
无患子生殖生长物候期液流特征及其对枝叶修剪的响应
2024-12-12 00:00:00王冕之郑玉琳贾黎明李露罗水晶刘济铭刘俊涛
南京林业大学学报(自然科学版) 2024年6期




摘要:【目的】了解无患子(Sapindus saponaria)关键物候期液流速率特征及其对修剪和气象因子的响应规律,为无患子生殖生长各关键物候期水分的科学管理提供理论参考。【方法】在福建省三明市建宁县无患子国家林木种质资源库试验区,以5年生无患子新品种‘媛华’为试验材料,利用热扩散液流探针和自动气象站监测2021年4月20日—11月9日的树干液流和气象数据分析气象因子和枝叶修剪对无患子液流特征的影响。试验中探针安装于距地面约30 cm处以保证所有处理的探针都安装于第1分枝之下。修剪组样树在花序抽生期长出花序后进行一次性修剪,去掉遮蔽花序阳光的枝组和复叶。对照组样树不做修剪处理。【结果】①无患子日均液流速率随时间在7个生殖生长关键物候期尺度上呈现降低—升高—降低的趋势。在初果期蒸腾活动最强,日均液流速率高达(1.13±0.05)×10-3(修剪)和(1.48±0.05)×10-3cm/s(对照),在果实成熟期蒸腾活动最弱,日均液流速率低至(0.15±0.02)×10-3(修剪)和(0.26±0.03)×10-3 cm/s(对照)。除花芽膨大期外,修剪显著降低了无患子各物候期液流速率,降低作用在树体生长旺盛的晴天较明显、在果实发育末期最强。②太阳辐射、空气温度、风速和饱和水汽压亏缺是无患子液流速率的主要驱动因子,而降水量和空气相对湿度是主要限制因子。持续降水等气象因素是花芽膨大期至开花期前期和果实膨大期中期无患子蒸腾活动的主要限制因素,叶片老化、树体逐渐进入休眠是果实成熟期液流速率低的主要原因。【结论】修剪显著降低了树体蒸腾作用,降低效应随关键物候期向后发展而增强。无患子生殖生长全物候期液流速率波动明显,推测树木和果实生长对水分的需求与树体耗水行为密切相关,而花序生长发育时期持续过量降雨等气象因素不利于无患子蒸腾耗水。
关键词:无患子;液流;修剪;物候期;气象因子
中图分类号:S725.5"""""" 文献标志码:A开放科学(资源服务)标识码(OSID):
文章编号:1000-2006(2024)06-0013-10
The" sap flow characteristics and responses to branch and leaf pruning during reproductive phenological periods in Sapindus saponaria
WANG Mianzhi1, ZHENG Yulin1, JIA Liming1*, LI Lu1, LUO Shuijing2, LIU Jiming1, LIU Juntao1
(1.National Energy R amp; D Center for Non-Food Biomass, Key Laboratory for Silviculture and Conservation of Ministry of Education, College of Forestry," Beijing Forestry University, Beijing 100083, China; 2. Fujian Yuanhua Forestry Biotechnology Co., Ltd., Jianning 354500, China)
Abstract: 【Objective】The present study aimed to investigate the characteristics of sap flow velocity of Sapindus saponaria"" and its response pattern to pruning and meteorological factors during the key phenological periods, and" to provide a theoretical reference for scientific water management during key reproductive phenological periods of Sapindus.【Method】The present study was conducted in the experimental area of the Sapindus National Forest Germplasm Repository in Jianning County, Sanming City, Fujian Province. Using five-year-old" S. saponaria ‘Yuanhua’. Thermal dissipation probes (TDPs) and automatic weather stations were utilized to monitor sap flow and meteorological data from April 20, 2021 to November 9, 2021. The probes were installed approximately 30 cm above the ground to ensure that all probes were installed under the first branch. Sample trees in the pruning group were pruned once after inflorescence growth at the inflorescence growth period to remove branches and leaves that shaded the inflorescence from sunlight. Trees in the control group were left unpruned for comparison.【Result】The average daily sap flow velocity of S. saponaria showed a decreasing-increasing-decreasing trend over time during the seven key reproductive phenological periods. The strongest transpiration activities were (1.13±0.05)×10-3 cm/s (pruning) and (1.48±0.05)×10-3 cm/s (CK) in the early ovary growing period, while the weakest transpiration activities were (0.15±0.02)×10-3 cm/s (pruning) and (0.26±0.03)×10-3 cm/s (CK) in the fruit maturity period. Pruning significantly reduced the sap flow velocity of S. saponaria in all phenological periods, except for the bud swelling period. This reduction effect was more pronounced on sunny days when the tree was actively growing and was" strongest at the end of the fruit development period. In addition, solar radiation, air temperature, wind speed" and vapor pressure deficit (VPD) were the main drivers of sap flow velocity, while precipitation and air humidity were the main limiting factors. Meteorological factors, such as persistent rainfall, were the main limitations of the" transpiration activity of S. saponaria from the bud swelling period to the early flowering period and middle fruit development period. Leaf aging and gradual dormancy of the tree were the main reasons for low sap flow velocity during the fruit maturity period.【Conclusion】Pruning significantly reduced the transpiration, with the reduction effect becoming more pronounced as the phenological periods progressed. There were large fluctuations in sap flow velocity throughout the reproductive growth of S. saponaria. Thus, these findings suggested that the water demand for tree and fruit growth is closely related to the water consumption behavior of" trees. Meteorological factors, such as persistent excessive rainfall, during inflorescence growth and development are not conducive to transpiration.
