
喀斯特原生林9个树种水力学性状与解剖结构对树干液流的影响
2024-12-12 00:00:00叶雨艳丁访军吴鹏周华李源永周汀崔迎春
南京林业大学学报(自然科学版) 2024年6期





摘要:【目的】揭示茂兰喀斯特森林主要树种水力结构和解剖结构特征及其对树干液流密度的影响。【方法】以茂兰喀斯特森林9个树种为研究对象,用“冲洗法”测定其导水率(Kh)、比导率(Ks)、胡伯尔值(Hv);以解剖学方法测定其导管面积(A)、导管直径(D)、导管水力学直径(Dh)、导管密度(WD)、管胞壁厚度(WVD)等解剖结构参数;采用热扩散探针法监测树干液流密度(Js),并对已测的几个指标进行排序,比较各树种抗旱能力强弱。【结果】①Kh从大到小表现为南酸枣(Choerospondias axillaris)gt;柿(Diospyros kaki)gt;山乌桕(Triadica cochin chinensis)gt;化香(Platycarya strobilacea)gt;鹅耳枥(Carpinus turczaninovii)gt;新木姜子(Neolitsea aurata)gt;山矾(Symplocos sumuntia)gt;天峨槭(Acer wangchii)gt;润楠(Machilus nanmu);Ks从大到小表现为南酸枣gt;山乌桕gt;柿gt;化香gt;鹅耳枥gt;天峨槭gt;新木姜子gt;山矾gt;润楠;Hv表现为鹅耳枥gt;新木姜子gt;润楠gt;山矾gt;化香gt;山乌桕gt;天峨槭gt;南酸枣gt;柿,落叶树种的Kh和Ks大于常绿树种;②除鹅耳枥外,落叶树种的导管面积、直径、水力直径均大于常绿树种,而导管密度则相反;③各树种的Kh、Ks、Hv与Js的相关性均不显著(Pgt;0.05),山乌桕树干液流密度与水力直径呈显著正相关(Plt;0.05),而天峨槭与水力直径呈显著负相关(Plt;0.05),山矾的液流密度与导管密度呈显著正相关(Plt;0.05),其余树种的解剖结构与树干液流密度相关性均不显著(Pgt;0.05);④树种抗旱能力随导管直径的增大而增加,从大到小表现为南酸枣gt;山乌桕gt;柿gt;化香gt;鹅耳枥gt;新木姜子gt;山矾gt;润楠gt;天峨槭。【结论】茂兰喀斯特森林中落叶树种的导水、输水和抗旱能力均强于常绿树种。水分充足时,水力学性状对树干液流密度的影响较小,而解剖结构对其影响存在种间差异;优势树种通过增大单位横截面积上的蒸腾拉力抵消树高带来水分运输阻力,为叶片蒸腾和光合作用供给水分。在进行喀斯特植被恢复时应综合考虑落叶、常绿及林分垂直结构的群落配比。
关键词:茂兰喀斯特森林;树干液流密度;水力学性状;解剖结构;抗旱能力
中图分类号:S718.5"""""" 文献标志码:A开放科学(资源服务)标识码(OSID):
文章编号:1000-2006(2024)06-0111-10
Effects of hydraulics and anatomical structure on sap flow of nine tree species in Karst primary forest
YE Yuyan1,2, DING Fangjun2,3*, WU Peng2, ZHOU Hua2, LI Yuanyong1,2, ZHOU Ting1,2, CUI Yingchun2
(1. Key laboratory of Plant Resource Conservation and Germplasm Innovation in Mountainous Region, Ministry of Education, College of Life Sciences/Institute of Agro-Bioengineering, Guizhou University, Guiyang" 550025, China; 2. Guizhou Academy of Forestry, Guiyang 550005, China; 3. Guizhou Libo Observation and Research Station for Karst Forest Ecosystem, National Forestry and Grassland Administration, Libo 558400, China)
Abstract: 【Objective】This study aims to explore the hydraulic and anatomical characteristics of predominant tree species in the Maolan Karst Forest and their impact on sap flow density.【Method】 The hydraulic conductivity (Kh), specific conductivity (Ks), and Huber value (Hv) of nine tree species in the Maolan Karst Forest were determined through the ‘washing method’. Anatomical structure parameters such as vessel area (A), vessel diameter (D), hydraulic vessel diameter (Dh), vessel density (WD), and vessel wall thickness (WVD) were assessed using anatomical techniques. The thermal diffusion probe method was employed to monitor the sap flow density (Js) of the tree trunks. The measured indexes were subsequently ranked via the subordinate function method to discern drought resistance. 【Result】 (1)The Kh value behaved Choerospondias axillarisgt; Diospyros kakigt; Triadica cochinchinensisgt; Platycarya strobilacea gt; Carpinus turczaninovii gt; Neolitsea aurata gt; Symplocos sumuntia gt; Acer wangchiigt; Machilus nanmu, the Ks value behaved C. axillaris gt; T. cochinchinensis gt; D. kaki gt; P. strobilacea gt; C. turczaninovii gt; A. wangchii gt; N. aurata gt; S. sumuntiagt; M. nanmu, and the Hv value behaved C. turczaninovii gt; N. aurata gt; M. nanmu gt; S. sumuntia gt; P. strobilacea gt; T. cochinchinensis gt; A. wangchii gt; C. axillaris gt; D. kaki, the Kh and Ks values of deciduous trees are higher than those of evergreen trees. (2) In addition to C. turczaninowii, the vessel area, vessel diameter and hydraulic diameter value of eight trees revealed that deciduous trees were lager than evergreen trees. In contrast, the vessel density of evergreen trees was larger than that of deciduous trees; overall, the relevance of nine anatomical parameters varied between trees. (3) No significant correlation was found between the Kh, Ks, Hv of the nine trees and sap flow density (Plt;0.05), Sap flow density of T. chinensis showed a significantly positive correlation with vessel hydraulic diameter(Plt;0.05), while that of A. wangchii indicated a significantly negative correlation(Plt;0.05). Conversely, the sap flow density of S. sumuntia positively correlated with vessel density (Plt;0.05), but the sap flow density of the remaining six species didn’t significantly correlate with anatomical structure parameters (Pgt;0.05); (4) Drought resistance of the nine species increased with vessel diameter, ranking as follows: C. axillarisgt; T. cochin" chinensis gt; D. kakigt; P. strobilacea gt; C. turczaninovii gt; N. auratagt; S. sumuntiagt; M. nanmugt; A. wangchii. 【Conclusion】The water conductivity, water transport, and drought resistance of the nine studied species were superior to evergreens. While water availability was ample, the hydraulic traits minimally impacted the sap flow density, whereas the anatomical structure variably influenced it. The dominant species managed to counterbalance the water transport resistance caused by tree height by boosting transpiration tension per unit cross-sectional area, thus facilitating leaf transpiration and photosynthesis. For the effective restoration of Karst vegetation, it is essential to consider the community ratio of deciduous to evergreen species and the stand vertical structure.
