
蓝莓自花和异花授粉花柱转录组SSR位点分析
2024-12-31 00:00:00杨芩付燕邓洁张婷渟刘雅兰
安徽农学通报 2024年11期

摘要 为鉴定蓝莓自交不亲和基因,本研究对“杰兔”蓝莓品种自花和异花授粉花柱进行了转录组测序,对简单重复序列(SSR)位点进行分析,对含SSR位点且差异表达的非冗余序列(Unigene)进行基因本体论(GO)功能富集和京都基因与基因组数据库(KEGG)注释分析,并开展SSR标见引物设计。结果表明,蓝莓花柱转录组共获得85 890条Unigene,其中有21 962个Unigene含有26 557个SSR位点,发生频率为25.57%。在所有SSR位点中包含有297种重复基元,以单核苷酸和二核苷酸重复为主要重复类型,占总SSR的89.52%;其中A/T类型和AG/CT类型SSR位点数占SSR总数的83.44%。针对所有SSR位点,共设计出17 212对引物,成功率78.37%。差异转录组筛选到58、27、16、3、2和1条含有SSR位点的差异表达Unigene,分别与植物激素信号转导、细胞骨架、蛋白质泛素化、花粉管生长、核糖核酸酶T2家族和细胞程序化死亡等代谢途径相关。本研究丰富了蓝莓的EST-SSR分子标记,为蓝莓自交不亲和的分子机制研究提供了参考。
关键词 蓝莓;自花授粉;异花授粉;花柱;转录组;简单重复序列
中图分类号 S603.2" "文献标识码 A
文章编号 1007-7731(2024)11-0033-09
Analysis of SSR loci in blueberry self-pollinated and cross-pollinated styles transcriptome
YANG Qin1,2" " FU Yan3" " DENG Jie1,2" " ZHANG Tingting1,2" " LIU Yalan1,2
(1Key Laboratory of Molecular Breeding and Variety Creation of Horticultural Plants for Mountain Featuresnbsp;in Guizhou Province, Kaili 556000, China;"2Development Planning Division, Kaili University, Kaili 556000, China;3Qiandongnan Vocational and Technical College, Kaili 556000, China)
Abstract In order to carry out the identification of self-incompatibility genes in blueberry, transcriptome sequencing was performed on the self-pollinated and cross-pollinated styles of blueberry cultivar “Premier” in this study, and simple repeat sequence (SSR) loci were analyzed. On this basis, SSR loci of assembled and redundant Unigene were analyzed, as well as it was conducted that the significant enrichment analysis of GO and KEGG for the differential expression of unigenes containing SSR, and the SSR primers were designed. The results showed that a total of 85 