
平茬对鄂托克旗生态脆弱区柠条叶功能性状的影响
2024-12-31 00:00:00刘璐祁伟郭月峰樊霞霞池久威
中国农业科技导报 2024年7期

摘要:为探明平茬对鄂托克旗生态脆弱区柠条叶功能性状的影响,筛选促进柠条恢复生长的最优平茬高度,以内蒙古鄂尔多斯鄂托克旗生态脆弱区的柠条林为研究对象,对柠条进行距地表0(H1)、10(H2)、15(H3)、20 cm(H4)高度平茬处理,以未平茬植株为对照(CK),分析柠条叶功能性状对平茬的响应。结果表明,平茬对叶碳(C)含量无显著影响,对比叶面积(specific leaf area,SLA)、叶组织密度(leaf tissue density,LTD)、叶干物质含量(leaf dry matter content,LDMC)、叶氮(N)含量、C/N、净光合速率(net photosynthetic rate,Pn)、蒸腾速率(transpiration rate,Tr)、气孔导度(stomatal conductance,Gs)均有显著影响,H3的SLA、LTD、LDMC、N、C/N均较CK显著增加。与CK相比,平茬后SLA、N、Pn、Tr、Gs的增幅分别为7.87%~19.46%、3.81%~18.75%、17.71%~40.86%、14.85%~35.30%、13.93%~27.68%;LTD、LDMC、C/N 的降幅分别为0.18%~1.20%、1.51%~4.84%、5.31%~16.82%。不同平茬高度处理的Pn、Tr均表现为“双峰”变化趋势,Gs表现为“单峰”变化趋势。SLA、N、Pn、Tr、Gs之间呈显著正相关,且分别与LTD、LDMC、C/N呈显著负相关。平茬后柠条向“快速投资-收益型”的资源权衡策略移动。不同平茬高度处理的叶功能性状基本表现为H3gt;H2gt;H1gt;H4gt;CK,即15 cm平茬高度处理最优,表明鄂托克旗生态脆弱区柠条应采取15 cm平茬高度处理以提高其生长速率。
关键词:平茬;叶功能性状;柠条;生态脆弱区
doi:10.13304/j.nykjdb.2023.0789
中图分类号:S58 文献标志码:A 文章编号:1008‑0864(2024)07‑0061‑08
柠条锦鸡儿(Caragana korshinskii)又称柠条,是豆科锦鸡儿属的落叶灌木[1],其适应性和抗逆性较强,可改善土壤结构及营养状况、提高土壤保水和持水能力,在受到干旱胁迫时可进行自身水分调节,降低蒸腾作用,并减少水分散失,胁迫解除后,可迅速恢复生长,是中国干旱、半干旱地区水土保持和固沙造林的重要树种[2-4]。近年来,鄂托克旗生态脆弱区环境恶劣,且人工生态林建设存在树种组成单一、林分结构简单、林分密度偏大、抚育措施不当等诸多问题,该区域柠条在生长6~8年后便开始出现生长衰退、生产力下降等现象,因此需对该区人工植被的栽培技术及其适应性进行深入研究,便于柠条林植抚育更新。
平茬是利用植物补偿生长的特性,通过人为去除植物的地上部分,促进植物的生长和发育[5-7]。研究发现,平茬后植株的再生能力明显提高,其新生枝条数、枝长、生物量、叶面积及含水量均呈明显增加趋势,植株的光合速率、蒸腾速率和枝水势也均有所提高[8-10];平茬有利于提高植株对土壤水分的利用效率,更有利于植株的更新复壮和持续发展[11]。然而,这方面的研究目前仅局限于伐桩萌枝方面,尤其是萌枝数量、生长量与平茬高度的关系分析[12‑13],缺乏平茬对叶功能性状影响规律的探讨。功能性状作为植物对环境变化的响应指标,表征植物在逆境条件下的生存和资源利用策略[14],其中叶片功能特性在植物碳同化、水分关系和能量平衡中发挥着重要作用[15],然而,现有的植物叶片功能性状研究主要集中于森林与草原[16‑17],目前对内蒙古自治区鄂尔多斯市鄂托克旗生态脆弱区柠条叶功能性状尚缺乏深入研究。