Keywords:Sapindus saponaria; sap flow; pruning; phenological period; meteorological factor
树干液流是植物水力学特性的重要指标,受到气象因子、植物生物学特性和营林措施的共同作用,反映了植物的蒸腾耗水状况。太阳辐射[1]、空气温湿度[2]、土壤水热条件[3-4]和空气流通速率[5-6]等气象因子相互作用,共同影响着植物的蒸腾耗水过程。在相同立地条件下,不同的植物往往有其独特的液流规律[7],而同一植物在不同的年龄和物候期也会因为蒸腾和需水规律的不同而表现出不同的液流特征[8-10]。
无患子(Sapindus saponaria)广泛分布于中国南部、印度、中南半岛、日本和朝鲜半岛[11]。因其在生物日化、生物柴油、生物医药和生态修复领域的价值而受到广泛关注[12-15],现已初步建立无患子多联产高效产业链模式[15]。无患子作为喜光树种,其下层花序(果序)易受枝叶遮挡,光照不足,导致果实产量和品质降低,在生产实践中常通过修剪改善无患子树冠光环境,提高无患子果实产量和品质[16-19]。同时,修剪还会改变林木的耗水情况与水分利用效率[20-21],从而影响植株的营养生长和生殖生长状况[20, 22-24]。因此,了解修剪措施对植株水力学特征的影响对经济林抚育管理与林木水分高效利用具有重要意义,但目前鲜见研究探索修剪对无患子液流特性影响规律的报道。此外,植株在不同物候期的耗水规律不同[10, 25],其蒸腾和液流特性对气象因子的敏感性和响应规律也不同[8-9],了解植株各物候期的蒸腾特征及其对气象因子的响应规律可以为其水分管理提供科学理论参考,但目前有关无患子不同生殖生长物候期液流特征及无患子不同物候期液流特征对气象因子响应规律的研究报道较少。
因此,为了探索遮蔽花序枝叶修剪措施对无患子液流特性的影响、并为无患子生殖生长各关键物候期水分的科学管理提供理论参考,以5年生无患子新品种‘媛华’(‘Yuanhua’)试验林为实验材料,结合热扩散液流探针和自动气象站,采集了无患子修剪后各关键物候期的液流和气象因子数据。主要解决以下几个问题:①无患子生殖生长关键物候期液流速率有何特征?如何变化?②修剪对无患子液流速率有何影响?③无患子各物候期液流速率对气象因子有何响应规律?
1 材料与方法
1.1 试验地概况与试验材料
试验地位于福建省三明市建宁县无患子国家种质资源库(116°35′~117°04′E,26°30′~27°06′N),气候特征为中亚热带海洋性季风气候,并具有大陆性山地气候特点,年均气温17.64 ℃,年均降水量1 950 mm。土壤为长石砂岩风化母质上发育成的山地黄红壤,pH 5.02,土壤肥力状况为中等偏下[26]。
研究对象为5年生‘媛华’嫁接苗示范林,为2016年嫁接到2年生无患子实生苗砧木上而成,试验林地立地条件基本一致,株行距4 m × 4 m,栽植密度为635株/hm2[26]。试验林地采用统一的营林管护措施:全年除草4次,5—8月每月除草1次;全年杀虫5次,4、6和9月每月喷施杀虫剂1次,7月喷施杀虫剂两次;全年施肥3次,在4、7和11月施肥,每株施尿素0.35 kg、过磷酸钙1.10 kg、氯化钾0.18 kg。
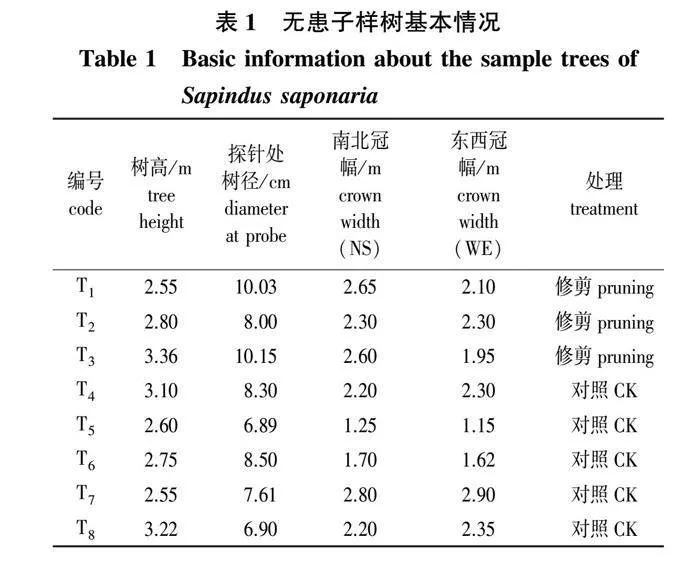
本试验在示范林中选取了3株样树,在2021年4月26日(花序抽生期)长出花序后进行一次性修剪,去掉遮蔽花序阳光的枝组和复叶,修剪强度以保证被剪样树花序无光线遮挡为准,各待修剪样树的修剪量保持一致,枝叶修剪量约占树冠总枝叶量的1/5;选择5株样树不做修剪为对照。样树基本情况见表1。
1.2 测定指标与方法
1.2.1 物候观测
将无患子生殖生长的关键物候期分为7个阶段:花序抽生期(inflorescence growth period,IG)、花芽膨大期(bud swelling period,BS)、开花期(flowering period,FP)、初果期(early ovary growing period,EOG)、果实膨大期(fruit development period,FD)、果实转色期(fruit colour change period,FCC)、果实成熟期(fruit maturity period,FM)[27-28]。连续观测8株样树的物候期,每次随机观测树冠4个方向的1年生枝共20个,当有15个1年生枝出现某一物候特征时视为达到该物候期[27]。同时采用日期和积日记录物候期,积日表示为一年中的日次序,以1月1日为积日1,积日值每天增加1[29]。
1.2.2 树干液流
于2021年3月7日安装热扩散液流探针(TDP-10,北京时域通科技有限公司),探针安装于距地面约30 cm处以保证所有处理的探针都安装于第1分枝之下。液流速率测定周期为10 s,数据每10 min计算1次平均值并保存于G12数据采集器(北京时域通科技有限公司)。液流速率计算公式[30]如下:
Fd=0.011 9ΔTm-ΔTΔT1.231。
(1)
式中:Fd为树干液流速率,cm/s;ΔTm为24 h内上下探针的最大温差, ℃;ΔT为某时刻加热探针和受热探针间的温差, ℃。
为消除可能因样树大小的个体差异对树干液流速率造成的误差,利用探针处树径大小对液流速率进行校正[31-32],公式如下:
Fd′=FdpDp。
(2)
式中:Fd′为校正后的液流速率,cm/s;p为所有样树探针处平均树径,cm;Dp为样树探针处树径,cm。
1.2.3 气象因子