890 unigenes were obtained from the blueberry styles transcriptome, of that 21 962 Unigene were found to contain 26 557 SSR loci, with a frequency of 25.57%. The mononucleotide and dinucleotide repeats were the main repeat types of the 297 kinds of repeat motif in all SSR loci, accounting for 89.52% of the total SSR, of which the number of A/T and AG/CT SSR locations accounted for 83.44% of the total number of SSR locations. A total of 17 212 pairs of primers were designed for all SSR loci, with a success rate of 78.37%. The differential transcripome combined with SSR loci information showed that 58, 27, 16, 3, 2 and 1 unigene with SSR sites were screened, which were related to plant hormone signal transduction, cytoskeleton, protein ubiquitination, pollen tube growth, ribonucnase T2 family, and programmed cell death, respectively. This study not only enriched EST-SSR markers of blueberry, but also laid a foundation for molecular mechanism of self-incompatibility in blueberry.
Keywords blueberry; self-pollination; cross-pollination; styles;transcriptome; simple sequence repeat
简单重复序列(Simple sequence repeat,SSR)是指1~6个核苷酸为基本单元经多次重复而组成的一段核苷酸序列,其广泛分布于真核生物和原核生物基因组中[1]。与其他分子标记相比,该序列为共显性标记,具有数量丰富,等位基因变异较大,共显性、特异性、多态性和稳定性高,重复性好、覆盖面广等优点[1-2]。目前,SSR分子标记被广泛应用于果树植物的亲缘关系鉴定[3]、遗传多样性分析[4-5]、亲本分析和分子标记辅助育种[6-7]以及特异性状鉴定[8-9]中。随着生物技术的发展,SSR根据来源可分为基因组SSR和表达序列标签SSR(EST-SSR)[10],与基因组SSR相比,EST-SSR标记对分子标记辅助选择更有价值,其与果实大小、果实形状和果皮颜色等农艺性状基因座相关[11]。近年来,相关学者研究分析了薄壳山核桃雌雄花序[1]、黑莓果实[2]、梨果皮[11]、柿雌雄花芽[12]、黑果枸杞经干旱胁迫的叶和根[13]以及枇杷花[14]等转录组数据的SSR特征,并在此基础上开发了SSR引物,为这些果树的遗传多样性分析、种质鉴定、QTL定位和分子标记辅助育种等工作提供了丰富的EST-SSR标记。
