
发酵饲料对蛋鸡粪便堆肥氨气排放与微生物群落的影响
2024-11-09 00:00:00蒲俊华卢建赵华轩李尚民曲亮王洪志窦新红
农业环境科学学报 2024年8期



摘要:为探究饲喂发酵饲料对蛋鸡粪便堆肥腐熟、氨气排放及微生物群落组成的影响,以饲喂基础日粮蛋鸡为对照组(T1),试验组蛋鸡分别饲喂基础日粮加2.5%(T2)、5.0%(T3)和10.0%(T4)的发酵饲料,收集各组鸡粪进行好氧堆肥试验。结果表明:随堆肥进行,各组堆肥温度均先升高再降低,pH在高温期快速从7.0左右升至9.1。至堆肥结束,T3组和T4组的种子发芽指数分别为99.4%和87.1%,远高于T1组的57.7%,达到有机肥标准。氨气日排放量在高温期达到峰值,随后逐渐降低;氨气总排放量T3组最低,为14.43 mg·kg-1,较T1组低0.76 mg·kg-1。微生物生物信息学分析显示,添加发酵饲料降低了蛋鸡粪便中细菌的多样性,主要的细菌门为厚壁菌门(Firmicutes)、拟杆菌门(Bacteroidota)、放线菌门(Actinobacteriota)和变形菌门(Proteobacteria),其中T2、T3和T4组的乳杆菌属相对丰度较Tl组分别增加20.61%、6.62%和8.13%。随堆肥进行厚壁菌门的丰度降低,其他菌门的丰度增加。研究表明,蛋鸡日粮添加5.0%的发酵饲料可提高蛋鸡粪便中乳杆菌属丰度,促进蛋鸡粪便的堆肥腐熟,并可减少堆肥过程中氨气的总排放量。
关键词:发酵饲料;蛋鸡粪便;堆肥;氨气排放;微生物
中图分类号:S141.4 文献标志码:A 文章编号:1672-2043(2024) 08-1876-12 doi:10.11654/jaes.2023-0918
随着国内畜牧业的快速发展,畜禽养殖量不断增加,产生的大量畜禽粪污造成了较高的环境污染风险。对畜禽粪便进行资源化利用是有效防范和降低畜禽粪便污染环境风险的有效途径。从养殖源头提高饲料利用率,减少未消化吸收的营养物质如氮、磷、钾等的排出,从养殖末端采用堆肥处理畜禽粪便等方式,转化利用未消化吸收的营养物质,有助于实现畜禽养殖业绿色高效循环发展。