针对鄂托克旗典型生态脆弱区柠条人工灌木林发育较差、退化严重、生态经济效益较低等瓶颈问题,本研究以该区柠条人工灌木林为研究对象,对柠条林分别进行0、10、15、20 cm高度平茬处理,以未平茬处理为对照,在生长季对不同平茬处理柠条林叶的形态性状、组织化学性状及光合生理性状进行测定,分析柠条林叶功能性状对平茬的响应,以期为内蒙古典型生态脆弱区柠条林高质量可持续发展及地区生态环境建设提供理论依据。
1 材料与方法
1.1 研究区自然概况
研究区位于内蒙古自治区鄂尔多斯市鄂托克旗乌兰镇乌素其日嘎村。该地属典型温带大陆性季风气候,降水少,年均降水量250 mm,且时空分布极为不均,主要集中在7—9月,蒸发量大,年均蒸发量3 000 mm;年均日照时数3 000 h,年均气温6.4 ℃,无霜期122 d。地带性土壤为风沙土及栗钙土。原生植被以克氏针茅(Stipa krylovii)为主,次生植被以油蒿(Artemisia ordosica)、沙竹(Phyllostachys propingue) 、沙蓬(Agriophyllumsquarrosum)为主。主要植物群落有蒿属(Artemisia)、羊柴(Corethrodendron fruticosum)、花棒(Hedysarum scoparium) 、白草(Pennisetumflaccidum)、冰草(Agropyron cristatum)、苦豆子(Sophora alopecuroides)、柠条等。
1.2 试验设计
以内蒙古鄂尔多斯鄂托克旗典型生态脆弱区柠条人工灌木林为研究对象,在试验地选择立地条件、管护措施、生长状况等基本一致的柠条人工灌木林地作为试验样地。并对样地基本情况进行调查,样地坡度小于5°,样地内柠条林的株行距为2 m×4 m。对柠条林地分别采取0(H1)、10(H2)、15(H3)和20 cm(H4)高度的平茬处理,以未平茬处理作为对照(CK)。于2021年3月上旬对试验地的柠条锦鸡儿进行平茬处理,每个处理3次重复,每个重复面积50 m×50 m。
1.3 试验方法
1.3.1 叶形态及组织化学性状的测定
于2022年8月在不同处理样地选取长势基本一致、具有代表性的柠条叶片,每丛柠条采集10片中等大小的健康叶片,至少5丛柠条作为1个样本。将采集的叶片装入有标签的保鲜袋中,于当天放入2~3 ℃冰箱中冷藏。将叶片用去离子水清洗,使用11000XL 扫描仪对叶片进行扫描,并结合使用WinRHIZO Pro软件测定叶面积(leaf area,LA)、叶体积(leaf volume,LV)等形态特征。测定完成后,将每个样品的叶片置于水中,在4 ℃下遮光存储24 h[18],使叶片水分饱和后取出,用吸水纸吸干叶片表面的水分后称取叶片的饱和鲜重(leafsaturated fresh mass,LMSF);再将所有叶片在60 ℃干燥48 h,测定叶片干物质量(leaf dry mass,LMD)[19]。分别计算比叶面积(specific leaf area,SLA)、叶干物质含量(leaf dry matter content,LDMC)、叶组织密度(leaf tissue density,LTD),公式如下。
SLA=LA/LDW (1)
LTD=LMD/LV (2)
LDMC=LMD/LMSF (3)
使用高通量球磨系统将叶片干样粉碎,过0.149 mm筛用于化学特性分析,通过元素分析仪测定碳(C)、氮(N)含量,并计算叶碳氮比(C/N)。