蓝莓是一种多年生常绿或落叶灌木类经济小浆果作物[15-16],蓝莓果实酸甜适度、口感细腻,富含花色苷、维生素、氨基酸和多酚等活性成分,具有抗氧化、调节肠道菌群和增强视力等功能[17]。蓝莓被认为是优化人类膳食结构的重要鲜食果品之一,也是较具发展潜力的水果之一[18]。相关研究表明,高丛蓝莓、兔眼蓝莓和矮丛蓝莓三大栽培类型[19]的多数品种自花授粉结实率较低,表现出一定的自交不亲和性[20-23]。目前,关于蓝莓自交不亲和的研究主要集中在其自交不亲和的表型观察与评价上,如花粉在柱头上的水合萌发,花粉管在花柱中的生长情况[22],坐果率与品质差异等[20,24],而关于其形成的分子机制研究较少。SSR分子标记可以有效地鉴定枇杷[25]、红树莓[9]以及油橄榄[26]等的自交不亲和性及S等位基因。目前关于蓝莓EST-SSR分子标记的开发较为有限,部分学者利用蓝莓花芽[27]、叶片[28]、花粉[16]和果实[29]的转录组数据开发了一些EST-SSR标记,而关于利用蓝莓花柱转录组数据开发EST-SSR以及利用EST-SSR标记开展自交不亲和基因鉴定的研究报道较少。本研究对蓝莓自花和异花授粉的花柱进行转录组测序,分析其SSR位点,对含SSR位点且差异表达的非冗余序列(Unigene)进行基因本体论(GO)功能富集和京都基因与基因组数据库(KEGG)注释分析,并开展引物设计,目的在于为蓝莓自交不亲和基因鉴定,亲和性品种的分子标记辅助育种以及自交不亲和的分子机制研究等提供参考。
1 材料与方法
1.1 试验材料
以兔眼蓝莓品种“杰兔”为研究对象,选取60棵植株作为供试材料;异花授粉的花粉来源于“灿烂”和“粉蓝”两个兔眼蓝莓品种;参试各品种材料均种植于凯里学院卓越农林人才培养实训基地内,为生长健壮且无病虫害的8年生植株。
1.2 授粉与取材
初花期,按照杨芩等[30]的蓝莓结果枝分类标准,选取“杰兔”参试植株树冠外围生长健壮且无病虫害的中结果枝和长结果枝,用镊子将1 500朵花蕾期花去雄后,采用尼龙网 (100目) 对去雄后的结果枝进行套袋隔离处理,然后挂牌标记各结果枝的去雄时间和花朵数,其余已开放或太小的花均去除。在去雄后的第3天,分别采集“粉蓝”“灿烂”和“杰兔”的花粉,并立即实施异花授粉和自花授粉[23]。“杰兔”ד粉蓝”、“杰兔”ד灿烂”以及“杰兔”ד杰兔”3个处理分别授粉450朵花,余下的150朵未授粉花的花柱为对照材料。分别取未授粉花及自花授粉和异花授粉后24、48和96 h的花,取后立即用液氮速冻并保存,带回实验室将材料置于经液氮预冷的无菌磁盘中,用手术刀将花柱从基部与子房分离,分离的花柱立即放入装满液氮和已标记相应处理标记的5 mL离心管中;各处理每分离50枚花柱即移入-80 ℃超低温冰箱中保存备用。
1.3 转录组测序及Unigene组装
委托生物科技公司采用Illumina NovaSeq 6000 platform(San Diego)平台对“杰兔”未授粉的花柱,以及自花和异花授粉后24、48和96 h的花柱,共10个不同处理的花柱进行转录组测序,获得10个蓝莓花柱转录组的Raw Reads数据。通过去除含有接头的Reads;去除N的比例大于10%的Reads;去除质量值Q≤10的碱基数占整条Reads 50%以上的Reads进行质量控制,获得高质量的序列,之后按照Beedanagari等[31]的方法利用Trinity组装软件对其进行Denovo拼接,得到Unigene;然后采用序列聚类软件对不同样品组装得到的Unigene进行进一步的序列拼接和去冗余处理,进而得到尽可能长的非冗余Unigene[1]。
1.4 SSR鉴定及含SSR的Unigene功能注释
采用MISA软件对1 kb以上的Unigene进行SSR序列鉴定,通过Blastx将筛选的含SSR的差异表达Unigene比对NR、SwissProt、KEGG、GO和COG/KOG数据库,获得含SSR的差异表达Unigene的GO基因功能、KOG分类和KEGG代谢途径[32]。以含SSR的差异表达Unigene功能富集分析为基础,以配子体自交不亲和花粉管信号转导过程为线索筛选蓝莓花柱自交不亲和及花粉管信号转导相关的SSR位点。