近年来随着在饲料中禁用促生长抗生素饲料添加剂,发酵饲料作为抗生素促生长替代物已在畜禽生产中有较多应用。发酵饲料是以植物性农副产品(豆粕、玉米、棉籽粕、菜籽粕和中草药等)为主要原料,采用固态或液态发酵方式,通过有益微生物(乳酸菌、酵母菌、芽孢杆菌和霉菌等)的代谢作用,形成的营养丰富、适口性好及活菌含量高的生物饲料,目前家禽发酵饲料多采用固体发酵方式生产。发酵饲料对畜禽生产的影响主要有3个方面:一是改善饲料品质,降低粗纤维含量,提高粗蛋白含量,增强蛋白酶等酶活性,促进肠道对营养物质的吸收利用,提高畜禽生产性能,如提高蛋鸡产蛋率、改善蛋品质等。二是调节肠道菌群,抑制有害菌群定殖和生长,促进有益菌群增殖。如Guo等发现发酵饲料对蛋鸡盲肠微生物多样性无显著影响,但使厚壁菌门丰度显著降低,乳酸菌、Megasphaera和Peptococcus的相对丰度增加。张孟阳等指出发酵饲料可降低仔鸡肠道菌群丰度和多样性,显著增加乳杆菌的丰度,降低脱硫弧菌属、螺杆菌等致病菌的数量。三是改变畜禽粪便的成分含量、理化性质和微生物群落构成。如饲喂生物发酵饲料的牛粪全氮、全磷和有机质含量显著降低。饲喂发酵饲料的蛋鸡粪便氮、钙等含量降低。Winsen等指出发酵饲料使猪粪pH值显著升高,同时减少猪粪肠杆菌科菌群。Tang等的研究表明发酵全价饲料可显著提高仔猪粪便微生物群的OTU数量和多样性。Zhu等发现饲喂发酵豆粕可显著提高仔猪粪便放线菌、变形杆菌的水平。综上所述,目前发酵饲料对畜禽生产影响的研究多集中在饲料利用方面,鲜有对后续的粪便堆肥处理的研究报道。
畜禽粪便需经过堆肥腐熟,降解其中的植物毒性物质等,才可还田供植物吸收利用。堆肥是利用微生物的代谢活动,将粪便中的有机质降解转化为腐殖质的过程。氮是粪肥中的主要营养物质,与猪、牛等大家畜相比,家禽的消化道较短,粪便中的氮含量较高。氮在腐熟过程中易以氨气的形式向外排放,是造成粪肥氮损失的主要因素,同时也会带来环境污染问题。在堆肥时可通过降低氨气排放,减少氮损失,从而提高有机肥效力并降低环境污染。有研究指出堆肥腐熟的时长与氮损失相关,如人粪便高温堆肥的腐熟期较中温堆肥缩短3-5 d,加快了有机物的腐熟,有效减少了氮损失。另有研究认为粪便堆肥腐熟与日粮来源相关。徐张义等指出与高蛋白日粮相比,低蛋白日粮提高猪粪短时间内好氧堆肥的腐熟度;盛清凯等发现饲喂发酵饲料的猪粪易腐熟。上述研究提示堆肥的腐熟时长与日粮来源和氮损失相关,而日粮来源不同的堆肥腐熟是否与氮损失直接相关并不明确。发酵饲料中的微生物是否会对鸡粪堆肥腐熟产生影响,进而影响氮元素的转化和微生物群落的组成未见相关报道。本研究主要探究不同添加水平的发酵饲料饲喂蛋鸡对粪便堆肥腐熟、氨气排放和微生物群落组成的影响,以期为发酵饲料的开发利用和后续粪污的资源化利用提供理论依据。