Keywords:Maolan Karst forest; sap flow density;" hydraulic trait; anatomy structure; drought resistance capacity
全球气候变暖导致水文循环加快,使得干旱发生频率加快、时间延长[1]。在长期干旱下,植物会发生水力失效,进而导致植物死亡[2-3]。与干旱地区相比,湿润地区的树木缺乏适应干旱的条件,在面对干旱时更易发生水力失效[4]。尤其是高大乔木,水力运输途径长,加大了水力失效风险,因此水分在高大乔木体内的运输是植物对生态环境适应的重要论题。树木水力学主要研究木质部水分运输过程和机理,包括水分传导的效率性和安全性,是探讨树种生理生态适应机制的关键内容[5],水力学结构及解剖结构是树木水力学的重要表征参数,蒸腾速率是植物从下到上运输水分效率的宏观表征,同为表征水分运输效率的重要参数,开展蒸腾速率与水力学结构及解剖结构之间关系的研究[6-10],有助于认识树木的水分生理生态特征,阐明树木的耐旱特性以及树木蒸腾的生理调控机理。
茂兰喀斯特森林是世界上同纬度地区仅存的原生性喀斯特森林生态系统,其生态环境、群落外貌、组成特征、垂直结构和演替动态等方面与常态地貌的森林生态系统相比存在明显的差异[11],由于土层薄且不连续,碳酸盐岩基岩易溶蚀、岩隙发达导致水分下渗等特征,极易形成“岩溶性”干旱生境[12],水分成为限制喀斯特地区植物生长和分布的主要因子[13],因此研究这一地区植物特别是乔木树种的水分生理生态特征,揭示其对“岩溶性”干旱生境的适应策略尤为重要。目前已有学者测定了贵州普定、花江、荔波喀斯特地区圆叶乌桕(Triadica rotundifolia)、朴树(Celtis sinensis)、青冈(Quercus glauca)[14]等常见种的水力学特性和解剖结构特征,同时也有学者研究茂兰喀斯特原生林树种的树干液流特征及其对叶片气孔[15]和环境因子[16-17]的响应,但针对茂兰喀斯特原生林树种的水力学性状与解剖结构特征方面的研究较为缺乏,特别是树种的水力学性状与解剖结构特征对树干液流密度的影响方面鲜见报道。为此,本研究以茂兰喀斯特原生林9个优势树种为研究对象,分析其水力学性状和木质部解剖结构特征,以及两者与树干液流密度的关系,以期探明喀斯特森林主要树种的导水能力、输水能力和木质部解剖结构等水力结构的差异及其对树干液流密度的影响,阐明树种的抗旱能力,揭示喀斯特森林对“岩溶性”干旱生境的适应策略,补充岩溶地区木本植物水力结构的研究体系,为茂兰喀斯特原生林保护提供理论支撑,也为石漠化地区植被恢复时物种选择等提供科学依据。
1 材料与方法
1.1 研究区概况及样地选择
研究地位于贵州荔波喀斯特森林生态系统国家定位观测研究站内(107°52′2″~108°5′7″E, 25°9′3″~25°20′8″N),该地属中亚热带湿润季风气候,雨量充沛但年内分布不均,4—10月降雨集中,10月到次年3月降雨较少,年均降水量1 725.5 mm;年平均气温15.3℃;年太阳辐射总量63 289.8 kW/m2。该地区碳酸盐岩的透水性强,成土速度慢,土壤持水量低,呈现地下径流丰富、地表水缺乏的“岩溶性”干旱生境特征。
样地选择生态站原有固定样地(107°33′36″E,25°10′12″N),样地面积为900 m2,海拔742 m,坡度26°,坡向为东坡,植被类型为常绿落叶阔叶混交林,主要优势种包括润楠(Machilus nanmu)、天峨槭(Acer wangchii)、化香(Platycarya strobilacea)、鹅耳枥(Carpinus turczaninovii)等。
1.2 研究方法
1.2.1 树干液流观测方法
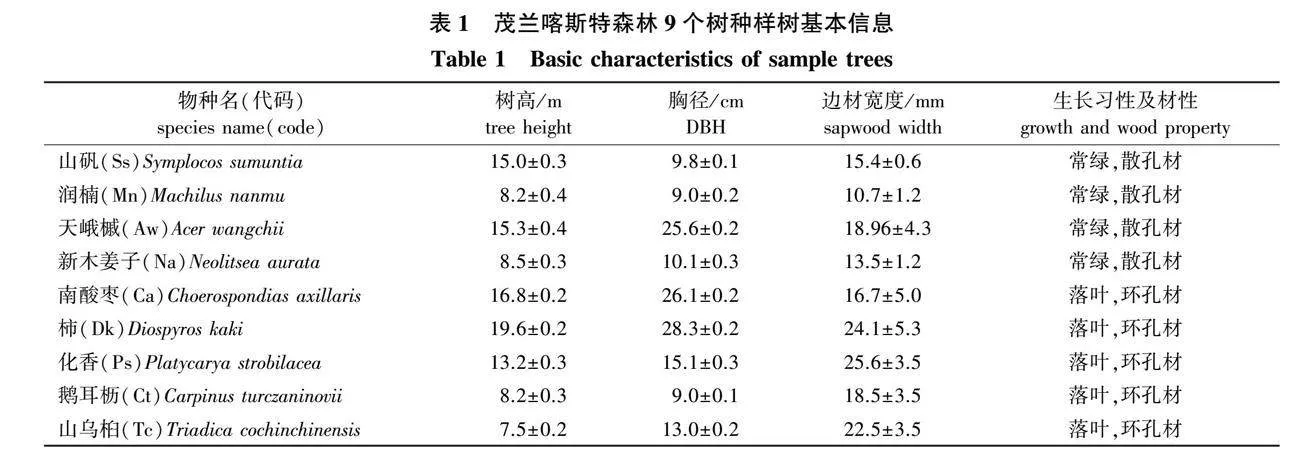
在样地调查的基础上,根据树种重要值选择观测样树(表 1)。采用热扩散探针法监测树干液流,使用Campbell公司的CR1000(美国)进行数据采集,记录步长为10 min。选用30 mm长的热扩散特征安装在胸高处(130 cm)[18]。树干液流(Js,式中记为DJs),计算方法参照Granier[19]经验公式:
DJs=119×10-6×[(ΔTmax-ΔT)/ΔT]1.231。(1)
式中:ΔTmax为每天的探针温差的最大值;ΔT为实时的探针温差。
1.2.2 枝条水力学性状测定及枝条木质部解剖结构观测