1.3.2 叶光合及水分生理性状的测定
于2022年8月选择天气晴朗日,在各处理样地中分别选择具代表性且长势良好的柠条各5 丛,用GFS-3000 光合作用-荧光测定仪测定植株的生理参数,包括叶片净光合速率(net photosynthetic rate,Pn,μmol·m-2·s-1)、蒸腾速率(transpiration rate,Tr,mmol·m-2·s-1)、气孔导度(stomatal conductance,Gs,mmol·m-2·s-1)。选择植株中上部的健康叶片从7:00 至19:00,每2 h 对叶片进行测定,共记录5次数据,取平均值。
1.4 数据统计与分析
使用SPSS 26.0软件进行数据分析。采用单因素方差分析(one-way ANOVA)统计不同处理柠条叶片性状差异,并采用Fisher 最小显著差异(least significant difference,LSD)进行显著性检验。利用Origin 9.4制图。
2 结果与分析
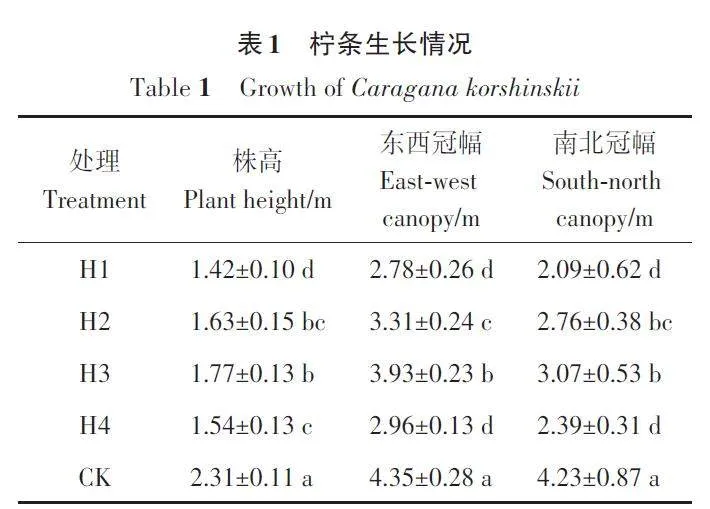
2.1 不同平茬处理柠条生长状况分析
由表1可知,平茬处理的株高和冠幅均显著小于CK。在3个平茬处理中,H3处理的株高和南北冠幅显著高于H1和H4处理,与H2处理差异不显著;H3 处理的东西冠幅显著大于其他平茬处理。
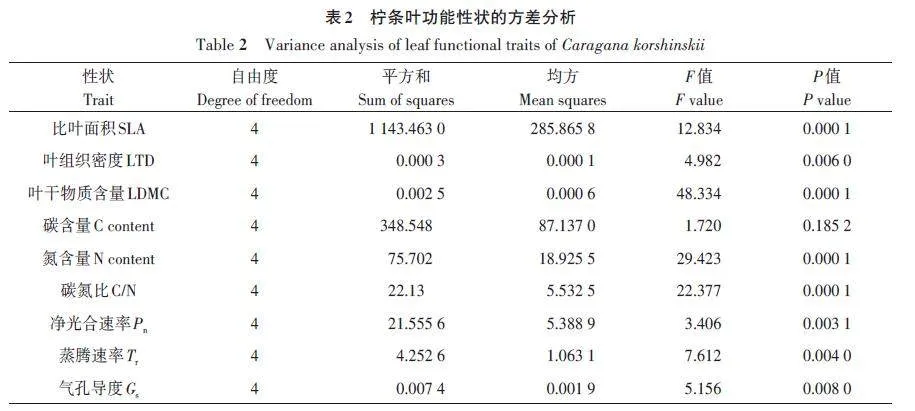
2.2 不同平茬处理柠条的叶功能性状的方差分析
对柠条叶功能性状进行方差分析,结果(表2)表明,不同平茬处理对叶C含量影响不显著,对比叶面积(SLA)、叶组织密度(LTD)、叶干物质含量(LDMC)、N含量、C/N、净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)均有显著影响。
2.3 不同平茬高度对叶形态性状的影响
不同平茬处理柠条叶片的形态性状如图1所示。平茬处理柠条的比叶面积(SLA)均高于CK,其中H1、H2和H3处理显著高于CK。平茬处理柠条的叶组织密度(LTD)、叶干物质含量(LDMC)低于CK,其中H3 处理的LTD 显著低于CK;H1、H2、H3、H4 的LDMC 均显著低于CK。H1、H2、H3、H4 和CK 处理柠条的SLA 分别为119.22、120.30、128.84、116.35 和107.86 cm2·g-1,H1、H2、H3、H4处理分别是CK的1.11、1.12、1.19、1.08倍;而LTD 和LDMC 分别较CK 降低0.18%~1.20% 和1.51%~4.84%。
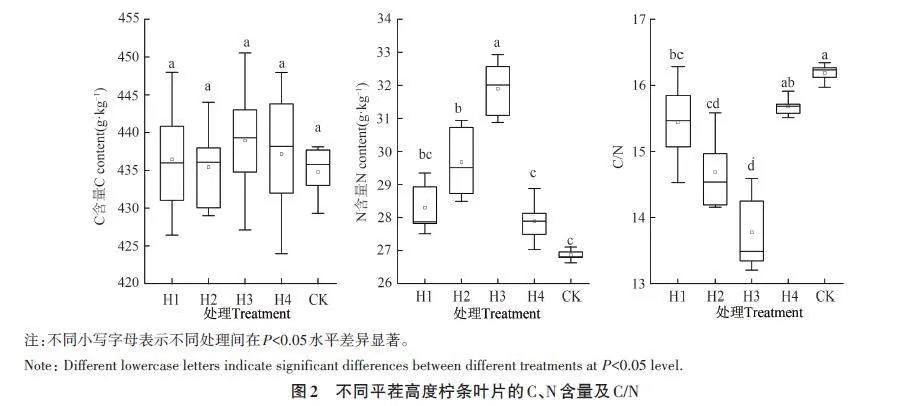
2.4 不同平茬高度对叶组织化学性状的影响
不同平茬处理柠条的叶组织化学性状如图2所示。H1、H2、H3、H4 柠条叶片的C 含量为434.78~438.96 g·kg-1,均高于CK,但差异不显著;叶片N 含量也均高于CK,较CK 增加3.81%~18.75%,其中H2和H3处理显著高于CK,不同平茬处理表现为H3gt;H2gt;H1gt;H4;平茬处理叶片的C/N均低于CK,较CK降低5.31%~16.82%,不同平茬处理表现为H3
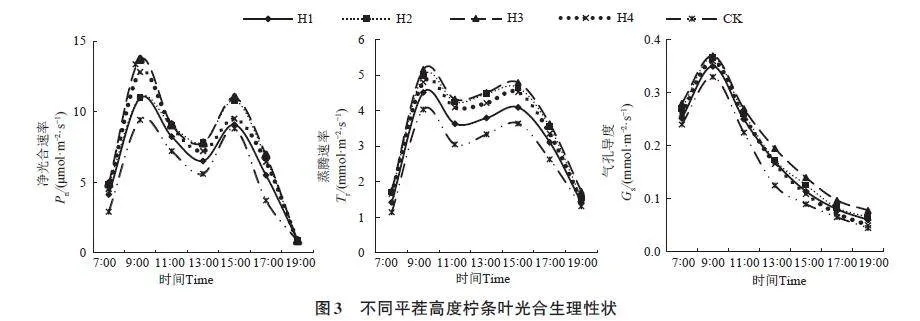
2.5 不同平茬高度对叶光合生理性状的影响