1 材料与方法
1.1 试验材料
选取体质量相近的36周龄的海兰褐蛋鸡240只,分为4组(T1、T2、T3、T4),每组60只,设4个重复,每个重复15只。T1组为对照组,饲喂基础日粮,T2、T3、T4组为试验组,分别在基础日粮中添加2.5%、5.05%和10.0%的发酵饲料。基础日粮组成:玉米62.7%、豆粕26.4%、石粉8.6%、预混料2.3%;营养成分:代谢能11.09 MJ·kg-1、粗蛋白16.5%、钙3.5%、总磷1.0%、蛋氨酸0.35%、赖氨酸0.85%。发酵饲料的主要原料:玉米、玉米副产品、小麦麸、豆粕;营养成分:总酸(有机酸总量)≥6%、粗蛋白≥18%、粗灰分≤12%、粗纤维≤8%、水分≤40%;粪肠球菌(Enterococcusfaecalis)≥8x108 CFU.g-1、丁酸梭菌(Clostridiumbutyricum)≥8x 108 CFU·g-1、酿酒酵母菌(Saccharomycescerevisiae)≥2X 108 CFU·g-1、凝结芽孢杆菌(Bacilluscoagulans)≥2x108 CFU·g-1。蛋鸡饲喂8周后,在第9周连续3d收集各组鸡粪进行堆肥试验。
1.2 堆肥试验
称取各组鸡粪8 kg与稻壳按5:1比例混合均匀,碳氮比为24,置于通气发酵罐中,发酵罐通气量为0.4 L·min-1·kg-1,每日上午10时记录堆肥温度,并用2%硼酸溶液20 mL采集氨气(NH3)1 h。在堆肥第0、1、3、5、10、14、21、28天进行翻堆后采集堆肥样品,样品一部分保存于-20℃备测pH、电导率(EC)、总氮(TN)、硝态氮(NO-3-N)、铵态氮(NH+4-N)、种子发芽指数(GI),一部分保存于-80℃待提取微生物总DNA。
1.3 测定指标及方法
理化指标的测定。氨气:盐酸滴定法;pH、EC:将样品与超纯水按1:10的质量体积比混合均匀,中速振荡2h,取上清液,分别用pH计(PHS-3C,上海雷磁)和电导率测定仪(DDS-307A,上海雷磁)测定pH值和EC值;总氮、硝态氮、铵态氮:参考《有机肥料》(NYIT 525-2021)和《肥料硝态氮、铵态氮、酰胺态氮含量的测定》(NY/T 1116-2014)。