1.5 SSR位点信息的发掘与引物设计
按照杨芩等[16]的方法,采用MIcroSAtellite(MISA)工具进行SSR的识别鉴定,其最小单位数搜索设置条件分别为10、6、5、5、5和5个重复序列,通过对Unigene序列的SSR分析,鉴定出SSR 6的重复类型;在此基础上参照Yang等[33]的默认参数设置,采用Primer 3软件对获得的SSR进行引物设计。
2 结果与分析
2.1 蓝莓自花和异花授粉花柱转录组中SSR位点的分布特点
完成“杰兔”未授粉的花柱、自花和异花授粉后24、48和96 h花柱的转录组测序,各处理转录组的Clean Data均达到6.21 Gb以上,共获得67.12 Gb的Clean Data。10个处理的花柱转录组的Q30碱基百分比在94.35%以上。如表1所示,组装后共获得总长度82 908 720 bp的Unigene 85 890个,其中26 956条Unigene的长度在1 kb以上;N50长度1 547 bp,其中绝大多数Unigene的G+C含量46.09%~46.48%。搜索结果表明,共发现26 557个SSR位点,拥有SSR位点的Unigene 21 962个,即85 890个Unigene中SSR的发生频率25.57%。此外,在拥有SSR位点的21 962个Unigene中,有1 366个Unigene含有3个及以上的SSR位点,含有2个SSR位点的Unigene有709个。
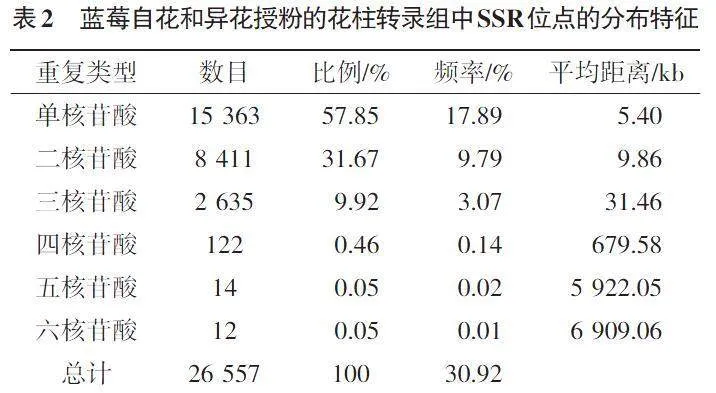
如表2所示,单核苷酸重复SSR出现频率最高(17.89%),所占的比例最大(57.85%);二核苷酸重复SSR次之,其出现频率和占比分别为9.79%和31.67%;两者出现的频率之和为27.68%,所占总SSR的比例之和为89.52%。即蓝莓花柱转录组中SSR的主要重复类型是单核苷酸重复SSR和二核苷酸重复SSR。四核苷酸、五核苷酸和六核苷酸重复SSR的数量较少,三者所占的比例总计0.56%,三者出现的频率分别为0.14%、0.02%和0.01%。
不同类型核苷酸重复SSR的平均分布距离与其出现频率密切相关,总体表现为出现频率越低,分布平均距离越大的趋势。六核苷酸重复SSR的出现频率最低(0.01%),其分布平均距离最大(6 909.06 kb);单核苷酸重复SSR的出现频率最高(17.89%),其分布的平均距离最小(5.40 kb)。五核苷酸、四核苷酸、三核苷酸和二核苷酸重复SSR的分布平均距离分别为5 922.05、679.58、31.46和9.86 kb。
2.2 蓝莓自花和异花授粉的花柱转录组SSR重复基元特点
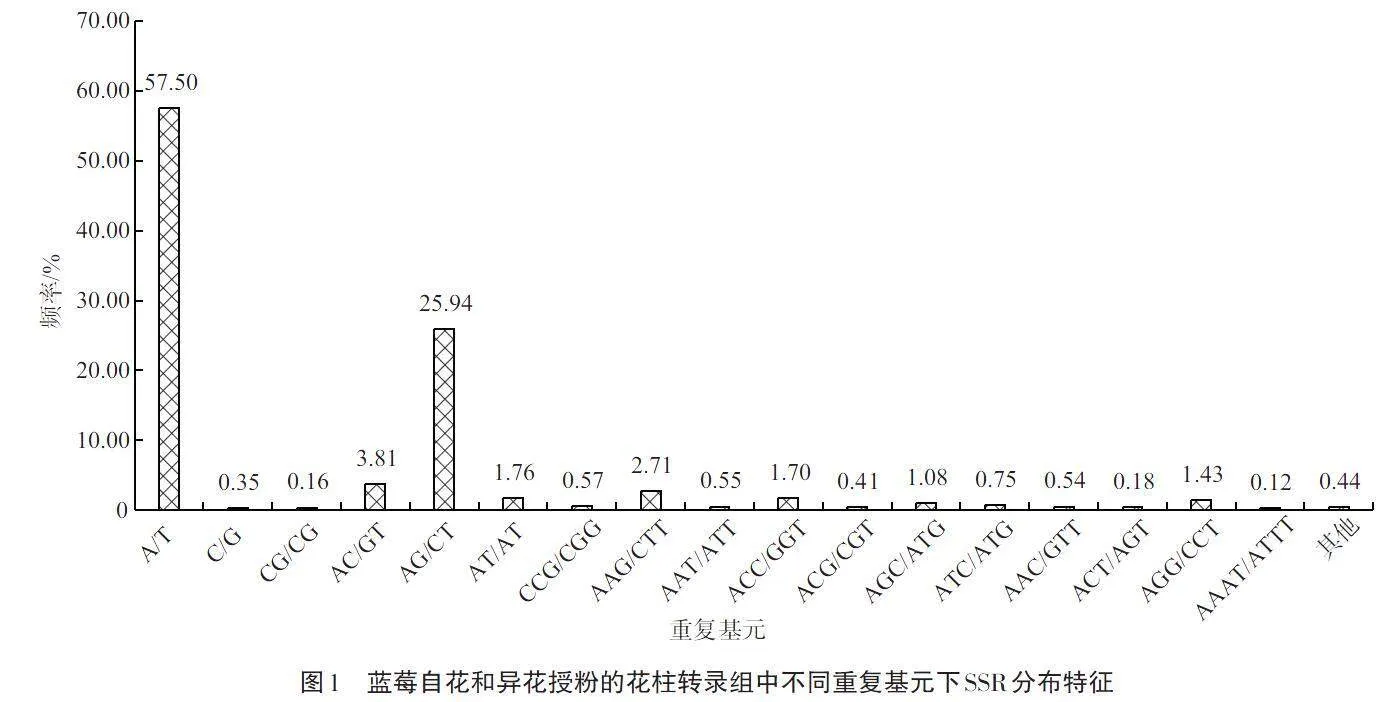
如图1所示,26 557个蓝莓自花和异花授粉的花柱转录组SSR的重复基元共有297种。重复基元的种类多少与SSR的重复类型和出现频率有着密切联系,五核苷酸和六核苷酸重复SSR的重复基元出现频率均在0.50%以下,其重复基元种类最少,均只有9种。单核苷酸重复SSR的重复基元种类相较五核苷酸和六核苷酸有所增加,为36种,其中A/T的重复基元在分布上占比最高,出现频率57.50%;二核苷酸重复SSR的重复基元种类相较单核苷酸重复SSR的增加19种,以AG/CT为重复基元的SSR出现频率最高,为25.94%。在6种核苷酸重复SSR类型中,三核苷酸重复SSR的重复基元共有147种,其中有721个SSR以AAG/CTT为重复基元,其出现频率在三核苷酸重复SSR中最高,为2.71%;其次是以ACC/GGT为重复基元的三核苷酸重复SSR,有451个,出现频率1.70%;再次是以AGG/CCT为重复基元的三核苷酸重复SSR,有380个,出现频率1.43%。
2.3 蓝莓自花和异花授粉的花柱转录组中SSR的重复次数和基序长度