于2022年7月中旬、8月初和8月中旬,选择连续晴天的5:00—7:00分别从各观测样树上取向阳生健康枝条3枝,每个树种总样本量为27。采用樊大勇等[20]改良的冲洗法测定最大边材导水率,冲洗溶液为10 mmol/L的CaCl2溶液,并使用0.2 μm的微孔过滤器过滤溶液。首先,从取下的枝条上剪取约15 cm无侧枝的茎段,如果无法避开侧枝则需把侧枝剪掉用密封胶封死切口,在水下用刀片环剥1 cm左右的形态学下端表皮,露出木质部后在水下用生料带缠绕后与乳胶管连接防止漏水,胶管另一端连接三通阀、水压表、增压泵以及储水装置,给增压泵插上电源让管道内的溶液加压到0.1 MPa左右使得植物离体茎段产生水流量,用秒表计时20 min后计算单位时间水流量,用天平称重法计算单位时间内通过茎段的最大水流速度(V, g/s);用直尺测量枝条长度(L,cm);用游标卡尺测量茎段横截面和芯材直径并计算茎段边材横截面积(S,cm2);新鲜植株叶片摘下后用扫描仪(EPSON,2019,American)扫描并用Image J测量总叶片面积(LSA, cm2,式中记为SLSA),用于计算水力结构指标。导水率[Kh, g/(MPa·m·min),式中记为DKh],比导率[Ks, g/(cm2·MPa·m·min),式中记为DKs]和胡伯尔值(Hv,式中记为DHv)。各指标计算公式如下:
DKh=V×L/ΔP。(2)
DKs=DKh/S;(3)
DHv=S/SLSA。(4)
式中,ΔP为利用增压泵给离体茎段加压后茎段两端的压力差,MPa。
将剩余枝条置于FAA固定液内约7 d,带回实验室用蒸馏水冲洗1 min,置于体积分数10%的乙二胺溶液中软化3 d左右,根据木质部硬度不同软化时间有差异,软化的枝条取出后用蒸馏水冲洗1~2 min,用梯度浓度的叔丁醇脱水与透明后直接用半自动切片机(KD2260)切为20 μm左右薄片置于载玻片上,用体积分数1%番红(染色10 min)与0.5%固绿染色(10 s后用蒸馏水冲洗)处理,染色后置于4倍显微镜下拍摄3个视野用于导管密度的计算,20倍显微镜下拍摄50个导管用于导管面积(SA,μm2)和管胞壁厚度(WVD,μm)的测量。等效圆直径(D,μm)和导管水力学直径(Dh,μm,式中记为DDh),计算方法如下:
D=4SA/π;(5)
DDh=∑ni=1D5i/∑ni=1D4i。(6)
式中, Di为第i个导管的等效圆直径。
1.2.3 抗旱能力综合评价方法
与抗旱性成正比的指标隶属函数U(Xj)计算公式[21]如下:
U(Xj)=(Xj-Xj,min)/(Xj,max-Xj,min)。(7)
与抗旱性成反比的指标隶属函数U(Xj)计算公式如下:
U(Xj)=1-[(Xj-Xj,min)/(Xj,max-Xj,min)]。(8)
式中:Xj表示第j个综合指标;Xj,max与Xj,min分别表示第j个综合指标中的最大值与最小值。
1.2.4 数据分析
采用Excel 2019进行数据预处理与统计,用Image J分析木质部解剖结构参数,用SPSS 18.0进行树干液流与水力结构和解剖结构的Pearson相关性分析,所有图片的绘制在 Origin 8.1及Adobe Illustrator CC2018内完成。
2 结果与分析
2.1 9个树种水力学性状与树干液流密度比较
茂兰喀斯特森林9个主要树种导水率(Kh)为0.97~8.23 g/(MPa·m·min)(图1a),比导率(Ks)为1.49~12.72 g/(cm2·MPa·m·min)(图1b),胡伯尔值(Hv)为0.8×10-4~1.41×10-4(图1c),树干液流密度(Js)为1.17~4.25 g/(cm2·h)(图1d),南酸枣的导水率大、比导率和树干液流密度的值最大,润楠的导水率和比导率的值最小,山矾的树干液流值最小,鹅耳枥的胡伯尔值最大,柿树的胡伯尔值最小。
研究发现,各树种的胡伯尔值与树高呈线性关系,即随树高的增高而降低(图2),这是由于树高越高,单位叶面积蒸腾耗水供水所需的木质部结构越多,即胡伯尔值越小。同一森林中不同树种的导水率、比导率、胡伯尔值和树干液流密度均存在一定差异,这种差异在不同的种间表现各异。整体上看落叶树种的Kh和Ks大于常绿树种,表明在喀斯特原生林中,落叶树种的导水能力、输水能力普遍强于常绿树种。
2.2 9个树种木质部解剖结构特征比较
茂兰喀斯特森林9个树种的木质部解剖结构特征如表 2所示,导管面积大小表现为南酸枣gt;山乌桕gt;柿gt;化香gt;新木姜子gt;鹅耳枥gt;山矾gt;天峨槭gt;润楠,新木姜子和鹅耳枥以及山矾和天峨槭的导管面积种间差异不显著(Pgt;0.05),其余树种的导管面积种间差异显著(Plt;0.05)。导管直径大小表现为南酸枣gt;山乌桕gt;柿gt;化香gt;新木姜子gt;山矾gt;天峨槭gt;鹅耳枥gt;润楠,柿和化香、新木姜子和山矾及天峨槭和鹅耳枥的导管直径种间差异不显著(Pgt;0.05),其余树种的导管直径种间差异显著(Plt;0.05)。导管水力直径(Dh)大小表现为南酸枣gt;山乌桕gt;柿gt;化香gt;鹅耳枥gt;新木姜子gt;天峨槭gt;山矾gt;润楠,山乌桕和柿、鹅耳枥和新木姜子及天峨槭、山矾和润楠的Dh种间差异不显著(Pgt;0.05),其余树种的Dh种间差异显著(Plt;0.05)。