GI的测定。采用萝卜种子,参考NY/T 525-2021标准测定,具体操作:取4.5 g样品,加入45 mL超纯水混合均匀,中速振荡th,静置30 min,移取上清液10 mL于培养皿中,培养皿内放1张定性滤纸,在滤纸上均匀放置10颗萝卜种子,盖上培养皿盖,25℃避光培养48h,统计发芽的种子数,并测量主根长,同时用超纯水做空白对照。GI=样品发芽率X样品平均根长/(空白发芽率X空白平均根长)%100%。
微生物群落组成的测定。采用16S rRNA基因测序分析,具体操作:提取样品微生物的总DNA(试剂盒QIAamp Fast DNA Stool Mini Kit),对16S rRNA基因的V3-V4区用引物338F 5’-ACTCCTACGGGAG-CCAGCAG -3'和806R 5'- GGACTACHVGCGWTC-TAAT-3'进行扩增,采用Illumina MiSeq平台对PCR产物测序,利用美吉生物信息分析云平台进行微生物生物学信息分析。
1.4 数据处理
采用Excel 2019进行数据整理,采用GraphPadPrism 9.0软件分析绘图。
2 结果与分析
2.1 堆肥温度、pH值和EC值的变化
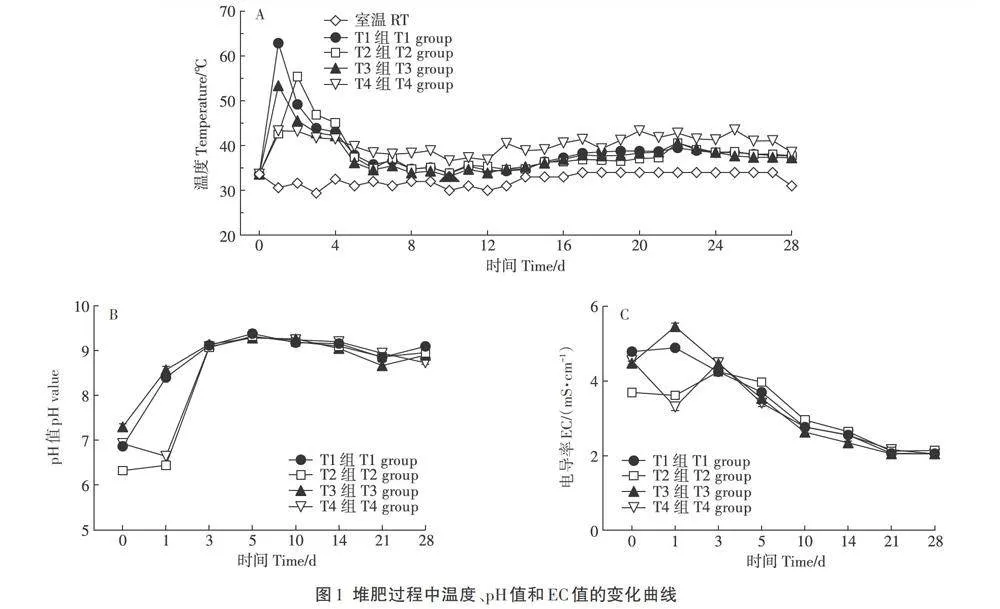
温度是堆肥的一个重要参数,温度的变化显示着堆肥的进程。由图1A可见,对照组和试验组在堆肥过程中的发酵温度先升高后降低,依据堆肥温度的变化可将堆肥过程分为升温期(0-2 d)、高温期(3-5d)、降温腐熟期(6-28 d)。其中T1和T3组升温较快,经过24 h堆肥,温度即上升至62.9℃和53.4℃,而T2和T4组经过48 h堆肥温度上升至55.4℃和43.2℃。在腐熟期T1、T2和T3组温度降至40℃以下,T4组温度在40℃左右。
由图IB可见,堆肥初始期,各组的pH值范围为6.3-7.3,在升温期,T1和T3组的pH值迅速升高,T2组的pH值先缓慢升高而后再迅速升高,T4组的pH值先降低而后再迅速升高,至第3天各组pH值接近9.1,随后变化趋势相似,末期在小范围波动。整个堆肥过程中pH值的变化符合堆肥发酵的一般规律,腐熟的有机肥一般呈碱性。
EC反映堆肥有机离子和无机离子浓度的大小。图1C显示,堆肥初始期T1组的EC值为4.8 mS·cm-1,高于试验组的EC值,试验组EC值高低依次为T4>T3>T2组。堆肥期间T1与T3组的EC值先升高后逐渐降低,T2与T4组的EC值先降低后增加再逐渐下降。至堆肥期结束,各组的EC值均降至约2.0 mS·cm-1。