在距离试验地20 m处安装自动气象站(北京时域通科技有限公司)实时监测气象因子。气象因子包括太阳总辐射(Rs,W/m2)、降水量(RG,mm)、风速(AWS,m/s)、大气压强(Pr,hPa)、空气温度(Ta, ℃)、空气相对湿度(RHa,式中记为HRa,%)、土壤温度(Ts, ℃)、土壤湿度(RHs,%)。气象因子与液流测定同步,其中土壤温度和土壤湿度为距离地面20、40、60和80 cm处的温湿度平均值。饱和水汽压亏缺(VPD,式中记为VPD, kPa)计算公式[33]如下:
VPD=0.611×e(17.502 TaTa+240.97)×(1-HRa)。
(3)
1.3 数据分析
使用Baseliner 4.21计算液流数据[34]。使用SPSS 24.0进行独立样本t检验(Student’s t test),以了解修剪组和对照组在各物候期的日均液流和日均最大液流速率是否存在显著差异(P lt; 0.05),使用Pearson相关性分析评价液流速率与气象因子的相关性。采用Origin 2022作图。
2 结果与分析
2.1 无患子液流速率特征
2.1.1 生殖生长全期液流速率特征
无患子‘媛华’植株自花序抽生期(积日110)开始,修剪组和对照组的液流速率变化总体呈波动状态,变化幅度较大(图1)。第1个液流速率低谷出现在花芽膨大期和开花期早期(积日138~152),该阶段修剪组和对照组平均液流速率相近,最低值至(0.45±0.07)×10-3cm/s;第2个液流速率低谷出现在果实膨大期中期(积日217~236),修剪组和对照组平均液流速率最低值分别为(0.48±0.05)×10-3和(0.91±0.16)×10-3 cm/s;第3个液流速率低谷出现在果实转色期末期和果实成熟期(积日287~313),修剪组和对照组平均液流速率最低值分别为(0.15±0.02)×10-3和(0.28±0.07)×10-3 cm/s。其中前两个液流速率低谷的形成伴随着持续降雨,而第3个低谷的产生伴随着树体落叶程度加强、逐渐进入休眠期。树体进入初果期和果实膨大前期时(积日161~202)出现第1个液流速率高峰,修剪组和对照组平均液流速率分别高达(1.20±0.09)×10-3和(1.52±0.09)×10-3 cm/s,该时期虽然有短暂的高降雨影响,但液流速率总体处于全时期的最高水平。第2个液流速率高峰出现在果实膨大期后期至果实转色期前期(积日243~282),修剪组和对照组平均液流速率分别高达(0.72±0.13)×10-3和(1.22 ± 0.21)×10-3 cm/s。
修剪组液流速率总体低于对照组(图1)。修剪前(积日110~115)待修剪样树和对照组日均液流速率无明显差异,表明样树水分状况接近,可比性强。从花序抽生期至开花期前期(积日116~155),修剪组和对照组日均液流速率差异较小,该时期伴随着持续降水。类似地,在初果期中期(积日179~182)出现了短暂持续降雨,该时期修剪组和对照组日均液流速率差异同样处于较低水平。但从开花期后期至果实转色期中期(积日156~283),修剪组和对照组日均液流速率差异逐渐增大,该阶段降水较少,反映了不同处理样树的水分耗散情况。直到进入果实转色期后期和果实成熟期(积日284~313)后两者差异逐渐下降。可见,修剪降低了无患子的蒸腾速率,减少了树体整体的蒸腾耗水,且降低效应在生长旺盛的晴天更明显。
2.1.2 各物候期液流速率特征
总体来说,修剪组与对照组的各关键物候期日均液流速率和日均液流速率峰值呈现降低—升高—降低的趋势(图2)。日均液流速率从花序抽生期(修剪前)的(1.02±0.14)×10-3和(1.19±0.09)×10-3 cm/s(对照)下降至花芽膨大期的(0.42±0.08)×10-3(修剪)和(0.61±0.08)×10-3 cm/s(对照);在开花期、初果期逐渐升高,在初果期达到峰值[修剪(1.13±0.05)×10-3" cm/s,对照(1.48±0.05)×10-3 cm/s];然后逐渐降低,在果实成熟期达到最低值[修剪(0.15±0.02)×10-3 cm/s,对照(0.26±0.03)×10-3 cm/s]。日均液流速率峰值与日均液流速率规律基本一致,但变化幅度更大(图2)。
修剪显著降低了各物候期的日均液流值(图2)。修剪前,待修剪样树和对照组日均液流特征无显著差异,样树间水分状况接近;修剪后,除花芽膨大期两者日均液流特征差异不显著外,其余物候期修剪样树日均液流速率和日均液流速率峰值均显著低于对照组(P lt; 0.05)。在花序抽生期、花芽膨大期、开花期、初果期、果实膨大期、果实转色期和果实成熟期修剪组日均液流速率较对照组下降了22.50%、30.94%、23.57%、23.10%、29.48%、45.15%、43.56%,日均液流速率峰值较对照组下降了17.96%、28.21%、28.08%、26.99%、34.11%、41.25%、45.16%。日均液流速率以果实转色期下降比例最大,日均液流速率峰值以果实成熟期下降比例为最大。
选取4月21日、5月14日、5月26日、6月9日、6月24日、8月11日、10月3日和10月30日分别代表花序抽生期(修剪前)、花序抽生期(修剪后)、花芽膨大期、开花期、初果期、果实膨大期、果实转色期和果实成熟期的典型晴天,用以分析各物候期典型晴天液流峰型、启动时间、高峰时间等特征。修剪没有改变无患子各物候期典型晴天的液流速率趋势,修剪和对照在各物候期典型晴天液流速率趋势基本一致,呈现“单峰性”曲线(图3)。不同物候期典型晴天的液流启动时间为6:30—9:20,其中花序抽生期(修剪后)(5月14日)启动时间最早(修剪和对照均为6:30),果实成熟期(10月30日)启动时间最晚(修剪为9:20,对照为9:10),但两种处理的样树液流启动时间的差异较小(小于10 min)。两种处理的液流速率高峰持续时长在部分物候期之间存在差异。其中,花序抽生期(修剪后)(5月14日)高峰时长最大,修剪和对照分别为9.17和10.33 h,而果实成熟期(10月30日)高峰时长最小,修剪和对照分别为2.50和2.33 h。表明修剪对无患子的液流启动时间和高峰时长影响较小,不同物候期之间的高峰时长差异较大,表现为生长旺盛时期高于生长初期和生长末期。二者在峰值上的差异规律与上述日均液流速率峰值基本一致。