导管密度(WD)大小表现为润楠gt;天峨槭gt;新木姜子gt;山矾gt;鹅耳枥gt;化香gt;柿gt;南酸枣gt;山乌桕,润楠和天峨槭、新木姜子和山矾及南酸枣、柿和山乌桕的WD种间差异不显著(Pgt;0.05),其余树种的WD种间差异显著(Plt;0.05)。管胞壁厚度(WVD)介于6.66~23.75 μm之间,大小表现为山矾gt;山乌桕gt;鹅耳枥gt;新木姜子gt;南酸枣gt;化香gt;润楠gt;柿gt;天峨槭,化香、南酸枣和新木姜子,山乌桕和鹅耳枥及柿和天峨槭的WVD种间差异不显著(Pgt;0.05),其余树种的WVD种间差异显著(Plt;0.05)。除鹅耳枥外,4个落叶树种的导管面积、导管直径、导管水力直径均大于常绿树种,而导管密度则表现相反。
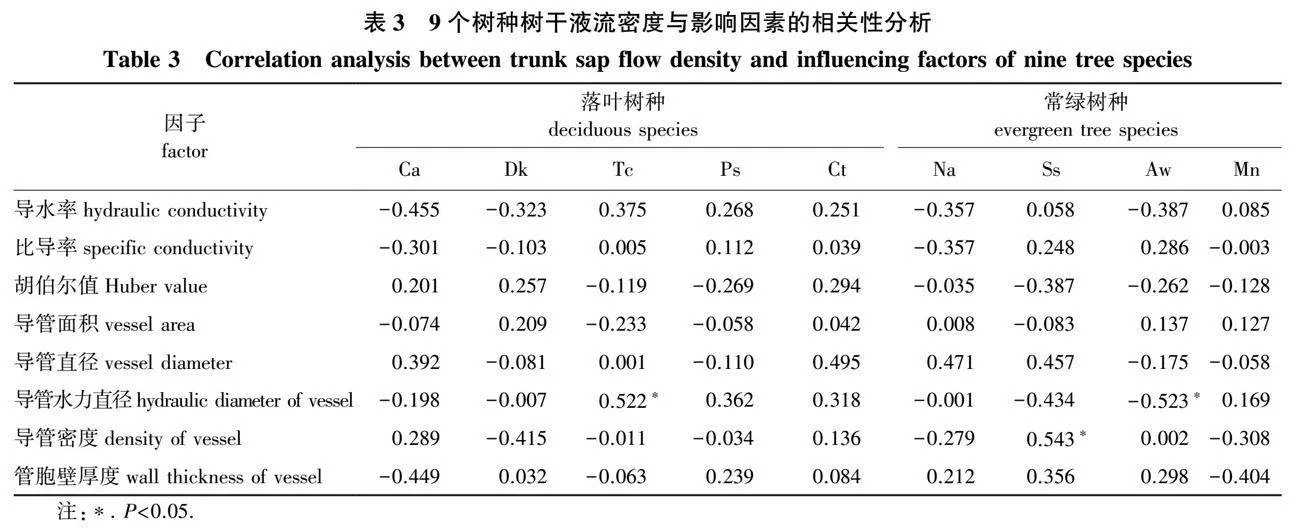
2.3 水力学性状和解剖结构与树干液流密度的相关性分析
通过对9个树种水力学性状和解剖结构与树干液流密度的相关性分析可知,山乌桕的树干液流密度与导管水力直径显著正相关(Plt;0.05),山矾的树干液流密度与导管密度显著正相关(Plt;0.05),天峨槭的树干液流密度与导管水力直径显著负相关(Plt;0.05),其余树种的树干液流密度与其解剖结构特征均无显著相关(Pgt;0.05),说明树种的木质部解剖结构对树干液流密度的影响因树种不同而存在差异。9个树种的树干液流密度与最大导水率(Kh)、比导率(Ks)和胡伯尔值(Hv)水力学参数均无显著相关(Pgt;0.05),水力结构参数均对其树干液流密度的影响较小(表3)。
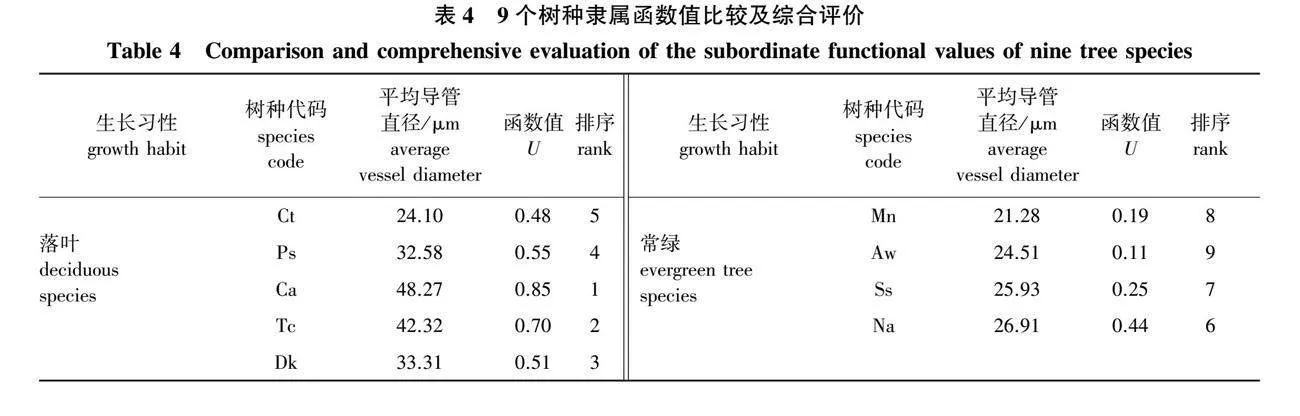
2.4 9个树种抗旱能力综合评价
植物抗旱能力受多种因素影响,在进行抗旱能力评价时,需采用多个指标进行综合评价,隶属函数法是普遍用于植物的抗旱能力排序的综合评价方法之一[22]。本研究选取了水力学性状、树干液流密度、解剖结构特征3个方面共9个评价指标,计算隶属函数均值,并对9个树种的隶属函数平均值进行排序,结果如表4所示,可知各树种抗旱能力强弱表现为南酸枣gt;山乌桕gt;柿gt;化香gt;鹅耳枥gt;新木姜子gt;山矾gt;润楠gt;天峨槭,整体上,落叶树种的抗旱能力强于常绿树种。
3 讨 论
3.1 喀斯特森林树种水力学性状与解剖结构特征的关系
本研究中,落叶树种的比导率比Fan等[23]在贵州省岩溶地区所测落叶植物比导率高,这可能是由于本研究中柿、南酸枣和山乌桕的导管直径较大,植物的导水能力与导管直径的4次方成正比[24],因此其比导率也偏大;常绿树种的比导率均值在Fan等[23]所测常绿植物的比导率范围之内。本研究发现9个树种的Kh和Ks存在一定的种间差异,这是因为水力学性状是植物在生长过程中木质部形态结构和输水策略对环境和生存竞争的适应,其种间差异主要取决于物种的生物学特征[24],同时由于喀斯特地区峰丛地貌带来的高异质性的生境使得不同物种获取资源的情况不同[25],因此其生理生态适应表现不尽相同,但小生境是如何影响树种的水分运输能力还待进一步的研究。本研究还发现落叶树种的导水和输水能力均强于常绿树种,主要有以下两点原因:①在本研究中,除鹅耳枥为散孔材外,其余落叶树种均为环孔材,常绿树种均为散孔材,环孔材树种具有蒸腾快、比导率高的特点[26],导管直径在不同材性间遵循环孔材树种gt;散孔材树种gt;无孔材树种的规律[27],根据Hagen-Poiseuille方程,植物的水分运输能力与导管直径的4次方成正比[28];②落叶树种和常绿树种不同的叶片生长习性使得落叶树种需要付出较高的生理代谢成本去获取更多的光、水资源以满足碳的合成[29],即落叶树种需要比常绿树种更强的导水和输水能力,大多数研究也证明了这一点[23, 30-31]。9个树种胡伯尔值与谭凤森[32]对桂西南北热带喀斯特季雨林所研究的结果接近。植物的胡伯尔值随树高的增高而降低,说明冠层树种通过增加单位横截面积上总叶面积的投资来获取更大的蒸腾拉力从而维持较高的水分运输能力,这与刘晓静等[33]和赵平等[34]对马占相思(Acacia mangium)的研究结果一致。
本研究中9个树种的平均水力直径与倪鸣源[35]在喀斯特地区所测被子植物的平均水力直径相近,均显著小于世界范围内木本植物的平均水力直径[36],这可能是由于喀斯特森林植物解剖结构对“岩溶性”干旱生境的适应,即通过降低导管直径来防止导管形成空穴[37]。本研究还发现生长在喀斯特原生林中的润楠的导管面积远大于其他地区润楠属树种的导管面积,导管密度则偏低(润楠属导管密度范围为228.86~426.87 个/mm2)[38];而鹅耳枥的导管直径比天台鹅耳枥(Carpinus tientaiensis)的结果偏大[39],化香的导管密度远大于韩丽娟等[40]在广东采集的化香树导管密度,喀斯特这些树种的解剖结构有别于生长在非喀斯特地貌上的同类植物,这是否是植物为适应喀斯特“岩溶性”干旱生境的策略还有待进一步的研究。