2.2 氨气排放量的变化
随着堆肥的进行,对照组和试验组氨气日排放量均表现为先升高再降低的变化过程(图2A)。在堆肥升温期,T2、T3和T4组氨气日排放量均低于T1组,在堆肥第4-5天各组氨气日排放量达到高峰,其中T3组最高,为1.81 mg·kg-1,其余T2、T4和T1组氨气日排放量均高于1.48 mg·kg-1。从降温期第6天开始各组氨气日排放量均呈下降趋势,至堆肥期结束(第28天)已低于0.13 mg·kg-1。图2B显示,各组氨气的周排放量呈依次降低趋势,氨气排放主要集中在第1周和第2周,第1周氨气排放量T2、T3和T4组均低于T1组,各组两周累计的排放量均超过总排放量的75%。由图2C可见,在整个堆肥期内,各组氨气总排放量呈上升趋势。在堆肥初期至堆肥第14天,氨气总排放量快速增加,而在堆肥后半段(14-28 d),氨气总排放量增加缓慢,至堆肥期结束各组氨气总排放量由高到低的顺序依次为T2组(15.29 mg·kg-1)>T1组(15.19 mg·kg-1)>T4组(15.05mg·kg-1)>T3组(14.43 mg·kg-1)。
2.3 总氮、铵态氮、硝态氮的含量变化
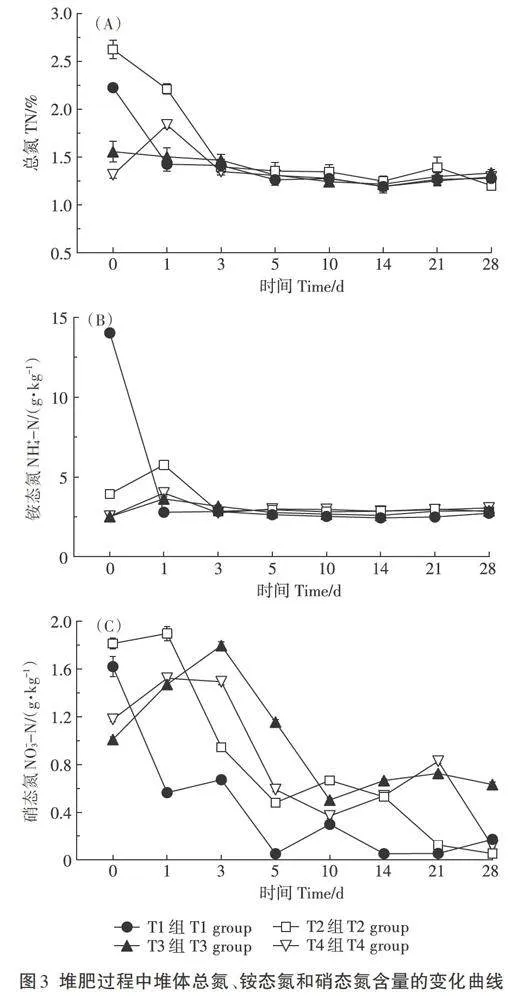
各组的总氮、铵态氮和硝态氮变化趋势见图3。在堆肥初始期,总氮含量为T2组>TI组>T3组>T4组,而后T4组总氮含量先升高(1 d)再降低,T1和T2组先快速降低(3 d)而后再缓慢降低,T3组一直呈缓慢降低趋势(图3A)。初始期铵态氮含量以T1组(1.40%)最高,远高于T2、T3和T4组。T1组第1天铵态氮含量快速降低,试验组铵态氮含量均先增加再降低(图3B)。综合图3A、图3B可见,高温期后各组总氮和铵态氮含量趋于一致,并保持至堆肥结束。硝态氮含量在堆肥初始期表现为T2组>T1组>T4组>T3组,在堆肥升温期T2、T3和T4组硝态氮含量上升,而T1组硝态氮含量急剧下降,在高温期,T3和T4组硝态氮含量高于T2和T1组,在降温期各组硝态氮含量呈波动性降低趋势(图3C)。
2.4 GI的变化
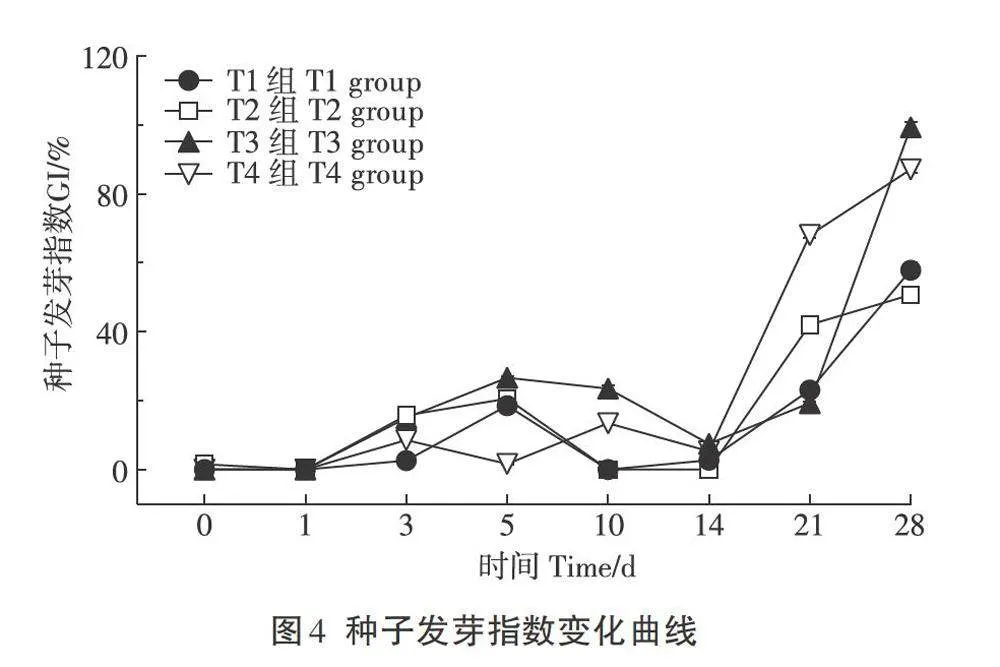