2.2 液流速率与气象因子的关系
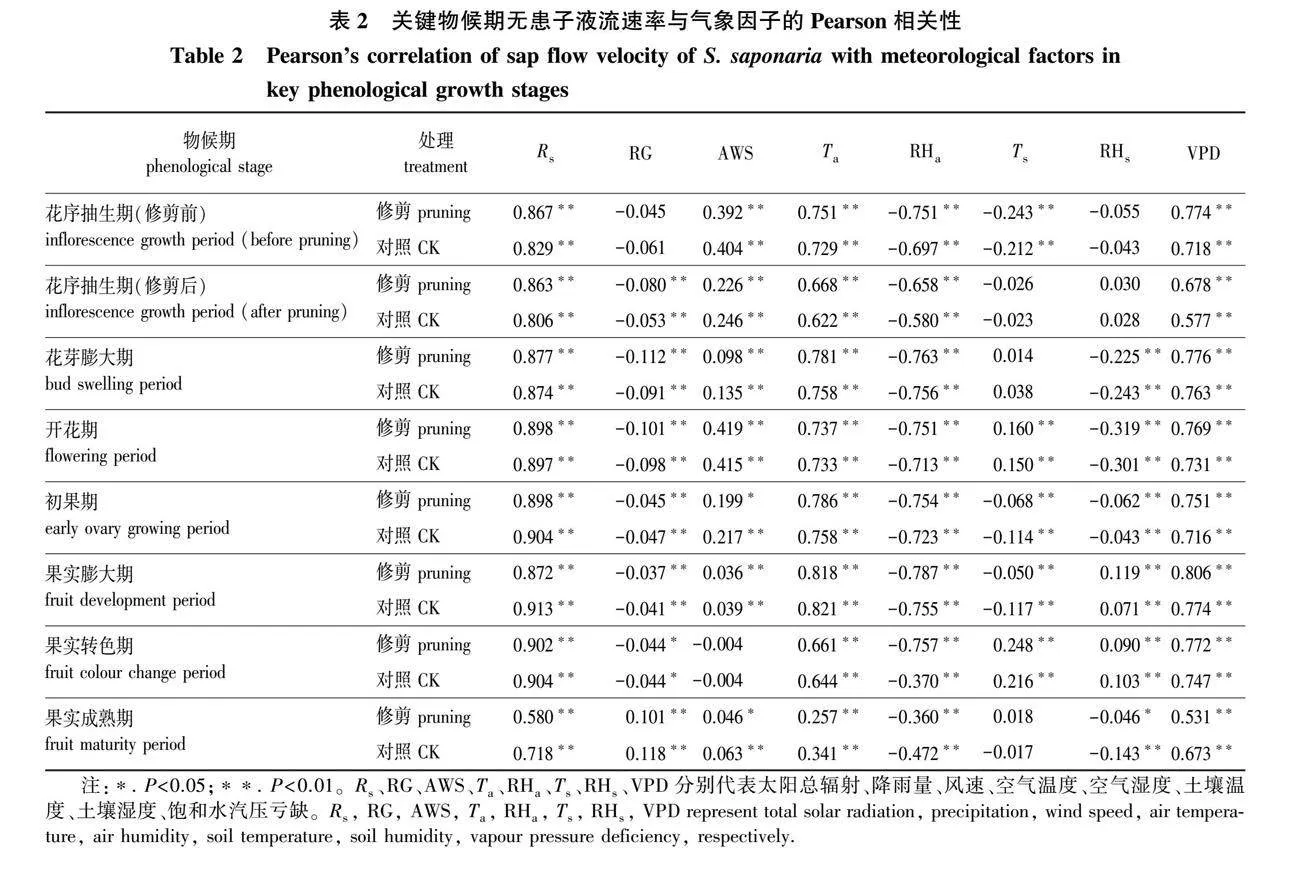
修剪组与对照组的液流速率对气象因子的响应基本一致,说明修剪没有显著影响气象因子对液流速率的驱动规律(表2)。Rs、Ta、RHa和VPD在7个物候期对液流速率的影响最稳定,其中Rs、Ta和VPD与液流速率呈极显著正相关关系(P lt; 0.01),而RHa则表现出极显著负相关关系(P lt; 0.01)。RG在花序抽生期(修剪前)与液流速率无显著相关,但从花序抽生期(修剪后)至果实转色期与液流速率呈现显著负相关关系(P lt; 0.05),而在果实成熟期表现出了极显著正相关关系(P lt; 0.01)。土壤温湿度在不同物候期对液流速率的影响不同,其中Ts在花序抽生期(修剪前)、初果期和果实膨大期与液流速率极显著负相关(P lt; 0.01),在开花期和果实转色期与液流速率极显著正相关(P lt; 0.01),而在花序抽生期(修剪后)、花芽膨大期和果实成熟期相关性不显著。RHs在果实膨大期和果实转色期与液流速率存在极显著正相关关系(P lt; 0.01),在花芽膨大期至初果期、果实成熟期表现为显著的负相关关系(P lt; 0.05),而在花序抽生期没有显著相关关系。
3 讨 论
3.1 关键生殖生长物候期液流速率的主要驱动与限制因素
无患子生殖生长全物候期液流速率呈现不断波动的变化趋势。尽管气象因子和树体自身的生长状态共同决定了无患子树干液流速率,但两者在不同时期的影响程度存在差异。在花芽膨大期和开花期早期(积日138~152)出现的第1个液流速率低谷与果实膨大期中期(积日217~236)出现的第2个液流速率低谷可能由气候因子主导。这两个时期存在持续高降雨量(RG),导致太阳总辐射(Rs)、空气温度(Ta)、土壤温度(Ts)、饱和水汽压亏缺(VPD)持续偏低,而RHa和RHs持续偏高,树干液流缺乏气象驱动力[35-36]。此外,在前两个液流速率低谷中,花芽膨大期和开花期早期液流速率低于果实膨大期中期,主要原因也是前者降雨量更大,对液流的气象驱动力更低。在果实转色期末期和果实成熟期(积日287~313)出现的第3个液流速率低谷则主要是无患子自身生长状态导致的,进入果实生长发育末期,树体逐渐进入休眠期,叶片老化、气孔导度降低[37],水力活动逐渐减弱。推测在初果期和果实膨大前期(积日161~202)出现的第1个液流速率高峰则主要是由树体自身主导的,此时虽然存在高降雨天气,但持续时间不长,未对液流速率产生较大影响,而树体已基本展叶完全[27-28],液流速率随着叶片等液流驱动器官的发育而提高[8, 38]。在果实膨大期后期至果实转色期前期(积日243~282)出现的第2个液流速率高峰则由气象因子主导,在此阶段降雨较少,液流的气象驱动力较高[35-36]。
降雨引起的蒸腾驱动力不足是花芽膨大期至初花期液流速率的主要限制因子。在花芽膨大期出现了持续高降雨,其中降雨天气占该物候期的80%,日降雨量高达92.6 mm,导致该物候期液流速率总体偏低,且液流速率降低趋势与降雨基本同步。然而花芽膨大期内的典型晴天液流速率处于较高水平,表明无患子在该时期可能存在较高蒸腾需求潜力。持续的蒸腾不足会影响植物营养运输[39],强降雨也可能引起水分胁迫,进而导致落花[40]。马长明等[41]也发现,核桃(Juglans regia)花期树体生命活动旺盛,液流速率处于较高水平,表明许多经济林木在花期存在较高的蒸腾需求。此外,高媛等[28]发现无患子初花期的落花等不良现象与降雨存在密切联系,进一步说明该时期无患子可能存在较高的蒸腾需求,而研究区域过量降雨引起的环境蒸腾驱动力降低不利于其开花坐果。
无患子在全物候期上表现为初果期和果实膨大初期蒸腾活动最强,果实成熟期蒸腾活动最弱。其他果树也存在类似的物候性蒸腾规律,例如孟秦倩[25]发现苹果在发育中期(果实膨大期)处于高耗水阶段,而在成熟期耗水降低;吴佳伟等[10]发现猕猴桃的耗水量从大到小表现为果实膨大期gt;果实成熟期gt;开花坐果期gt;萌芽展叶期;Hansen[42]发现结果数多的苹果耗水量更大,且高产果树气孔开放程度更高、蒸腾速率更大。这些结果表明果实生长发育初期树体蒸腾活动加剧、耗水增加。此外,郑玉琳等[43]发现无患子果实膨大后期果实含水率处于较高水平,而进入转色期末期时果实含水率骤降,直至果实成熟期含水率都处于低水平。本研究表明无患子树体液流速率也呈现出类似的变化规律,在果实膨大期后期至果实转色期前期液流速率处于较高水平,而在转色期末期液流速率骤降,而后在果实成熟期保持低水平。可见无患子果实对水分的需求与树体耗水间可能密切相关。
3.2 气象因子对无患子关键生殖生长物候期液流速率的影响
气象因子对修剪组和对照组无患子液流速率的影响基本一致。太阳总辐射(Rs)、空气温度(Ta)和饱和水汽压亏缺(VPD)在无患子各时期都与液流速率存在极显著正相关关系(P lt; 0.01),而RHa则表现为极显著负相关关系(P lt; 0.01)。这一结果与柽柳(Tamarix chinensis)[44]、毛白杨(Populus tomentosa)[29]、苹果(Malus domestica)[45-47]、猕猴桃(Actinidia chinensis)[10]和杏树(Prunus armeniaca)[48]的研究结果一致。例如,吴佳伟等[10]发现太阳有效辐射和气温在猕猴桃的萌芽展叶期、开花坐果期和果实膨大期均与猕猴桃的液流速率呈现极显著正相关关系,而空气湿度则呈现极显著负相关关系(P lt; 0.01)。说明Rs、Ta和VPD对许多乔木的影响相对稳定。