不同平茬处理柠条的叶光合生理性状如图3所示。各处理的净光合速率(Pn)和蒸腾速率(Tr)均表现为“双峰”变化趋势,7:00—9:00呈上升趋势,于9:00达到第1次峰值,且为全天最大值,之后随光合有效辐射上升的同时,气温升高、空气湿度降低,Pn及Tr均呈下降趋势,Pn在13:00左右、Tr在11:00左右出现低谷,此时为“热休眠”现象;然后又逐渐升高,在15:00左右出现第2次峰值,之后又呈下降趋势。各处理的气孔导度(Gs)均表现为“单峰”变化趋势,7:00—9:00 呈上升趋势,于9:00左右达到峰值,之后呈持续下降趋势。不同平茬处理的Pn、Tr、Gs均高于CK,其中H1、H2、H3、H4的Pn较CK分别增加17.71%、32.51%、40.86%、30.52%; Tr 较CK 分别增加14.85%、31.36%、35.30%、25.88%;Gs 较CK 分别增加13.93%、20.09%、27.68%、13.57%。
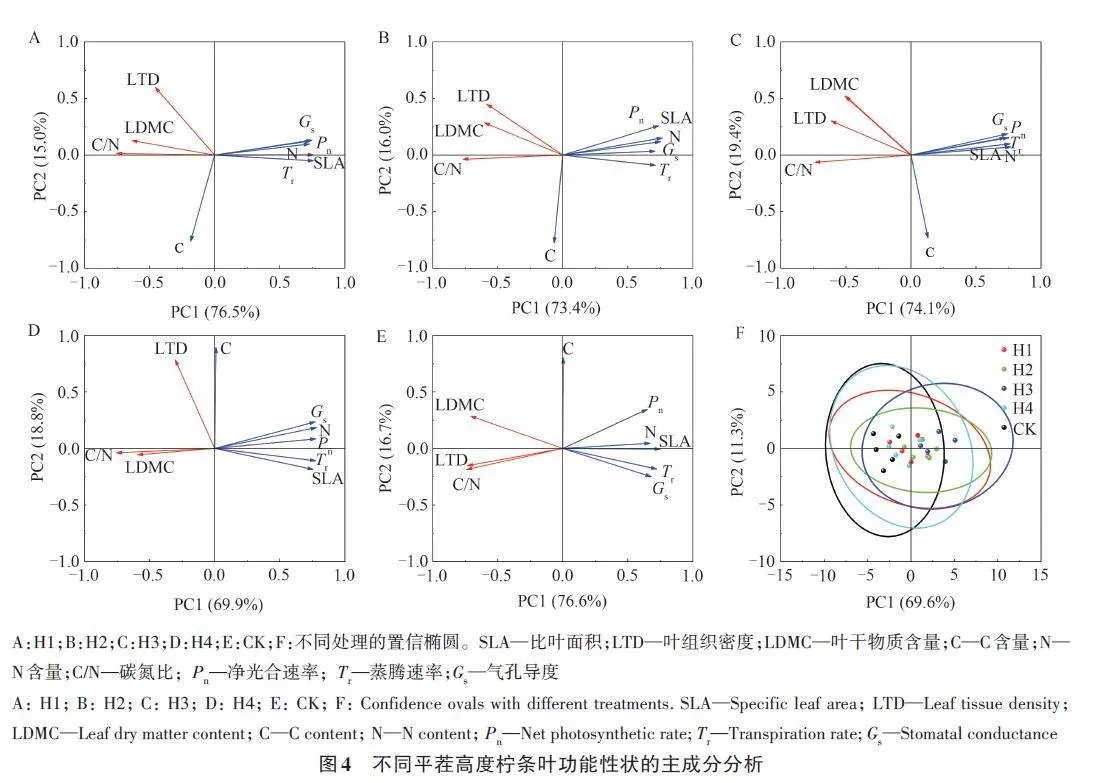
2.6 不同平茬高度对叶性状及其相互关系的影响
不同平茬高度柠条叶形态性状、组织化学性状及光合生理性状的主成分分析结果(图4)表明,除叶片C含量与其他性状无显著关系外,其余性状间均存在显著的相关关系。其中,SLA、N、Pn、Tr、Gs 之间呈显著正相关,且分别与LTD、LDMC、C/N呈显著负相关。2个主成分的累计贡献率达80%以上,能较好地反映叶片功能性状之间的关系。CK的PC1轴主要是由SLA、N、LTD定义的结构轴,PC2 轴由C 定义;CK 的SLA 和N、LTD 和C/N、Tr 和Gs 的关系最为紧密。平茬后,H1、H2、H3、H4的PC1轴由C/N和Tr定义,PC2轴变化较小,SLA与LTD和LDMC的相关性减弱,与N、C/N、Pn、Tr、Gs的相关性增强。不同平茬高度的置信分组虽然有一些重叠,但各处理叶性状是聚集的。CK的叶性状主要分布在PC2轴的左半部,而H1、H2、H3、H4的的叶性状沿PC2轴从左到右移动,其中H3主要分布在PCA2轴的右半部。
3 讨论
本研究发现,相比未平茬处理,经平茬处理后柠条的比叶面积显著增大,而叶组织密度、叶干物质含量显著降低,这与前人研究结果一致[20]。在资源丰富的环境中往往具有较高的比叶面积,当植物受养分限制或种间竞争时,通过增加比根长、根比表面积可提高对养分的获取能力或竞争力;叶组织密度和干物质含量的降低能够加快植物生长,使植物减少水分和养分的流失,从而提高水分和养分的利用效率,增强防御力[21]。在平茬后的补偿性恢复生长过程中,改变了柠条的资源获取条件,降低了柠条叶片的构建成本,从而减小了植物的蒸腾散失,促使柠条调整适应策略,以提高生存能力,为植物的生长代谢提供水分和养分,形成高比叶面积、低组织密度及低干物质含量的生长策略[22]。
植物组织的基本化学营养元素在植物生长发育和各种生理生化调节机制中发挥着重要作用,是研究植物响应与适应策略的重要指标。植物叶片N是光合物质代谢和植物生长的关键性因子,是合成叶绿素和有关光合蛋白的重要成分,叶N含量高常具有较高的光合速率[23‑24]。C/N代表着植物吸收营养元素时所能同化C的能力,低C/N表征植物具有较快的生长速率[25]。本研究表明,不同平茬高度处理对柠条叶N含量和C/N均有显著影响,但叶C含量在不同平茬高度处理间无显著差异,且叶C的稳定性较高,这是因为植物器官内有机碳一般不直接参与植物生产活动,而主要在植物体内起骨架的作用,为植物活动提供能源,因此变异较小[26]。不同平茬处理柠条叶N含量较CK显著增加,这是由于平茬处理后植物根系的新陈代谢增强,提高了对土壤N等营养元素的吸收与运输能力。此外叶面积是衡量叶片光合能力的指标,叶面积越大,制造的有机物越多[27]。本研究表明,平茬后的叶面积均显著大于CK。平茬后,叶面积的增加和氮运输能力的增强,共同提升了植物的光合作用效率,促进叶片叶绿素和光合蛋白的合成,显著提高了叶片N含量,使植物快速恢复生长。本研究表明,平茬处理叶的C/N均显著低于CK,其中15 cm平茬处理的叶C/N最小,表明15 cm平茬处理下柠条具有更高的生长速率。