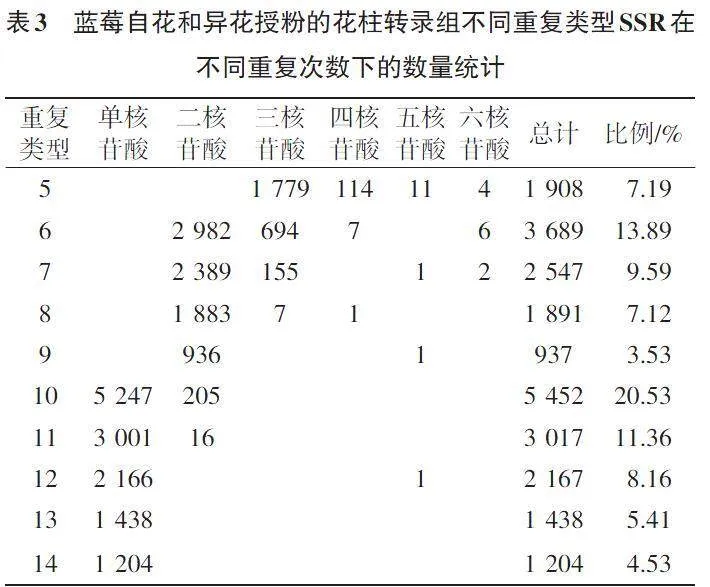
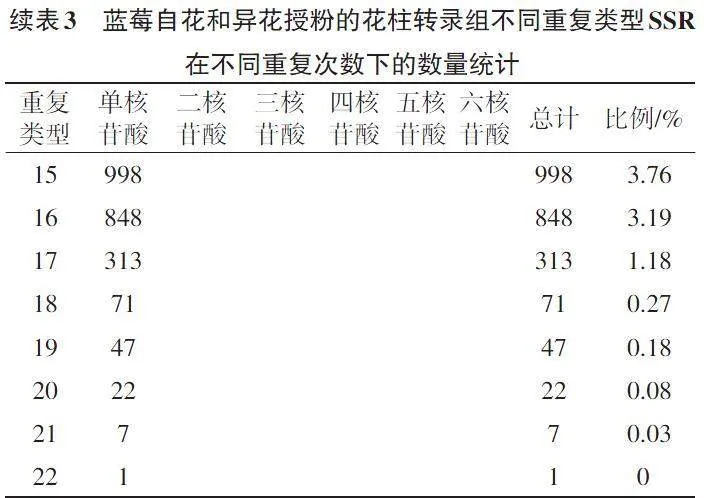
如表3所示,在10~22的SSR重复次数范围内,蓝莓自花和异花授粉花柱转录组6种类型SSR随着重复次数的增加,其总计数量呈现逐渐下降趋势。蓝莓自花和异花授粉花柱转录组SSR重复5~22次,大多重复5~16次。同时,重复10次的6种核苷酸重复SSR共有5 452个,出现频率最高,为20.53%;重复6次的6种核苷酸重复SSR有3 689个,占总数的13.89%;重复11次的6种核苷酸重复SSR有3 017个,占总数的11.36%。此外,有2 167个SSR的重复基元重复12次,占总数的8.16%;有1 438个SSR的重复基元重复13次,占总数的5.41%;6种核苷酸重复SSR,共有3 511个SSR的重复基元重复14次及以上,占总数的13.22%。
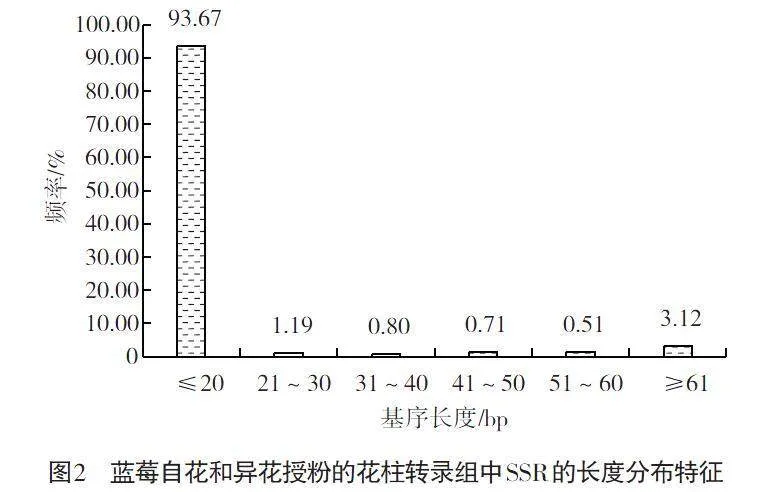
如图2所示,蓝莓自花和异花授粉的花柱转录组中有93.67%的SSR的基序长度12~20 bp,表明蓝莓花柱转录组SSR的基序长度以12~20 bp为主,长度21~60的基序占比3.21%,超过61 bp的基序长度占比3.12%。
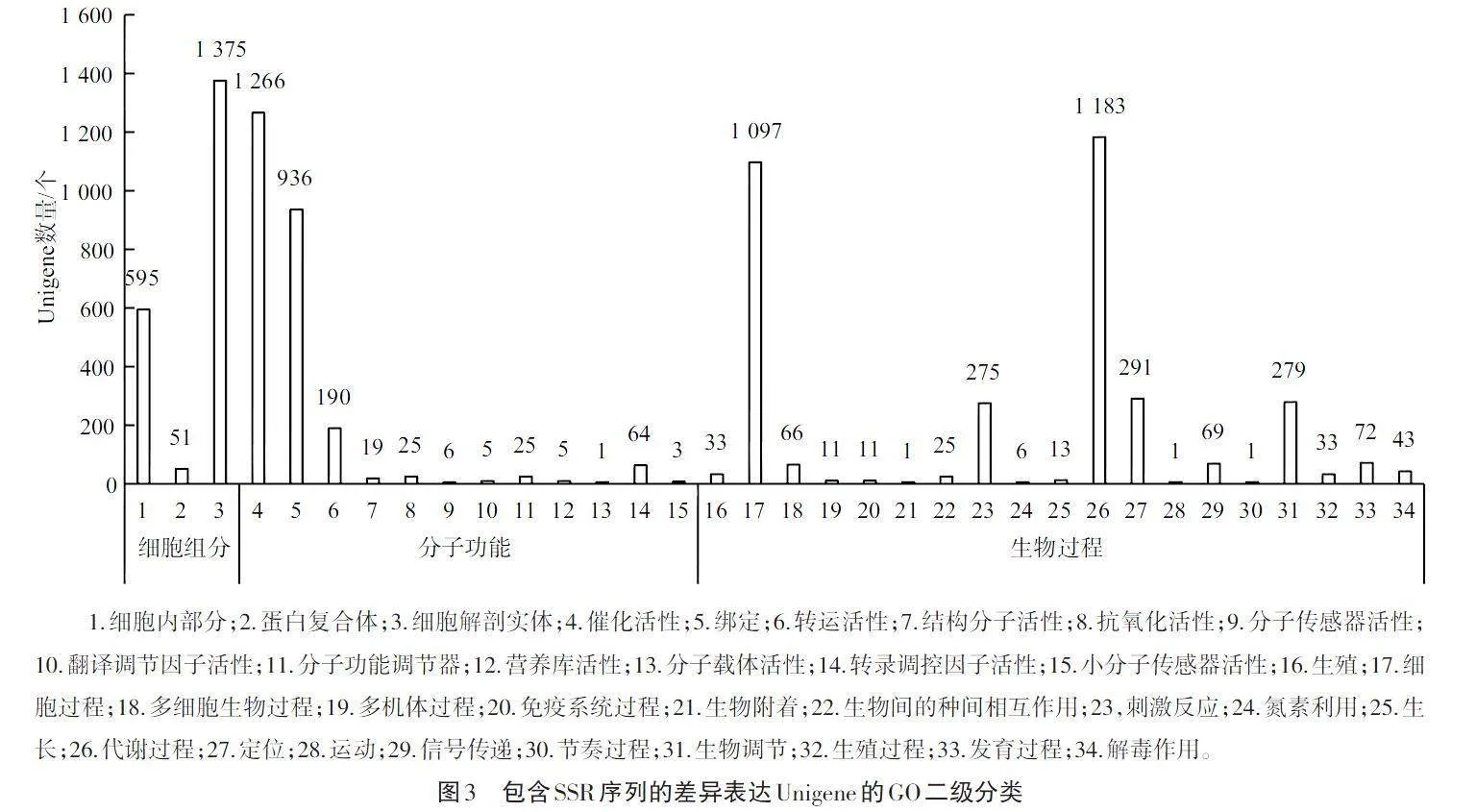
如图3所示,对含SSR的差异表达Unigene进行GO功能富集分析表明,在21 962个具SSR位点的Unigene中,有2 126个含有SSR位点的差异表达Unigene被富集到生物学、分子和细胞组分3个Ontology类别的34个GO Term中。细胞组分涉及的GO Term在3类Ontology中最少,为3个(8.82%),分子功能涉及的GO Term较多,为12个(35.29%),而生物过程涉及的GO Term最多,为19个(55.89%)。在生物过程中,代谢过程和细胞过程富集具有SSR位点的差异表达Unigene分别为1 183和1 097个,余下的代谢类别中,定位、生物调节和刺激反应3类代谢过程富集具有SSR位点的差异表达Unigene数量分别为291、279和275个;余下的代谢类别富集具有SSR位点的差异表达Unigene均未超过100个。在分子过程中,催化活性和绑定两类代谢过程富集具有SSR位点的差异表达Unigene较为丰富,分别有1 266和936个。在细胞组分相关代谢中,仅有细胞内部分、蛋白复合体和细胞解剖实体三类代谢过程富集到具有SSR位点的差异表达Unigene,其中细胞解剖实体代谢过程富集的具有SSR位点的差异表达Unigene有1 375个。
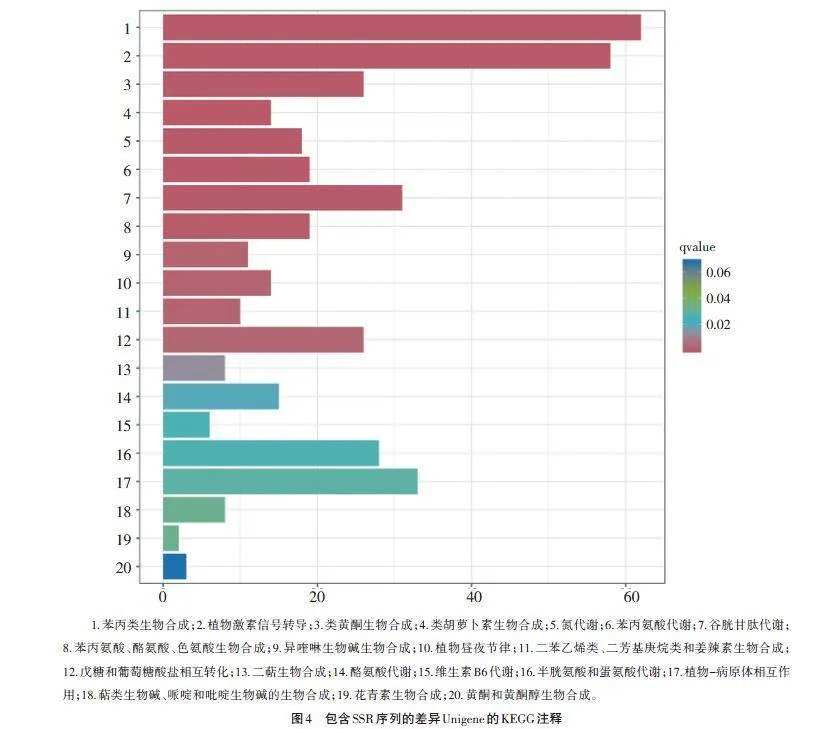
如图4所示,对含SSR位点的差异表达Unigene进行KEGG功能富集分析表明,共有659个含SSR位点的差异表达Unigene被富集到112个代谢途径中,其中被明显(qvaluelt;0.02)富集的代谢途径有12个,包括苯丙类生物合成(ko00940),植物激素信号转导(ko04075),类黄酮生物合成(ko00941),类胡萝卜素生物合成(ko00906),氮代谢(ko00910),苯丙氨酸代谢(ko00360),谷胱甘肽代谢(ko00480),苯丙氨酸、酪氨酸、色氨酸生物合成(ko00400),异喹啉生物碱生物合成(ko00950),植物昼夜节律(ko04712),二苯乙烯类、二芳基庚烷类和姜辣素生物合成(ko00945),糖和葡萄糖酸盐相互转化(ko00040)。58个含有SSR位点的差异表达Unigene被富集到植物激素信号转导途径,其中与生长素合成信号转导相关的有23个,与细胞分裂素信号转导相关的有5个,与赤霉素信号转导相关的有3个,与脱落酸信号转导相关的有10个,与乙烯信号转导相关的有5个,与油菜素内酯信号转导相关的有4个,与茉莉酸信号转导相关的有6个,与水杨酸信号转导相关的有4个。同时,进一步筛选获得与蛋白质泛素化、花粉管生长、核糖核酸酶T2家族以及细胞程序化死亡等相关的含有SSR位点的差异表达Unigene分别有16、3、2和1个。此外,KOG功能富集分析筛选出27个与细胞骨架相关的含有SSR位点的差异表达Unigene。