相关性分析表明,大部分物候期降雨量(RG)都显著降低了无患子的液流速率(P lt; 0.05),这与荔枝(Litchi chinensis)等无患子科植物的响应规律基本一致[49]。风速(AWS)对无患子液流速率总体上呈现促进作用,仅在果实转色期对液流速率未产生显著影响。类似地,在前人研究中,AWS也常被视为液流速率的驱动因子[5, 46-47]。可能的原因是风能在一定程度上降低树叶温度,导致气孔水汽压降低,增大了其与饱和水汽压之间的差值,进而促进了植物的蒸腾作用[6]。Ts和RHs在不同时期对无患子液流速率的影响不同,且它们对液流速率的影响总体处于较低水平。土壤温度(Ts)对蒸腾作用的影响是复杂的,因为植物的生长状态和地面其他气象因素(如空气温度和日照时间等)都会影响土壤温度对植物蒸腾产生的作用[4]。例如,Mellander等[4]发现在约8 ℃以下时,植物叶片气孔导度和根系渗透性降低,此时土壤温度成为蒸腾作用的限制因素之一;而Petzold等[3]发现尽管土壤温度有利于林分蒸腾作用,但由于其他气象因素的干扰,蒸腾作用依旧降低。此外,吴桂伟等[10]也发现土壤湿度(RHs)对猕猴桃的液流速率影响在不同物候期存在差异:土壤湿度在萌芽展叶期、果实膨大期和果实成熟期对猕猴桃液流速率未产生显著影响,而在开花坐果期显著促进了其液流速率。表明土壤湿度在不同环境条件和植物的不同物候期对植物液流速率的影响不同。
3.3 修剪对无患子液流速率的影响
与未修剪的对照组相比,在花序抽生期修剪后,除花芽膨大期外,修剪处理显著降低了无患子所有物候期的日均液流速率和日均液流速率峰值。类似地,有报道称修剪措施降低了苹果、樱桃(Prunus pseudocerasus)和枣(Ziziphus jujuba)等经济林木的液流活动、蒸腾作用和树体耗水[20-21, 23]。Forrester等[22]发现修剪可以通过改变桉树人工林叶面积与边材面积之比和径向液流密度特征来改变树木的水力结构,进而降低了林分蒸腾量。这些结果说明修剪对叶片蒸腾普遍存在降低作用。
修剪对蒸腾的降低作用在晴天更明显,而在阴雨天作用较小,例如花芽膨大期的持续降雨限制了树体蒸腾,但修剪组和对照组之间液流速率差异较小。表明修剪在无患子花期缺乏蒸腾环境驱动力时不是其液流速率的主要限制因子。这一结果与Ma等[21]的研究结果不同,他们发现修剪对耗水的降低作用与降雨量存在正相关关系。造成结果差异的主要原因可能是地区气候特征的差异,Ma等[21]的研究区位于中国西北,属温带、半干旱气候带,平均年降水量为451.6 mm,长期的水分不足可能成为其蒸腾作用的主要限制因子,因此修剪组与对照组的耗水差异在降水相对充足的时期有了明显的上升;而本研究区域属于中亚热带海洋性季风气候,同时具有大陆性山地气候特点,年降水约为1 950 mm,水分供应不是无患子液流速率的主要限制因子,而降雨引起了液流的环境驱动力下降,进而使得修剪的影响不明显。
修剪对蒸腾的影响随着物候期的发展而逐渐增大,在果实发育后期(果实转色期和果实成熟期)对液流速率的降低比例最多。这一结果与Ma等[21]的研究结果类似,他们发现枣树从萌发抽叶期到果实成熟期的生长季内,修剪对蒸腾的降低作用随着物候期的发展而增大,在果实生长中后期降低作用最明显,但在果实成熟期末期差异减小。主要原因可能是随着叶片的生长发育,叶片蒸腾对液流速率的驱动效应逐渐增大[38],而修剪组冠层叶面积小于对照组,驱动效应相应减小,使得其液流速率差值逐渐增大。在果实成熟期后期,由于叶片老化导致的气孔导度降低和落叶等因素[37],修剪组和对照组液流速率差异显著降低。
无患子在各物候期典型晴天液流速率呈现“单峰性”曲线。生殖生长季液流速率波动较大,在生殖生长各关键物候期推进中表现为降低—升高—降低的趋势。花芽膨大期至开花期前期和果实膨大期中期的液流低谷是由降雨导致的蒸腾环境驱动力不足引起的,而持续降雨导致花期蒸腾不足、影响花果发育。树体叶片的成熟与老化是初果期至果实膨大期初期液流速率高峰和果实成熟期液流速率低谷的主要驱动因子,果实对水分的需求与树体耗水行为密切相关。修剪显著降低了花序抽生期、开花期、初果期、果实膨大期、果实转色期和果实成熟期的液流速率,对蒸腾的降低作用在晴天明显增强,在果实转色期和果实成熟期对液流速率的降低效应最大。太阳辐射、空气温度、风速和饱和水汽压亏缺是无患子液流速率的主要驱动因子,而降雨量和空气湿度是主要限制因子,气象因子之间相互关联、共同影响无患子各物候期的液流速率。
参考文献(reference):
[1]OOGATHOO S,HOULE D,DUCHESNE L,et al.Vapour pressure deficit and solar radiation are the major drivers of transpiration of balsam fir and black spruce tree species in humid boreal regions,even during a short term drought[J].Agric For Meteor,2020,291:108063.DOI: 10.1016/j.agrformet.2020.108063.
[2]HAN C,CHEN N,ZHANG C K,et al.Sap flow and responses to meteorological about the Larix principis-rupprechtii plantation in Gansu Xinlong Mountain,northwestern China[J].For Ecol Manag,2019,451:117519.DOI: 10.1016/j.foreco.2019.117519.