平茬可改变植物叶片光合生理性状,其中光合作用的升高是植物出现超补偿生长的基础生理活动。本研究发现,平茬处理柠条的叶Pn日均值显著高于CK,这与前人研究结果一致。周晓红等[28]等通过研究刈割后黑麦草发现,其光合作用显著升高;董雪[29]发现,平茬后沙冬青的Pn高于未平茬处理。萌蘖植物经过平茬后,新生枝叶的分生组织活动强烈,细胞分裂速度较快,需要消耗大量的同化产物,而这一需求只能通过旺盛的呼吸作用来满足[30],因此导致平茬处理柠条的Tr相对较高。光合作用的显著增高与Gs密切相关,本研究发现,平茬处理后柠条叶片的Gs显著增大,CO2供应得到了保证,进一步增加了柠条的Pn及水分利用效率[31]。本研究还发现,SLA、叶N含量均表现为H3gt;H2gt;H1gt;H4gt;CK。LTD、LDMW、叶C/N则与之相反,表现为H3
植物的功能性状并非相互独立,而是通过协同作用或相互权衡促进植物生长。本研究发现,SLA、N、Pn、Tr、Gs 之间呈显著正相关,且分别与LTD、LDMC、C/N呈显著负相关,与前人研究结果一致[32];经平茬处理后,SLA与LTD、LDMC的相关性减弱,与N、C/N、Pn、Tr、Gs的相关性增强。由此表明,平茬后叶片形态结构、生长模式、养分元素分配策略的调整会改变性状间的相关性。平茬处理后,植物为快速恢复其地上枝叶的缺失会提高根系的养分吸收能力,并增强向叶片运输养分的能力[33]。叶片形态性状与养分吸收之间的联系是直接且紧密的[34],这可能是平茬后叶形态性状与化学组织性状相关性增强的原因,而形态性状间的变化可能是由于叶片化学组织性状对某一形态性状影响后所发生的间接作用,因此在平茬的补偿恢复生长过程中叶片形态性状间的相关性相对减弱。
根据叶经济谱论(leaf economic spectrum,LES),植物功能性状如SLA、LTD、LDMW、N含量、C/N及Pn等是衡量植物环境资源权衡策略的重要指标[35]。具体而言,具有高SLA的物种表现出高叶N含量、高光合速率和短叶片寿命以及低LTD,即资源获取策略,植物通常具有较高的光合速率和生长速率;相反,SLA较低的物种表现出保守特性[36]。本研究表明,与未平茬柠条相比,平茬后柠条的LTD、LDMW显著降低,SLA显著增大,同时叶片N含量及叶Pn较高,表明平茬后柠条在有限的资源环境中通过权衡策略在叶片功能性状之间进行了资源的优化配置;平茬后的柠条较未平茬柠条在叶经济谱中更靠近“快速投资-收益”型物种的一端。本研究结果对鄂托克旗柠条的恢复生长及生态管理工作具有重要意义。
参考文献
[1] 于晓燕,汪星,吕雯,等.黄土丘陵区带状柠条锦鸡儿林地深层土壤干化及根系分布[J].干旱区地理,2023,46(5):753-762.
YU X Y, WANG X, LYU W, et al .. Deep soil desiccation androot distribution of belted Caragana korshinskii forest in loesshilly region [J]. Arid Land Geogr., 2023, 46(5):753-762.
[2] 张力斌,何明珠,张珂.柠条锦鸡儿生物量分配规律与异速生长对氮、磷添加的响应[J].生态学报,2023,43(16):6627-6636.