3.2 水力学性状和解剖结构对树干液流密度的影响
本研究9个树种的树干液流密度存在一定的种间差异,这是因为树干液流密度受环境因子和自身生物因子的综合影响[41-42]。整体上看落叶树种的树干液流密度大于常绿树种,这可能是由于落叶树种的输水能力和导水能力强于常绿树种,且落叶树种需要在生长季积蓄更多的能量[43],相同叶面积上拥有更多的气孔[15-16]导致的。
在本研究中,9个树种的Ks、Kh、Hv对树干液流密度无显著影响,与赵延涛等[24]的研究结果一致。这是由于植物Ks、Kh和Hv等参数是植物水力结构的表征,植物水力结构是长期适应环境和生存策略的形态结构,在水分充足的条件下(本研究在2022年7—8月进行,水分充足)具有稳定性[14],而树干液流密度是植物蒸腾作用的表征,受更宏观因子(环境因子)的调控。只有当植物水分出现亏缺时,或者当植物的水分供给失衡时,水分因子在决定栓塞程度上占主导地位[42],植物的水力学特性就发挥调控作用,调节植物失水和供水的动态平衡。
本研究中,山乌桕的树干液流密度与导管水力直径显著正相关,与对橡胶树的研究结果一致[44]。天峨槭的树干液流密度与导管水力直径呈显著负相关,王秀伟等[45]发现白桦、兴安落叶松和水曲柳的树干液流密度与水力直径也呈现相同的结果。山矾的树干液流密度与导管密度呈显著正相关,酸枣和荆条在干旱季节表现出同样的特征[46]。另外6个树种的解剖结构与树干液流密度均无显著相关性,与赵培强等[47]的研究结果一致。本研究的9个树种树干液流密度与解剖结构之间的相关性存在种间差异,这是由于在喀斯特原生林中,基岩的不保水性和小生境的高异质性,导致供给植物的水分不一致,有的植物在缺水状态下通过增加导管直径、降低厚度跨度比来适应干旱[48],有的植物在水分缺乏时通过增加导管数量、降低导管面积来适应不利生境[49],有的植物导管解剖结构特征在种内具有一定的稳定性,不随生境的改变而改变[50]。因此,在面临“岩溶性”干旱时,植物解剖结构的适应策略不同,树干液流密度主要受环境因子的调控。
3.3 喀斯特森林主要树种抗旱能力评价
树种的抗旱能力随着导管直径的增大而增加,与张海昕[51]和张硕新等[52]的研究结果一致,但也有研究认为被子植物的导管直径增大不利于植物在干旱情况下生存[53],与本研究结果相反,这是由于喀斯特地区植物的木质部导管直径兼具“二元性”,即植物木质部同时具有粗大的导管和细小的导管,能兼具“高安全性”和“高效率”的水分运输方式[54]。本研究中抗旱能力较强的南酸枣、山乌桕、化香等树种同时具有宽、窄两种导管,因此其抗旱能力比新木姜子、鹅耳枥等仅具有小导管(导管直径在10~30 μm的超过60%)的树种强,周朝彬等[55]的研究也证明这一结论。本研究表明落叶树种抗旱能力强于常绿树种,这与落叶树种导水能力、输水能力均强于常绿树种结果相一致,说明落叶树种在长期适应自然环境的过程中,形成了一套最适宜自身生长发育的生理生态行为来抵御或忍耐干旱胁迫的影响[56]。
综上,在茂兰喀斯特原生林中,9个树种的导水能力、输水能力、胡伯尔值和解剖结构存在一定差异,且在不同的种间表现各异。落叶树种的导水、输水、抗旱能力均强于常绿树种。冠层树种通过增大单位横截面积上的蒸腾拉力抵消树高带来水分运输阻力,为冠层叶片蒸腾和光合作用供给水分。在水分充足的条件下,水力学性状对茂兰喀斯特原生林树种树干液流密度的影响较小,而解剖结构对树干液流密度的影响存在种间差异。为此,建议在进行喀斯特植被恢复时应综合考虑树木的生活习性、林分结构配比。另外,由于条件限制,本研究在小生境和树种的选择上不够全面,未来将进一步扩大研究对象的样本数,以获取更具普遍性的结论。
参考文献(reference):
[1]HUNTINGTON T G.Evidence for intensification of the global water cycle:review and synthesis[J].J Hydrol,2006,319(1/2/3/4):83-95.DOI: 10.1016/j.jhydrol.2005.07.003.
[2]TYREE M T,SPERRY J S.Vulnerability of xylem to cavitation and embolism[J].Annu Rev Plant Physiol Plant Mol Biol,1989,40:19-36.DOI: 10.1146/annurev.pp.40.060189.000315.