[3]PETZOLD R,SCHWRZEL K,FEGER K H.Transpiration of a hybrid poplar plantation in Saxony (Germany) in response to climate and soil conditions[J].Eur J For Res,2011,130(5):695-706.DOI: 10.1007/s10342-010-0459-z.
[4]MELLANDER P E,BISHOP K,LUNDMARK T.The influence of soil temperature on transpiration:a plot scale manipulation in a young Scots pine stand[J].For Ecol Manag,2004,195(1/2):15-28.DOI: 10.1016/j.foreco.2004.02.051.
[5]CHU C R,HSIEH C I,WU S Y,et al.Transient response of sap flow to wind speed[J].J Exp Bot,2009,60(1):249-255.DOI: 10.1093/jxb/ern282.
[6]DIXON M,GRACE J.Effect of wind on the transpiration of young trees[J].Ann Bot,1984,53(6):811-819.DOI: 10.1093/oxfordjournals.aob.a086751.
[7]IQBAL S,ZHA T S,JIA X,et al.Interannual variation in sap flow response in three xeric shrub species to periodic drought[J].Agric For Meteor,2021,297:108276.DOI: 10.1016/j.agrformet.2020.108276.
[8]LIU C W,DU T S,LI F S,et al.Trunk sap flow characteristics during two growth stages of apple tree and its relationships with affecting factors in an arid region of Northwest China[J].Agric Water Manag,2012,104:193-202.DOI: 10.1016/j.agwat.2011.12.014.
[9]GU D X,HE W,HUANG K C,et al.Transpiration of Moso bamboo in southern China is influenced by ramet age,phenology,and drought[J].For Ecol Manag,2019,450:117526.DOI: 10.1016/j.foreco.2019.117526.
[10]吴佳伟,李苇洁,杨瑞,等. 红阳猕猴桃生长发育期树干液流特征及其与环境因子的关系[J]. 果树学报,2021,39(3):1-25. WU G W,LI W J,YANG R,et al. Characteristics of trunk SAP flow and its relationship with environmental factors during growth and development of ‘Hongyang’ kiwifruit[J]. J Fruit Sci,2021,39(3):1-25. DOI:10.13925/j.cnki.gsxb.20210178.
[11]刘济铭,孙操稳,何秋阳,等.国内外无患子属种质资源研究进展[J].世界林业研究,2017,30(6):12-18.LIU J M,SUN C W,HE Q Y,et al.Research progress in Sapindus L. germplasm resources[J].World For Res,2017,30(6):12-18.DOI: 10.13348/j.cnki.sjlyyj.2017.0071.y.
[12]UPADHYAY A,SINGH D K.Molluscicidal activity of Sapindus mukorossi and Terminalia chebula against the freshwater snail Lymnaea acuminata[J].Chemosphere,2011,83(4):468-474.DOI: 10.1016/j.chemosphere.2010.12.066.
[13]MUNTAHA S T,KHAN M N.Natural surfactant extracted from Sapindus mukurossi as an eco-friendly alternate to synthetic surfactant:a dye surfactant interaction study[J].J Clean Prod,2015,93:145-150.DOI: 10.1016/j.jclepro.2015.01.023.
[14]CHAKRABORTY M,BARUAH D C.Production and characterization of biodiesel obtained from Sapindus mukorossi kernel oil[J].Energy,2013,60:159-167.DOI: 10.1016/j.energy.2013.07.065.