2.5 蓝莓自花授粉和异花授粉花柱转录组SSR的引物设计
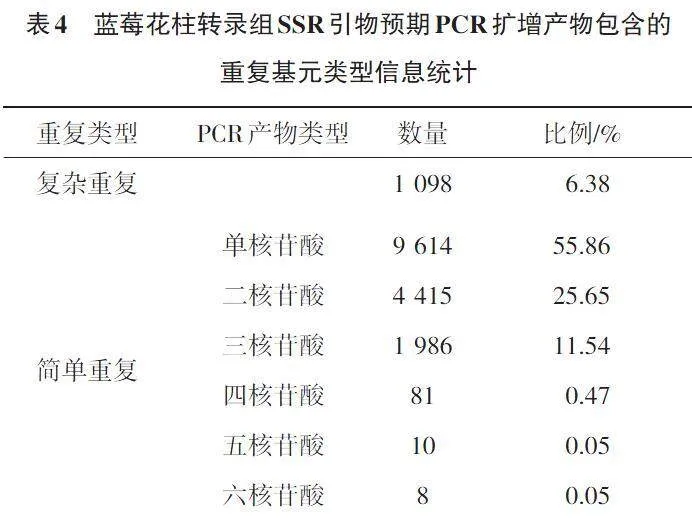
如表4所示,对21 962个含有SSR位点的Unigene采用Primer 3进行引物设计,成功率78.37%,共设计长度20~25 bp的SSR引物17 212对,预测这些引物扩增产物的大小100~280 bp。在成功设计的17 212对SSR引物中,预测的扩增产物为单核苷酸重复的引物占比最高,为55.86%,有9 614对;有4 415对引物的预测扩增产物为二核苷酸重复,占比25.65%;有1 986对引物的预测扩增产物为三核苷酸重复,占比11.54%。预测扩增产物为四核苷酸、五核苷酸和六核苷酸的引物占比均未超过1.00%。此外,在17 212对SSR引物中,有1 098对引物的预测扩展产物为复杂重复类型,占比6.38%。
3 结论与讨论
本研究通过对兔眼蓝莓“杰兔”未授粉花柱,自花和异花授粉花柱转录组总长82.91 Mb的85 890条序列进行检测,共获得26 557个SSR位点,SSR在所检测序列中的出现频率30.92%。SSR位点在“杰兔”花柱转录组序列中出现的频率,高于蓝莓花芽(12.67%)[27]、蓝莓叶片(21.32%)[28]和蓝莓果实(20.81%)[29],略低于蓝莓花粉(35.28%)[16],这可能是因为转录组测序所选取的器官不同。同时,本研究中蓝莓花柱转录组中SSR的出现频率高于黑莓果实(11.29%)[2]、梨果皮(12.45%)[11]、薄壳山核桃(21.29%)[1]、白掌(22.06%)[32]、黑果枸杞(26.36%)[13],低于油梨花和小果(35.05%)[3]、柿雌雄花(38.49%)[12和枇杷花(47.51%)[14],这可能与自花授粉和异花授粉后花柱中的一些基因具有时空表达和组织表达特性,也可能与不同植物的基因结构差异或设置的SSR搜索参数不同有关[2]。
SSR的重复次数和基本重复基元具有高度变异性,两者的变化均能导致SSR位点长度变化,进而使SSR拥有较强的多态性[2,14]。本研究发现单核苷酸和二核苷酸重复SSR是蓝莓花柱转录组SSR的主要重复类型,两者占总SSR的89.52%,出现频率27.68%;研究结果与在枇杷花[14]、薄壳山核桃[1]和蓝莓花粉[16]中研究的结果一致,但与蓝莓花芽和叶片转录组SSR分析结果不完全一致[27],蓝莓花芽和叶片的转录组SSR的主要重复类型为二核苷酸和三核苷酸重复,这可能与材料不同有关。此外,本研究发现,单核苷酸和二核苷酸重复序列的优势基元分别为A/T和AG/CT,这与在蓝莓花芽[27]、叶片[28]、果实[29]和花粉[16]转录组SSR信息位点优势基元分布的结果基本一致,这可能与AT键因能量较GC键低容易波动和断裂有关[14]。
SSR分子标记的基元重复次数和序列片段长度对其多态性高低程度有着重要的影响,一般来说,SSR基元的重复次数越高,其含多态性的位点越多,表明其开发多态性SSR分子标记的潜力越大[2]。本研究中蓝莓自花和异花授粉后花柱转录组的SSR重复5~22次,其中有26.79%的SSR重复12次以上;同时,蓝莓自花和异花授粉的花柱转录组中有93.67%的SSR的基序长度12~20 bp,基序长度≥20 bp的SSR占比6.33%,这表明蓝莓自花和异花授粉花柱转录组SSR的重复次数较高,具有良好的多态性潜能。此外,针对含有SSR位点的21 962个Unigene开展引物设计,共成功设计获得17 212对SSR引物,成功率78.37%,这与蓝莓花粉[16]、梨果皮[11]转录组开发SSR引物的成功率相近。