[9]洪震,刘术新,洪琮浩,等.5种造林树种对干旱胁迫的抗性应答[J].南京林业大学学报(自然科学版),2021,45(2):111-119.HONG Z,LIU S X,HONG C H,et al.Resistance response of five afforestation tree species under drought stress[J].J Nanjing For Univ (Nat Sci Ed),2021,45(2):111-119.DOI: 10.11833/j.issn.2095-0756.2016.04.012.
[4]JUMP A S,RUIZ-BENITO P,GREENWOOD S,et al.Structural overshoot of tree growth with climate variability and the global spectrum of drought-induced forest dieback[J].Glob Chang Biol,2017,23(9):3742-3757.DOI: 10.1111/gcb.13636.
[5]CHOAT B,JANSEN S,BRODRIBB T J,et al.Global convergence in the vulnerability of forests to drought[J].Nature,2012,491(7426):752-755.DOI: 10.1038/nature11688.
[6]刘娟娟,张建国,李吉跃,等.CO2浓度升高对4种乔木幼树水力结构的影响[J].生态学杂志,2017,36(7):1769-1776.LIU J J,ZHANG J G,LI J Y,et al.Influences of elevated CO2 concentration on hydraulic architecture of seedlings of 4 tree species[J].Chin J Ecol,2017,36(7):1769-1776.DOI: 10.13292/j.1000-4890.201707.037.
[7]郑茹萍,张马啸,吴亚岚,等.杉木不同形态特征对水力结构的影响[J].森林与环境学报,2023,43(1):17-25.ZHENG R P,ZHANG M X,WU Y L,et al.Effects of different morphological characteristics on hydraulic architecture of Chinese fir[J].J For Environ,2023,43(1):17-25.DOI: 10.13324/j.cnki.jfcf.2023.01.003.
[8]李吉跃,翟洪波.木本植物水力结构与抗旱性[J].应用生态学报,2000,11(2):301-305.LI J Y,ZHAI H B.Hydraulic architecture and drought resistance of woody plants[J].Chin J Appl Ecol,2000,11(2):301-305.DOI: 10.13287/j.1001-9332.2000.0079.
[9]STEPPE K,VANDEGEHUCHTE M W,TOGNETTI R,et al.Sap flow as a key trait in the understanding of plant hydraulic functioning[J].Tree Physiol,2015,35(4):341-345.DOI: 10.1093/treephys/tpv033.
[10]OREN R,PHILLIPS N,EWERS B E,et al.Sap-flux-scaled transpiration responses to light,vapor pressure deficit,and leaf area reduction in a flooded Taxodium distichum forest[J].Tree Physiol,1999,19(6):337-347.DOI: 10.1093/treephys/19.6.337.
[11]龙翠玲.不同地形部位喀斯特森林物种多样性的比较研究:以贵州茂兰自然保护区为例[J].中国岩溶,2007,26(1):55-60.LONG C L.Comparison of species diversity in Karst forest among different topography sites: a case study in Maolan Natural Reserve,Guizhou Province[J].Carsologica Sin,2007,26(1):55-60.DOI: 10.3969/j.issn.1001-4810.2007.01.009.
[12]朱秀勤.石林溶丘洼地区不同恢复阶段植物水分利用的稳定同位素研究[D].昆明:云南师范大学,2014.ZHU X Q.Study on stable isotopes of plant water use in different restoration stages in Rrongqiuwa area of Shilin[D].Kunming:Yunnan Normal University,2014.
[13]梁千慧.不同水分条件下黑麦草对喀斯特土壤深度的生长与生理响应[D].重庆:西南大学,2016.LIANG Q H.Growth and physiological response of Lolium perenne to Karst soil depth under different water conditions[D].Chongqing:Southwest University,2016.
[14]张启伟.北热带喀斯特山地不同坡位木本植物的水力结构与功能[D].南宁:广西大学,2021. ZHANG Q W.Hydraulic structure and function of woody plants at different slope positions in northern tropical Karst mountainous areas[D].Nanning:Guangxi University,2021.DOI: 10.27034/d.cnki.ggxiu.2021.000052.
[15]程娟,丁访军,谭正洪,等.贵州茂兰喀斯特森林两树种叶片气孔形态特征及其对蒸腾的影响[J].南京林业大学学报(自然科学版),2021,45(5):125-132.CHENG J,DING F J,TAN Z H,et al.Leaf stomatal morphological characteristics and their effects on transpiration for two tree species in Maolan Karst area,Guizhou Province[J].J Nanjing For Univ (Nat Sci Ed),2021,45(5):125-132.DOI: 10.12302/j.issn.1000-2006.202010005.
[16]袁丛军,程娟,丁访军,等.喀斯特森林优势树种蒸腾特征及其对气孔和气象因子的响应[J].中南林业科技大学学报,2022,42(7):96-105.YUAN C J,CHENG J,DING F J,et al.Transpiration characteristics of dominant tree species and their responses to stomatal and meteorological factors in Karst regions[J].J Cent South Univ For Technol,2022,42(7):96-105.DOI: 10.14067/j.cnki.1673-923x.2022.07.011.
[17]李成龙,刘延惠,丁访军,等.茂兰喀斯特森林小果润楠蒸腾特征及影响因素[J].南京林业大学学报(自然科学版),2019,43(3):51-58.LI C L,LIU Y H,DING F J,et al.Transpiration characteristics and influencing factors of the dominant species of Machilus microcarpa in Maolan Karst forest[J].J Nanjing For Univ (Nat Sci Ed),2019,43(3):51-58.DOI: 10.3969/j.issn.1000-2006.201808018.
[18]吴鹏,杨文斌,崔迎春,等.喀斯特区天峨槭(Acer wangchii)树干液流特征及其与环境因子的相关分析[J].生态学报,2017,37(22):7552-7567.WU P,YANG W B,CUI Y C,et al.Characteristics of sap flow and correlation analysis with environmental factors of Acer wangchii in the Karst area[J].Acta Ecol Sin,2017,37(22):7552-7567.DOI: 10.5846/stxb201609251934.
[19]GRANIER A.Evaluation of transpiration in a Douglas-fir stand by means of sap flow measurements[J].Tree Physiol,1987,3(4):309-320.DOI: 10.1093/treephys/3.4.309.
[20]樊大勇, 谢宗强, 赵常明, 等. 一种测定木本植物最大水力导度的装置及其使用方法[P]. 2015-11-11. FAN D Y, XIE Z Q, ZHAO C M. A device and method for measuring the maximum hydraulic conductivity of woody plants[P]. 2015-11-11.
[21]熊仕发,吴立文,陈益存,等.不同种源白栎幼苗叶片对干旱胁迫的响应及抗旱性评价[J].生态学杂志,2020,39(12):3924-3933.XIONG S F,WU L W,CHEN Y C,et al.Response of leaf of Quercus fabri seedlings from different provenances to drought stress and drought resistance evaluation[J].Chin J Ecol,2020,39(12):3924-3933.DOI: 10.13292/j.1000-4890.202012.022.