ZHANG L B, HE M Z, ZHANG K. Response of biomass allocationand allometric growth of Caragana korshinsküi to nitrogen andphosphorus addition [J]. Acta Ecol. Sin., 2023,43(16):6627-6636.
[3] 刘婧,缑倩倩,王国华,等.晋西北丘陵风沙区柠条锦鸡儿叶片与土壤生态化学计量特征[J].植物生态学报,2023,47(4):546-558.
LIU J, HOU Q Q, WANG G H, et al .. Leaf and soil ecologicalstoichiometry of Caragana korshinskii in windy and sandy hillyregion of northwest Shanxi, China [J]. Chin. J. Plant Ecol.,2023, 47(4):546-558.
[4] 杜雨佳,赵勇钢,刘小芳,等.黄土丘陵区柠条种植带状结构对坡面土壤水分的影响[J].水土保持研究,2020,27(6):52-59.
DU Y J, ZHAO Y G, LIU X F, et al .. Effects of strip Caraganakorshinskii plantation on soil moisture at hillslope scale in theLoess Hilly region [J]. Res. Soil Water Conserv., 2020, 7(6):52-59.
[5] 王震.不同留茬高度对四合木生长及生理生化特性的影响研究[D].北京:中国林业科学研究院,2013.
WANG Z. Effects of different stubble height on growth andphysiologicalbiochemical characteristics of Tetraena mongolicaMaxim [D]. Beijing: Chinese Academy of Forestry Sciences, 2013.
[6] WANG X, QI W, GUO Y F, et al .. Effects of stumping on fineroot architecture, growth, and physiology of Hippophaerhamnoides [J/OL]. PeerJ, 2023, 11: e14978 [2023-09-30].https://doi.org/10.7717/peerj.14978.
[7] 刘晓宇,郭月峰,姚云峰,等.砒砂岩区不同留茬高度及坡向下沙棘根系分形特征[J].生态环境学报,2021,30(1): 100-107.
LIU X Y, GUO Y F, YAO Y F, et al.. Fractal features of Hippophaerhamnoides roots under different stubble height and slopes in softsandstone area [J]. Ecol. Environ. Sci., 2021, 30(1): 100-107.
[8] 王丹,李熙颜,颜廷雨,等.不同季节平茬对云南松生物量分配与异速生长的影响[J].西南农业学报,2023,36(1): 47-52.
