
[10] QIU K X,LIANG W P,WANG S,et al. BACE2"degradation is mediated by both the proteasome and lysosome pathways[J]. BMC molecular and cell biology,2020,21(1):13.
[11] DÍAZ-CATALÁN D,ALCARRAZ-VIZÁN G,CASTAÑO C,et al. BACE2"suppression in mice aggravates the adverse metabolic consequences of an obesogenic diet[J]. Molecular metabolism,2021,53:101251.
[12] MALONEY B,GE Y W,GREIG N H,et al. Characterization of the human beta-secretase 2 (BACE2) 5’-flanking region:identification of a 268-bp region as the basal BACE2 promoter[J]. Journal of molecular neuroscience,2006,29(1):81-99.
[13] SÁEZ-VALERO J,PÉREZ-GONZÁLEZ R. BACE2 beyond β-processing of APP,its neuroprotective role in cerebrovascular endothelium[J]. Journal of neurochemistry,2023,166(6):887-890.
(责任编辑:胡立萍)
猜你喜欢
中国中药杂志(2016年22期)2017-02-13 16:47:19
中国校外教育(下旬)(2016年11期)2016-12-27 11:42:42
中国教育信息化·基础教育(2016年10期)2016-12-20 20:09:27
今传媒(2016年11期)2016-12-19 11:35:50
电脑知识与技术(2016年27期)2016-12-15 19:19:25
中国科技纵横(2016年17期)2016-11-30 21:40:27
科技视界(2016年23期)2016-11-04 10:07:53
考试周刊(2016年78期)2016-10-12 11:45:31
课程教育研究·学法教法研究(2016年10期)2016-06-16 16:04:38
安徽农学通报(2015年20期)2015-11-07 01:54:56