[15]刘诗琦,贾黎明,苏淑钗,等. 林业生物质能源“林油一体化”产业高效可持续发展路径研究[J]. 北京林业大学学报,2019,41(12):96-107. LIU S Q,JIA L M,SU S C,et al. Efficient and sustainable development path of forest-based bioenergy “forestry-oil integration” industry[J]. J Beijing Forestry Univ,2019,41(12):96-107. DOI:10.12171/j.1000-1522.20190433.
[16]张赟齐,刘晨,刘阳,等.叶幕微域环境对无患子果实产量和品质的影响[J].南京林业大学学报(自然科学版),2020,44(5):189-198.ZHANG Y Q,LIU C,LIU Y,et al.Effects of canopy micro-environment on fruit yield and quality characteristics of Sapindus mukorossi[J].J Nanjing For Univ (Nat Sci Ed),2020,44(5):189-198.DOI: 10.3969/j.issn.1000-2006.202001031.
[17]高媛,贾黎明,高世轮,等.无患子树体合理光环境及高光效调控[J].林业科学,2016,52(11):29-38.GAO Y,JIA L M,GAO S L,et al.Reasonable canopy light intensity and high light efficiency regulation of Sapindus mukorossi[J].Sci Silvae Sin,2016,52(11):29-38.DOI: 10.11707/j.1001-7488.20161104.
[18]ZHANG Y Q,WEN Y,BAI Q,et al.Spatio-temporal effects of canopy microclimate on fruit yield and quality of Sapindus mukorossi Gaertn[J].Sci Hortic,2019,251:136-149.DOI: 10.1016/j.scienta.2019.02.074.
[19]GAO Y,GAO S L,JIA L M,et al.Canopy characteristics and light distribution in Sapindus mukorossi Gaertn. are influenced by crown architecture manipulation in the hilly terrain of southeast China[J].Sci Hortic,2018,240:11-22.DOI: 10.1016/j.scienta.2018.05.034.
[20]MOLINA A J,ARANDA X,LLORENS P,et al.Sap flow of a wild cherry tree plantation growing under Mediterranean conditions:assessing the role of environmental conditions on canopy conductance and the effect of branch pruning on water productivity[J].Agric Water Manag,2019,218:222-233.DOI: 10.1016/j.agwat.2019.03.019.
[21]MA L H,WANG X,GAO Z Y,et al.Canopy pruning as a strategy for saving water in a dry land jujube plantation in a Loess Hilly Region of China[J].Agric Water Manag,2019,216:436-443.DOI: 10.1016/j.agwat.2018.12.007.
[22]FORRESTER D I,COLLOPY J J,BEADLE C L,et al.Effect of thinning,pruning and nitrogen fertiliser application on transpiration,photosynthesis and water-use efficiency in a young Eucalyptus nitens plantation[J].For Ecol Manag,2012,266:286-300.DOI: 10.1016/j.foreco.2011.11.019.
[23]LI K T,LAKSO A N,PICCIONI R,et al.Summer pruning reduces whole-canopy carbon fixation and transpiration in apple trees[J].J Hortic Sci Biotechnol,2003,78(6):749-754.DOI: 10.1080/14620316.2003.11511694.
[24]PINKARD E A,BEADLE C L,DAVIDSON N J,et al.Photosynthetic responses of Eucalyptus nitens (Deane and Maiden) Maiden to green pruning[J].Trees,1998,12(3):119-129.DOI: 10.1007/PL00009702.
[25]孟秦倩.黄土高原山地苹果园土壤水分消耗规律与果树生长响应[D].杨凌:西北农林科技大学,2011.MENG Q Q.Soil water consumption law and fruit tree growth response in apple orchards in mountainous areas of Loess Plateau[D].Yangling:Northwest A amp; F University,2011.
[26]刘俊涛,仲静,刘济铭,等.无患子初果期人工林土壤和叶片C、N、P化学计量特征[J].南京林业大学学报(自然科学版),2021,45(4):67-75.LIU J T,ZHONG J,LIU J M,et al.Stoichiometric characteristics of soil and leaves in Sapindus mukorossi plantation at an early fruiting stage[J].J Nanjing For Univ (Nat Sci Ed),2021,45(4):67-75.DOI: 10.12302/j.issn.1000-2006.202104011.
[27]ZHAO G C,GAO Y H,GAO S L,et al.The phenological growth stages of Sapindus mukorossi according to BBCH scale[J].Forests,2019,10(6):462.DOI: 10.3390/f10060462.
[28]高媛,贾黎明,苏淑钗,等.无患子物候及开花结果特性1)[J].东北林业大学学报,2015,43(6):34-40,123.GAO Y,JIA L M,SU S C,et al.Phenology and blossom-fruiting characteristics of Sapindus mukorossi[J].J Northeast For Univ,2015,43(6):34-40,123.DOI: 10.3969/j.issn.1000-5382.2015.06.007.
[29]李广德,张亚雄,邓坦,等.树干液流及其主要影响因子对摘芽强度的响应[J].农业工程学报,2021,37(5):131-139.LI G D,ZHANG Y X,DENG T,et al.Responses of tree stem sap flow and its main influencing factors to bud pruning[J].Trans Chin Soc Agric Eng,2021,37(5):131-139.DOI: 10.11975/j.issn.1002-6819.2021.05.015.
[30]GRANIER A.Evaluation of transpiration in a Douglas-fir stand by means of sap flow measurements[J].Tree Physiol,1987,3(4):309-320.DOI: 10.1093/treephys/3.4.309.
[31]刘洋,王烨,王斐,等.宽窄行栽植下毛白杨不同方位树干液流的差异[J].中南林业科技大学学报,2018,38(10):95-105.LIU Y,WANG Y,WANG F,et al.Azimuthal variation in sap lfux density of Populus tomentosa under wide and narrow row planting scheme[J].J Cent South Univ For Technol,2018,38(10):95-105.DOI: 10.14067/j.cnki.1673-923x.2018.10.015.
[32]赵飞飞,马煦,邸楠,等.毛白杨茎干不同方位夜间液流变化规律及其主要影响因子[J].植物生态学报,2020,44(8):864-874.ZHAO F F,MA X,DI N,et al.Azimuthal variation in nighttime sap flow and its mainly influence factors of Populus tomentosa[J].Chin J Plant Ecol,2020,44(8):864-874.DOI: 10.17521/cjpe.2020.0089.