WANG D, LI X Y, YAN T Y, et al.. Effects of flat stubble indifferent seasons on biomass allocation and allometric growth ofPinus yunnanensis [J]. Southwest China J. Agric. Sci., 2023, 36(1):47-52.
[9] 王震,张利文,虞毅,等.平茬高度对四合木生长及生理特性的影响[J].生态学报,2013,33(22): 7078-7087.
WANG Z, ZHANG L W, YU Y, et al .. Effect of differentstubble height treatments on the annual growth index andphysiological characteristics of Tetraena mongolica in twogrowing seasons [J]. Acta Ecol. Sin., 2013,33(22):7078-7087.
[10] HANDAVU F. The influence of stump diameter and height oncoppicing ability of selected key Miombo woodland tree species ofZambia: a guide for harvesting for charcoal production [J]. J. Ecol.Nat. Environ., 2011, 3(14):461-468.
[11] 胡小龙,薛博,袁立敏,等.科尔沁沙地人工黄柳林平茬复壮技术研究[J].干旱区资源与环境,2012,26(5):135-139.
HU X L, XUE B, YUAN L M, et al .. Techniques of stumpingand rejuvenating of Salix gordejevii plantation in Kerqin sandyland [J]. J. Arid Land Res. Environ., 2012, 26(5):135-139.
[12] 姚建成,梁海荣,张松林,等.沙柳不同平茬高度对比试验[J].内蒙古林业科技,2009,35(4):35-36.
YAO J C, LIANG H R, ZHANG S L, et al .. Comparison ondifferent stubble height after stumping for Salix psammophila [J]. J.Inner Mongolia For. Sci. Technol., 2009, 35(4):35-36.
[13] 赵菊英,武志博,谢宗才,等.平茬高度对柽柳萌条生长影响的研究[J].内蒙古林业科技,2013,39(1):27-30.
ZHAO J Y, WU Z B, XIE Z C, et al .. Influence of stumpingheight on growth of sprout of Tamarix chinensis [J]. J. InnerMongolia For. Sci. Technol., 2013, 39(1):27-30.
[14] REICH P, WRIGHT I J, CAVENDER-BARES J, et al .. Theevolution of plant functional variation: traits, spectra, andstrategies [J]. Int. J. Plant Sci., 2003, 164:143-164.
[15] ACKERLY D D, KNIGHT C A, WEISS B, et al .. Leaf size,specific leaf area and microhabitat distribution of chaparral woodyplants: contrasting patterns in species level and community levelanalyses [J]. Oecologia, 2002, 130(3):449-457.
[16] CUI E Q, LU R L, XU X N, et al .. Soil phosphorus drives planttrait variations in a mature subtropical forest [J]. GlobalChange Biol., 2002, 28:3310-3320.
[17] HOSSEINI A, HOSSEINI S M, LINARES J C. Linkingmorphological and ecophysiological leaf traits to canopydieback in Persian oak trees from central Zagros [J]. J. For.Res., 2019, 30:1755-1764.
[18] NING Z, LI Y, ZHAO X, et al .. Comparison of leaf and fineroot traits between annuals and perennials, implicating themechanism of species changes in desertified grasslands [J/OL].Front. Plant Sci., 2022, 12:778547 [2023-09-30]. https://doi.org/10.3389/fpls.2021.778547.
[19] JIANG X, JIA X, GAO S, et al .. Plant nutrient contents ratherthan physical traits are coordinated between leaves and roots ina desert shrubland [J/OL]. Front. Plant Sci., 2021, 12:734775[2023-09-30]. https://doi.org/10.3389/fpls.2021.734775.
[20] YANG J, CI X Q, LU M M, et al.. Functional traits of tree specieswith phylogenetic signal covary with environmental niches in twolarge forest dynamics plots [J]. J. Plant Ecol., 2014, 7:110-125.
[21] LAUGHLIN D C, MOMMER L, SABATINI F M, et al .. Roottraits explain plant species distributions along climaticgradients yet challenge the nature of ecological trade-offs [J].Nat. Ecol. Evol., 2021, 5:1-12.
[22] YUAN J H, LI H Y, YANG Y F. The compensatory tillering inthe forage grass Hordeum brevisubulatum after simulatedgrazing of different severity [J/OL]. Front. Plant Sci., 2020, 11:792 [2023-09-30]. https://doi.org/10.3389/fpls.2020.00792.
[23] XU B, XU W, WANG Z. Accumulation of N and P in thelegume Lespedeza davurica in controlled mixtures with the grassBothriochloa ischaemum under varying water and fertilizationconditions [J/OL]. Front. Plant Sci., 2018, 9: 165 [2023-09-30].https://doi.org/10.3389/fpls.2018.00165.