植物在包含时间和空间限定下的特定环境中发生的生理应激反应会引起某些特异功能基因的时空差异表达和组织差异表达,进而导致其生长和生理等表型特征发生变化。因此,开展不同处理及处理后不同时间材料的转录组测序能够高效筛选研究对象的功能基因,在此基础上进一步开展含SSR位点的差异表达基因筛选及SSR引物的设计,可能获得与目标性状直接相关联的定向标记[32]。自交不亲和是果树植物优质高产的重要性状之一,相关研究表明,蓝莓自花授粉和异花授粉后花粉管在花柱中的生长有明显差异,自花授粉的花粉管易出现在花柱中下部停止生长,表现出自交不亲和的现象[22-23]。本研究基于“杰兔”自花授粉、异花授粉后第24、48和96 h的花柱,及未授粉花柱共10个转录组数据的KEGG和KOG功能富集分析显示,659个具有SSR位点的差异表达基因被富集到112个代谢途径中,其中58、16、3、2、1和27个含SSR 位点的差异表达基因分别与植物激素信号转导、蛋白质泛素化、花粉管生长、核糖核酸酶T2家族、细胞程序化死亡以及细胞骨架等与植物自交不亲和、花粉管信号转导代谢途径相关[33]。
综上,本研究从高质量的蓝莓转录组中获得21 962个Unigene含有26 557个SSR位点,针对所有SSR位点,共设计17 212对引物,成功率78.37%。同时,筛选出107个与自交不亲和及花粉管生长信号转导等相关联的SSR功能位点,这不仅丰富了蓝莓的EST-SSR分子标记,而且为蓝莓种质鉴定、亲和性品种的分子标记辅助育种、自交不亲和基因鉴定以及自交不亲和的分子机制等研究提供了参考。
参考文献
[1] 贾展慧,宣继萍,张计育,等. 薄壳山核桃转录组中的SSR位点信息分析[J]. 分子植物育种,2019,17(10):3305-3311.
[2] 谢倩,张诗艳,江来,等. 橄榄转录组SSR信息分析及分子标记开发与应用[J]. 园艺学报,2023,50(11):2350-2364.
[3] 郭俊,朱婕,谢尚潜,等. 油梨转录组SSR分子标记开发与种质资源亲缘关系分析[J]. 园艺学报,2020,47(8):1552-1564.
[4] 叶宇芸,王清明,马建伟,等. 基于SSR标记的桃品种遗传多样性分析及遗传相似系数比较[J]. 华南农业大学学报,2017,38(3):39-45.
[5] 王丽媛,孙鹏,张嘉嘉,等. 柿野生雄性资源调查及其遗传多样性研究[J]. 园艺学报,2018,45(2):261-278.
[6] KIM H T,NOU I S. Confirmation of parentage of the pear cultivar ‘niitaka’ (Pyrus pyrifolia) based on self-incompatibility haplotypes and genotyping with SSR markers[J]. Horticultural science and technology,2016,34(3):453-460.
[7] AGARWAL M,SHRIVASTAVA N,PADH H. Advances in molecular marker techniques and their applications in plant sciences[J]. Plant cell reports,2008,27(4):617-631.
[8] DÍAZ A,MARTÍN A,RALLO P,et al. Self-incompatibility of ‘Arbequina’ and ‘Picual’ olive assessed by SSR markers[J]. Journal of the American society for horticultural science,2006,131(2):250-255.