[33]CAMPBELL G S,NORMAN J M. An introduction to environmental biophysics[M]. Berlin: Springer Science amp; Business Media,2000.
[34]OISHI A C,HAWTHORNE D A,OREN R.Baseliner:an open-source,interactive tool for processing sap flux data from thermal dissipation probes[J].SoftwareX,2016,5:139-143.DOI: 10.1016/j.softx.2016.07.003.
[35]HUANG L,ZHANG Z S,LI X R.Sap flow of Artemisia ordosica and the influence of environmental factors in a revegetated desert area:Tengger Desert,China[J].Hydrol Process,2010,24(10):1248-1253.DOI: 10.1002/hyp.7584.
[36]JARVIS P G,MONTEITH J L,WEATHERLEY P E. The interpretation of the variations in leaf water potential and stomatal conductance found in canopies in the field[J]. Philos T R Soc B,1976,273(927):593-610. DOI:10.1098/rstb.1976.0035.
[37]VOS J,OYARZN P J.Photosynthesis and stomatal conductance of potato leaves-effects of leaf age,irradiance,and leaf water potential[J].Photosynth Res,1987,11(3):253-264.DOI: 10.1007/BF00055065.
[38]LAVRIC M,ELER K,FERLAN M,et al.Chronological sequence of leaf phenology,xylem and phloem formation and sap flow of Quercus pubescens from abandoned Karst grasslands[J].Front Plant Sci,2017,8:314.DOI: 10.3389/fpls.2017.00314.
[39]TANNER W,BEEVERS H.Transpiration,a prerequisite for long-distance transport of minerals in plants?[J].Proc Natl Acad Sci USA,2001,98(16):9443-9447.DOI: 10.1073/pnas.161279898.
[40]MENZEL C M,SIMPSON D R.Effects of temperature and leaf water stress on panicle and flower development of Litchi (Litchi chinensis Sonn.)[J].J Hortic Sci,1991,66(3):335-344.DOI: 10.1080/00221589.1991.11516161.
[41]马长明,袁玉欣,翟明普.基于物候期的核桃树干液流特征[J].东北林业大学学报,2008,36(1):4-5,9.MA C M,YUAN Y X,ZHAI M P.Sap flow in Juglans regia during different phenophases[J].J Northeast For Univ,2008,36(1):4-5,9.DOI: 10.3969/j.issn.1000-5382.2008.01.002.
[42]HANSEN P.The effect of fruiting upon transpiration rate and stomatal opening in apple leaves[J].Physiol Plant,1971,25(2):181-183.DOI: 10.1111/j.1399-3054.1971.tb01424.x.
[43]郑玉琳,刘济铭,史双龙,等. 无患子果实成熟过程及其油脂、皂苷动态变化[J]. 南京林业大学学报(自然科学版),2021,45(4):76-82. ZHENG Y L,LIU J M,SHI S L,et al. Maturation processes and the dynamics of oil and saponin in Sapindus mukorossi[J]. J Nanjing For Univ (Nat Sci Ed),2021,45(4):76-82. DOI:10.12302/j.issn.1000-2006.202103047.
[44]黄雅茹,李永华,辛智鸣,等.不同时间尺度气象因子与柽柳树干液流关系研究[J].干旱区资源与环境,2020,34(11):149-154.HUANG Y R,LI Y H,XIN Z M,et al.Relationships between meteorological factors and Tamarix chinensis’s sap flow at different time scales[J].J Arid Land Resour Environ,2020,34(11):149-154.DOI: 10.13448/j.cnki.jalre.2020.312.
[45]马文涛,程平,李宏,等.干旱绿洲区富士苹果树干边材茎流动态及其对环境因子的响应[J].浙江大学学报(农业与生命科学版),2020,46(4):428-440.MA W T,CHENG P,LI H,et al.Stemflow dynamics of Fuji apple trunk sap in arid oasis area and its response to environmental factors[J].J Zhejiang Univ (Agric Life Sci),2020,46(4):428-440.DOI: 10.3785/j.issn.1008-9209.2019.10.281.
[46]王力,王艳萍.黄土塬区苹果树干液流特征[J].农业机械学报,2013,44(10):152-158,151.WANG L,WANG Y P.Characteristics of stem sap flow of apple trees in loess tableland[J].Trans Chin Soc Agric Mach,2013,44(10):152-158,151.DOI: 10.6041/j.issn.1000-1298.2013.10.024.
[47]党宏忠,却晓娥,冯金超,等.晋西黄土区苹果树边材液流速率对环境驱动的响应[J].应用生态学报,2019,30(3):823-831.DANG H Z,QUE X E,FENG J C,et al.Response of sap flow rate of apple trees to environmental factors in Loess Platea of western Shanxi Province,China[J].Chin J Appl Ecol,2019,30(3):823-831.DOI: 10.13287/j.1001-9332.201903.015.
[48]麦合木提·图如普,周伟权,丁想,等. 吐鲁番盆地杏树树干液流变化特征及其对环境因子的响应[J]. 生态学杂志,2021,40(8):2378-2387. Mahmoodt·Turup,ZHOU W Q,DING X, et al. Sap flow characteristics of Prunus armeniaca L. and its response to environmental factors in Turpan basin[J]. Chin J Ecol,2021,40(8):2378-2387. DOI:10.13292/j.1000-4890.202108.014.
[49]凡超,邱燕萍,李志强,等.荔枝树干液流速率与气象因子的关系[J].生态学报,2014,34(9):2401-2410.FAN C,QIU Y P,LI Z Q,et al.Relationships between stem sap flow rate of litchi trees and meteorological parameters[J].Acta Ecol Sin,2014,34(9):2401-2410.DOI: 10.5846/stxb201307041839.
(责任编辑 吴祝华)
猜你喜欢
现代农业科技(2017年2期)2017-03-16 20:29:21
安徽农学通报(2017年1期)2017-02-15 17:49:06
热带农业科学(2016年11期)2017-01-21 15:05:16
现代园艺(2016年7期)2017-01-09 22:25:27
农家科技下旬刊(2016年9期)2016-12-15 12:44:18
农业与技术(2016年15期)2016-11-09 07:16:25
科技视界(2016年14期)2016-06-08 18:58:03
中国科技博览(2016年7期)2016-04-25 07:43:53
天津农业科学(2016年3期)2016-03-12 15:57:14