[24] MORALES J, SQUEO F A, TRACOL Y, et al .. Resourceeconomics and coordination among above-and below-groundfunctional traits of three dominant shrubs from the Chileancoastal desert [J]. J. Plant Ecol., 2015, 8:70-78.
[25] HU M Y, MA Z L, CHEN H Y H. Intensive plantationsdecouple fine root C: N: P in subtropical forests [J/OL]. For.Ecol. Manage., 2022, 505:119901 [2023-09-30]. https://doi.org/10.1016/j.foreco.2021.119901.
[26] WANG X X, LI H B, CHU Q, et al .. Mycorrhizal impacts onroot trait plasticity of six maize varieties along a phosphorussupply gradient [J]. Plant Soil, 2022, 448:71-86.
[27] GU J. Effects of root diameter, branch order, soil depth andseason of birth on fine root life span in five temperate treespecies [J]. Eur. J. For. Res., 2017, 136(4):1-12.
[28] 周晓红,王国祥,杨飞,等.刈割对生态浮床植物黑麦草光合作用及其对氮磷等净化效果的影响[J]. 环境科学,2008,29(12):3393-3399.
ZHOU X H, WANG G X, YANG F, et al .. Effects of cutting onphotosynthesis and purification efficiencies on nitrogen andphosphorus of the Lolium multiflorum [J]. Environ. Sci., 2008,29(12): 3393-3399.
[29] 董雪.沙冬青平茬技术及刈割后生理生化特性研究[D].呼和浩特:内蒙古农业大学,2013.
DONG X. Studies on stumping technology of Ammopiptanthusmongolicus and physio-biochemical characteristic to clipping [D].Hohhot: Inner Mongolia Agricultural University, 2013.
[30] GAO Y B,REN A Z,WANG W,et al .. A comparative study onphotosynthesis and morphological characteristics of Salix gordejeviibetween regenerated shoots and standing shoots [J]. Acta Ecol. Sin.,2002, 22(10):1758-1764.
[31] 解婷婷,张希明,梁少民,等.不同灌溉量对塔克拉玛干沙漠腹地梭梭水分生理特性的影响[J]. 应用生态学报,2008,19(4):711-716.
XIE T T, ZHANG X M, LIANG S M, et al .. Effects of differentirrigation rates on water physiological characteristics ofHaloxylon in the hinterland of Taklimakan desert [J]. J. Appl.Ecol., 2008,19(4):711-716.
[32] MARECHAUX I, SAINT-ANDRE L, BARTLETT M K, et al ..Leaf drought tolerance cannot be inferred from classic leaftraits in a tropical rainforest [J]. J. Ecol., 2020, 108: 1030-1045.
[33] YANG Z P, MINGGAGUD H, BAOYIN T G T, et al .. Plantproduction decreases whereas nutrients concentration increases inresponse to the decrease of mowing stubble height [J/OL]. J.Environ. Manage., 2020, 253:109745 [2023-09-30]. https://doi.org/10.1016/j.jenvman.2019.109745.
[34] WRIGHT I J, REICH P B, WESTOBY M, et al.. The worldwideleaf economics spectrum [J]. Nature, 2004, 428: 821-827.
[35] CHEMG J, CHU P, CHEN D, et al .. Functional correlationsbetween specific leaf area and specific root length along aregional environmental gradient in Inner Mongolia grasslands [J].Funct. Ecol., 2016, 30: 985-997.
[36] 肖以华,付志高,许涵,等.城市化对珠江三角洲不同功能群植物叶片功能性状的影响[J]. 生态环境学报,2022,31(9):1783-1793.
XIAO Y H, FU Z G, XU H, et al .. Effects of urbanization onleaf functional traits of different functional groups in PearlRiver Delta [J]. Ecol. Environ. Sci., 2022, 31(9):1783-1793.
基金项目:内蒙古自治区自然科学基金项目(2022MS03029);河套学院高层次人才引进科研启动项目(HYRC202302);内蒙古自治区直属高校基本科研业务费项目(BR22-15-01);内蒙古自治区科技计划项目(2021GG0085);内蒙古自治区教育科学研究“十四五”规划课题项目(NGJGH2023054)。
猜你喜欢
现代农村科技(2021年5期)2021-12-07 16:08:41
乡村科技(2021年16期)2021-10-21 04:02:26
山西果树(2021年2期)2021-06-29 12:48:20
中国煤炭工业(2020年1期)2020-12-28 07:02:52
农家科技中旬版(2019年8期)2019-09-19 04:26:07
当代陕西(2019年7期)2019-04-25 00:22:52
江苏农业科学(2017年4期)2017-05-08 12:13:23
大陆桥视野·下(2016年11期)2017-02-28 20:29:51
农业工程技术·综合版(2016年3期)2016-07-27 07:42:12
现代农村科技(2016年6期)2016-03-28 22:55:01