[22]车路平. 14个造林树种木质部、叶片解剖特征及抗旱性评价[D]. 泰安: 山东农业大学, 2022. CHE L P. Anatomical characteristics and drought resistance evaluation of xylem and leaves of 14 afforestation tree species[D]. Taian: Shandong Agricultural University, 2022.
[23]FAN D Y,JIE S L,LIU C C,et al.The trade-off between safety and efficiency in hydraulic architecture in 31 woody species in a Karst area[J].Tree Physiol,2011,31(8):865-877.DOI: 10.1093/treephys/tpr076.
[24]赵延涛,许洺山,张志浩,等.浙江天童常绿阔叶林不同演替阶段木本植物的水力结构特征[J].植物生态学报,2016,40(2):116-126.ZHAO Y T,XU M S,ZHANG Z H,et al.Hydraulic architecture of evergreen broad-leaved woody plants at different successional stages in Tiantong National Forest Park,Zhejiang Province,China[J].Chin J Plant Ecol,2016,40(2):116-126.DOI: 10.3724/SP.J.1258.2014.00078.
[25]王佳敏,成应杰,陈金艺,等.模拟不同降雨时间格局下喀斯特垂直异质生境对桢楠幼苗光合和生长的影响[J].生态学报,2021,41(18):7348-7356.WANG J M,CHENG Y J,CHEN J Y,et al.Effects of simulated Karst vertical heterogeneous habitat on photosynthesis and growth of Phoebe zhennan S.Lee seedlings under different rainfall temporal pattern[J].Acta Ecol Sin,2021,41(18):7348-7356.DOI: 10.5846/stxb202007131824.
[26]李吉跃,翟洪波,刘晓燕.树木水力结构特征的昼夜变化规律[J].北京林业大学学报,2002,24(4):39-44.LI J Y,ZHAI H B,LIU X Y.Day and night change of tree hydraulic architecture characteristics[J].J Beijing For Univ,2002,24(4):39-44.DOI: 10.3321/j.issn:1000-1522.2002.04.009.
[27]黄恺翔,俞重阳,钱海蓉,等.鸡公山国家级自然保护区散孔材、环孔材树种木质部结构和功能的关系[J].浙江农林大学学报,2022,39(2):244-251.HUANG K X,YU C Y,QIAN H R,et al.Relationship between xylem structure and function of diffuse-porous and ring-porous wood species in Jigongshan Nature Reserve[J].J Zhejiang A F Univ,2022,39(2):244-251.DOI: 10.11833/j.issn.2095-0756.20210628.
[28]PARENDS L A, EWEL K C. Usefulness of annual growth rings of cypress trees (Taxodium distichum) for impact analysis[J]. Tree-Ring Bulletin, 1984(44): 39-43.
[29]CANNY M.Vessel contents during transpiration: embolisms and refilling[J].Am J Bot,1997,84(9):1223.
[30]CHOAT B,BALL M C,LULY J G,et al.Hydraulic architecture of deciduous and evergreen dry rainforest tree species from north-eastern Australia[J].Trees,2005,19(3):305-311.DOI: 10.1007/s00468-004-0392-1.
[31]CHEN J W,ZHANG Q,CAO K F.Inter-species variation of photosynthetic and xylem hydraulic traits in the deciduous and evergreen Euphorbiaceae tree species from a seasonally tropical forest in south-western China[J].Ecol Res,2009,24(1):65-73.DOI: 10.1007/s11284-008-0482-4.
[32]谭凤森.桂西南北热带喀斯特季雨林木本植物的水力学特征研究[D].南宁:广西大学,2019.TAN F S.Study on hydraulic characteristics of woody plants in tropical Karst monsoon forest in northwest Guangxi[D].Nanning:Guangxi University,2019.
[33]刘晓静,赵平,王权,等.树高对马占相思整树水分利用的效应[J].应用生态学报,2009,20(1):13-19.LIU X J,ZHAO P,WANG Q,et al.Effects of tree height on whole-tree water use of Acacia mangium[J].Chin J Appl Ecol,2009,20(1):13-19.
[34]赵平,孙谷畴,倪广艳,等.成熟马占相思水力导度对水分利用和光合响应的季节性差异[J].应用生态学报,2013,24(1):49-56.ZHAO P,SUN G C,NI G Y,et al.Seasonal differences in the leaf hydraulic conductance of mature Acacia mangium in response to its leaf water use and photosynthesis[J].Chin J Appl Ecol,2013,24(1):49-56.DOI: 10.13287/j.1001-9332.2013.0126.
[35]倪鸣源.中亚热带喀斯特天然林树种枝条木质部性状与生长速率的相关性研究[D].南宁:广西大学,2021. NI M Y.Correlation between xylem characteristics and growth rate of tree branches in Karst natural forest in central subtropical zone[D].Nanning:Guangxi University,2021.DOI: 10.27034/d.cnki.ggxiu.2021.001287.
[36]HACKE U G,SPICER R,SCHREIBER S G,et al.An ecophysiological and developmental perspective on variation in vessel diameter[J].Plant Cell Environ,2017,40(6):831-845.DOI: 10.1111/pce.12777.
[37]樊大勇,谢宗强.木质部导管空穴化研究中的几个热点问题[J].植物生态学报,2004,28(1):126-132.FAN D Y,XIE Z Q.Several controversial viewpoints in studying the cavitation of xylem vessels[J].Acta Phytoecol Sin,2004,28(1):126-132.
[38]陈昭.润楠属枝条和叶片的水力特征及协同关系[D].南宁:广西大学,2018.CHEN Z.Hydraulic characteristics and synergistic relationship of branches and leaves of Machilus[D].Nanning:Guangxi University,2018.
[39]陈模舜,柯世省,杨勇宇,等.珍稀濒危植物天台鹅耳枥营养器官的解剖学研究[J].浙江林业科技,2010,30(5):14-19.CHEN M S,KE S S,YANG Y Y,et al.Anatomical study on vegetative organs of Carpinus tientaiensis[J].J Zhejiang For Sci Technol,2010,30(5):14-19.DOI: 10.3969/j.issn.1001-3776.2010.05.004.
[40]韩丽娟,周春丽,吴树明.国产胡桃科次生木质部导管分子的比较解剖及其系统位置的讨论[J].西北植物学报,2002,22(6):146-151,296.HAN L J,ZHOU C L,WU S M.Comparative anatomical studies on the vessel elements in the secondary xylem of Juglandaceae from China and discussion on their evolutionary position[J].Acta Bot Boreali Occidentalia Sin,2002,22(6):146-151,296.DOI: 10.3321/j.issn:1000-4025.2002.06.024.