[9] PINCZINGER D,VON RETH M,HANKE M V,et al. Self-incompatibility of raspberry cultivars assessed by SSR markers[J]. Scientia horticulturae,2021,288:110384.
[10] 姚利华,滕元文. EST-SSR标记及其在果树研究中的应用[J]. 果树学报,2008,25(2):219-224.
[11] 薛华柏,杨健,王龙,等. 梨果皮转录组SSR位点信息分析[J]. 果树学报,2018,35(增刊1):55-60.
[12] 杜改改,孙鹏,索玉静,等. 基于柿雌雄花芽转录组测序的SSR和SNP多态性分析[J]. 中国农业大学学报,2017,22(10):45-55.
[13] 尹跃,安巍,赵建华,等. 黑果枸杞转录组SSR信息分析及分子标记开发[J]. 浙江农林大学学报,2019,36(2):422-428.
[14] 付燕,杨芩. 枇杷花转录组SSR位点信息分析[J]. 分子植物育种,2021,19(18):6080-6087.
[15] 杨雪莲,吴永飞,王霞,等. 黔东南地区蓝莓果实发育特性研究[J]. 山东农业科学,2023,55(10):66-73.
[16] 杨芩,付燕,刘雅兰,等. 蓝莓花粉转录组SSR位点信息分析[J]. 分子植物育种,2021,19(10):3383-3391.
[17] 熊颖,禹霖,柏文富,等. 不同品种蓝莓果实品质特征和抗氧化能力及多酚组成的比较[J]. 中南林业科技大学学报,2022,42(2):119-128.
[18] YANG Q,LIU E,FU Y,et al. High temperatures during flowering reduce fruit set in rabbiteye blueberry[J]. Journal of the American society for horticultural science,2019,144(5):339-351.
[19] 杨芩,付燕,陆承锐,等. 蓝莓坐果率与花柱内源激素和多胺的关联分析[J]. 经济林研究,2023,41(1):14-25.
[20] TABER S K,OLMSTEAD J W. Impact of cross- and self-pollination on fruit set,fruit size,seed number,and harvest timing among 13 southern highbush blueberry cultivars[J]. HortTechnology,2016,26(2):213-219.
[21] KENDALL L K,GAGIC V,EVANS L J,et al. Self-compatible blueberry cultivars require fewer floral visits to maximize fruit production than a partially self-incompatible cultivar[J]. Journal of applied ecology,2020,57(12):2454-2462.
[22] 杨芩,罗莉,罗娅,等. 自花和异花授粉对蓝莓花粉管动力学的影响[J]. 中国果树,2020(5):20-25.
[23] 杨芩,付燕,宋宇,等. 授粉受精关键生物学因素对兔眼蓝莓自花授粉坐果率的影响[J]. 浙江农业科学,2023,64(2):344-350.
[24] 杨芩,张建兰,张婷渟,等. 不同授粉品种对“灿烂” 兔眼蓝莓着果率与果实性状的影响[J]. 中国南方果树,2017,46(6):93-95,99.
[25] GISBERT A D,ROMERO C,MARTÍNEZ-CALVO J,et al. Genetic diversity evaluation of a loquat (Eriobotrya japonica (Thunb) Lindl) germplasm collection by SSRs and S-allele fragments[J]. Euphytica,2009,168(1):121-134.
[26] MONTEMURRO C,DAMBRUOSO G,BOTTALICO G,et al. Self-incompatibility assessment of some Italian olive genotypes (Olea europaea L.) and cross-derived seedling selection by SSR markers on seed endosperms[J]. Frontiers in plant science,2019,10:451.
[27] LIU Y C,LIU S,LIU D C,et al. Exploiting EST databases for the development and characterization of EST-SSR markers in blueberry (Vaccinium) and their cross-species transferability in Vaccinium spp.[J]. Scientia horticulturae,2014,176:319-329.
[28] 方茜,张园园,杨钰婷,等. 越橘叶片转录组SSR发掘及其多态性研究[J]. 园艺学报,2018,45(7):1359-1370.
[29] 王小敏,张春红,胡丽超,等. 基于高通量测序的蓝莓果实转录组SSR信息分析[J]. 北方园艺,2022(13):49-54.
[30] 杨芩,李性苑,秦绍钊,等. 结果枝类型对兔眼蓝莓着果率和果实品质的影响[J]. 中国南方果树,2015,44(3):133-135,138.
[31] BEEDANAGARI S R,DOVE S K,WOOD B W,et al. A first linkage map of pecan cultivars based on RAPD and AFLP markers[J]. Theoretical and applied genetics. Theoretische und angewandte Genetik,2005,110(6):1127-1137.
[32] 周辉明,林辉锋,曹奕鸯,等. 基于白掌株型突变转录组差异的SSR位点分析[J]. 分子植物育种,2023,21(16):5365-5377.
[33] YANG X D,TAN H W,ZHU W M. SpinachDB:a well-characterized genomic database for gene family classification and SNP information of spinach[J]. PLoS One, 2016,11(5):e0152706.
(责编:李 媛)
猜你喜欢
西北植物学报(2023年9期)2023-10-17 13:34:00
河北果树(2021年4期)2021-12-02 01:15:08
中国果树(2020年2期)2020-07-25 02:14:16
音乐教育与创作(2020年2期)2020-06-19 06:45:46
数学小灵通(1-2年级)(2018年12期)2018-12-24 13:16:12
商周刊(2017年5期)2017-08-22 03:35:22
作文周刊·小学一年级版(2017年26期)2017-08-10 20:31:15
中国中药杂志(2017年2期)2017-03-25 17:23:23
中国中药杂志(2017年1期)2017-03-06 21:20:38
中国中药杂志(2016年22期)2017-02-13 17:06:29