[41]张瑞婷,杨金艳,阮宏华.树干液流对环境变化响应研究的整合分析[J].南京林业大学学报(自然科学版),2022,46(5):113-120.ZHANG R T,YANG J Y,RUAN H H.Meta-analyses of responses of sap flow to changes in environmental factors[J].J Nanjing For Univ (Nat Sci Ed),2022,46(5):113-120.DOI: 10.12302/j.issn.1000-2006.202101029.
[42]王连春.酸枣荆条液流特征及其耗水尺度扩展研究[D].北京:北京林业大学,2013.WANG L C.Study on the characteristics of the sap flow of Vitex spinosa and the expansion of its water consumption scale[D].Beijing:Beijing Forestry University,2013.
[43]袁星明.湘西石漠化地区不同生境下三种乔木的树干液流特征[D].长沙:中南林业科技大学,2022.DOI: 10.27662/d.cnki.gznlc.2022.000075.YUAN X M.Characteristics of sap flow of three kinds of trees in different habitats in rocky desertification areas of western Hunan[D].Changsha:Central South University of Forestry amp; Technology,2022.DOI: 10.27662/d.cnki.gznlc.2022.000075.
[44]宋波,董豪,全飞,等.橡胶树干CO2释放速率垂直变化及其与树干液流和木质部结构间的关系[J].热带生物学报,2022,13(1):1-6.SONG B,DONG H,QUAN F,et al.Vertical variation of stem CO2 efflux and its relationship with sap flow and xylem structure of rubber trees(Hevea brasiliensis)[J].J Trop Biol,2022,13(1):1-6.DOI: 10.15886/j.cnki.rdswxb.2022.01.001.
[45]王秀伟,毛子军.输导组织结构对液流速度和树干CO2释放通量的影响[J].北京林业大学学报,2013,35(4):9-15.WANG X W,MAO Z J.Effects of conducting tissue structure on sap flow density and stem CO2 efflux[J].J Beijing For Univ,2013,35(4):9-15.DOI: 10.13332/j.1000-1522.2013.04.016.
[46]刘春鹏,马长明,王连春,等.酸枣荆条耗水特征及其茎木质部解剖构造[J].水土保持通报,2017,37(5):92-97.LIU C P,MA C M,WANG L C,et al.Water-consumption characteristics and stem xylem anatomical structure of Zizyphus jujuba and Vitex negundo var.heterophylla[J].Bull Soil Water Conserv,2017,37(5):92-97.DOI: 10.13961/j.cnki.stbctb.2017.05.016.
[47]赵培强,赵平,牛俊峰,等.华南地区常见树种导管特征与树干液流的关系[J].热带亚热带植物学报,2014,22(6):537-548.ZHAO P Q,ZHAO P,NIU J F,et al.Relationship between vessel characteristics and sap flow of eight subtropical tree species[J].J Trop Subtrop Bot,2014,22(6):537-548.DOI: 10.11926/j.issn.1005-3395.2014.06.001.
[48]方菁,叶琳峰,陈森,等.自然和人工生境被子植物枝木质部结构与功能差异[J].植物生态学报,2021,45(6):650-658.FANG J,YE L F,CHEN S,et al.Differences in anatomical structure and hydraulic function of xylem in branches of angiosperms in field and garden habitats[J].Chin J Plant Ecol,2021,45(6):650-658.DOI: 10.17521/cjpe.2020.0430.
[49]董彦君,李宗善,陈颖,等.黄耆属草本植物根部导管结构对环境因子的响应[J].生态学杂志,2023,42(12):2936-2943.DONG Y J,LI Z S,CHEN Y,et al.Effects of environmental factors on root vessel anatomical traits of Astragalus herbs[J].Chin J Ecol,2023,42(12):2936-2943.DOI: 10.13292/j.1000-4890.202312.012.
[50]张新英,曹宛虹.生长在不同生境下的沙棘次生木质部解剖学的研究[J].植物学报,1990,32(12):909-915,993-994.ZHANG X Y,CAO W H.Studies on the secondary xylem anatomy of Hippophae rhamnoides under different habitats[J].J Integr Plant Biol,1990,32(12):909-915,993-994.
[51]张海昕.几个杨属无性系木质部导管结构与栓塞脆弱性研究[D].杨凌:西北农林科技大学,2013.ZHANG H X.Study on vessel structure and embolism vulnerability of xylem of several Populus clones[D].Yangling:Northwest A amp; F University,2013.
[52]张硕新, 申卫军, 张远迎, 等. 几个抗旱树种木质部栓塞脆弱性的研究[J]. 西北林学院学报, 1997,12(2): 1-6. ZHANG S X, SHEN W J, ZHANG Y Y, et al. The vulnerability of xylem embolism in twigs of some drought-resistent tree species[J]. Journal of Northwest Forestry College, 1997,12(2): 1-6. DOI:CNKI:SUN:XBLX.0.1997-02-000.
[53]艾绍水,李秧秧,陈佳村,等.陕北沙地3种典型灌木根木质部解剖结构及水力特性[J].应用生态学报,2015,26(11):3277-3284.AI S S,LI Y Y,CHEN J C,et al.Root anatomical structure and hydraulic traits of three typical shrubs on the sandy lands of northern Shaanxi Province,China[J].Chin J Appl Ecol,2015,26(11):3277-3284.DOI: 10.13287/j.1001-9332.20150921.045.
[54]ZHU S D,CHEN Y J,FU P L,et al.Different hydraulic traits of woody plants from tropical forests with contrasting soil water availability[J].Tree Physiol,2017,37(11):1469-1477.DOI: 10.1093/treephys/tpx094.
[55]周朝彬,辛慧慧,宋于洋.梭梭次生木质部解剖特征及其可塑性研究[J].西北林学院学报,2014,29(2):207-212.ZHOU C B,XIN H H,SONG Y Y.Secondary xylem anatomical structure and its plasticity of Haloxylon ammodendron[J].J Northwest For Univ,2014,29(2):207-212.DOI: 10.3969/j.issn.1001-7461.2014.02.37.
[56]刘娟娟,李吉跃,王继强.北京城市绿化树种的水力结构特征[J].北京林业大学学报,2006,28(S1):38-46.LIU J J,LI J Y,WANG J Q.Hydraulic architecture characteristics of urban afforestation trees in Beijing[J].J Beijing For Univ,2006,28(S1):38-46.
[56]刘娟娟,李吉跃,王继强.北京城市绿化树种的水力结构特征[J].北京林业大学学报,2006,28(S1):38-46.LIU J J,LI J Y,WANG J Q.Hydraulic architecture characteristics of urban afforestation trees in Beijing[J].J Beijing For Univ,2006,28(S1):38-46. DOI:10.3969/j.issn.1000-2006.2008.06.032.
(责任编辑 孟苗婧 郑琰燚)