GI常用来反映有机肥的腐熟度,只有达到腐熟标准的有机肥才能施用于田间,供植物利用。一般来说,当CI<50%时认为堆肥未腐熟;农业行业标准《有机肥料》(NY/T 525-2021)中要求GI≥70%。由图4可见在堆肥初始升温期,各组的GI接近0,随堆肥的进行,各组的GI呈增高趋势,在堆肥的第21天,各组CI出现较大差异,其中T4组GI接近70≥,高于其余3组。至堆肥期结束,T3和T4组GI分别为99.4%和87.1%,高于T1(57.7%)和T2(50.6%)组,已达到有机肥标准。
2.5 微生物生物信息分析
2.5.1 微生物细菌分类统计及α多样性分析
所有样品经测序获得的数据在各分类水平上的结果为22个门、41个纲、113个目、213个科、402个属、604个种、1 004个操作分类单元(OTU)。图5显示在OTU水平上4个组共有631个共同OTU,各组特有的OTU数量分别是T1组24个、T2组8个、T3组17个、T4组7个。
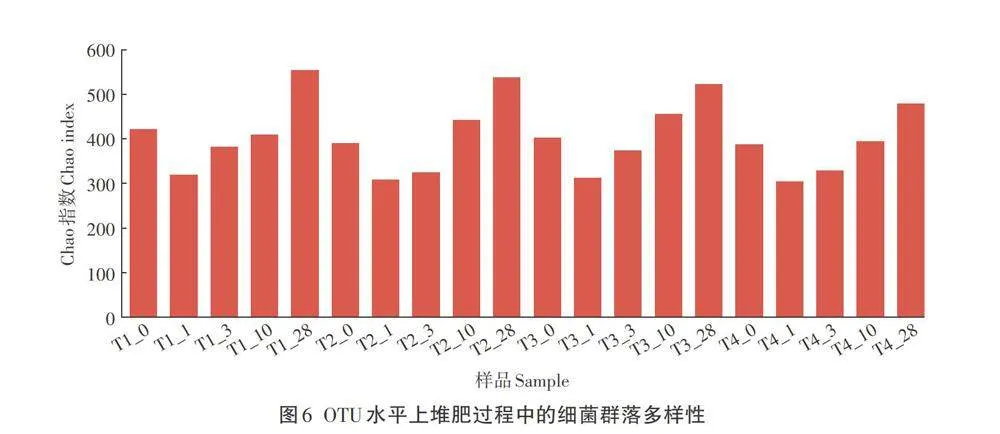
Chao指数常用来估计物种的总数,其数值越大,物种的数量越多。由OTU水平上Chao指数图(图6)可见,在堆肥初始期,T1组的Chao指数高于T2、T3和T4组,在堆肥过程中各组的Chao指数均先下降再逐渐升高,表明菌群的物种数量与堆肥进程相关。到堆肥结束,各组的Chao指数大小依次为T1组>T2组>T3组>T4组。
2.5.2 细菌群落结构分析
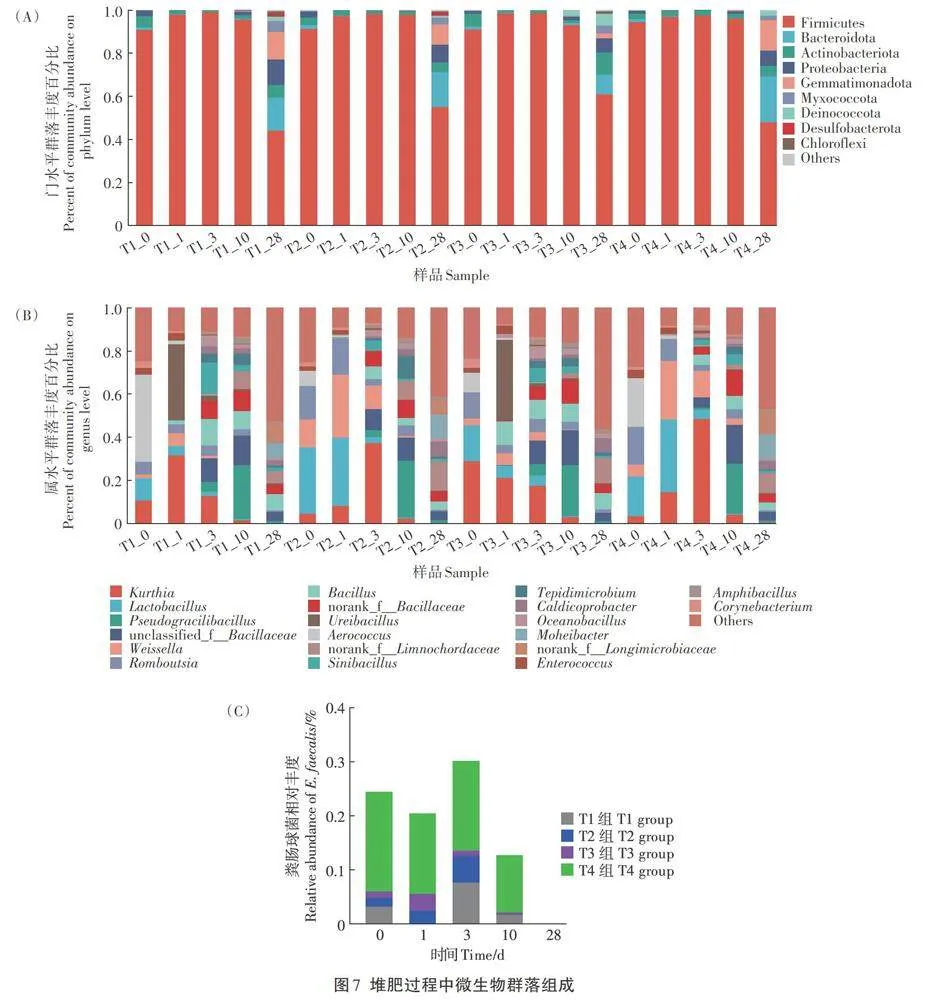
如图7所示,经过28 d的堆肥发酵,对照组和试验组的细菌群落结构在门和属水平的组成均发生了改变。图7A显示,门水平上厚壁菌门(Firmicutes)、拟杆菌门(Bacteroidota)、放线菌门(Actinobacteriota)和变形菌门(Proteobacteria)为堆肥初始期的主要菌门,其中厚壁菌门占绝对优势。随堆肥的进行,厚壁菌门丰度降低,至堆肥结束其在T1、T2、T3、T4组的丰度分别为44.12%、55.07%、61.01%、48.03%,而拟杆菌门、放线菌门和变形菌门的丰度升高。此外,芽单胞菌门(Gemmatimonadota)和黏球菌门(Myxococcota)的丰度也增加,上述6种菌门在T1、T2、T3、T4组的总丰度分别达到94.84%、96.59%、92.89%、97.47%。
图7B显示,属水平上乳杆菌属(LactobaciZlus)、罗姆布茨菌属(Romboutsia)和魏斯氏菌属(Weissella)是试验组堆肥初始期的优势菌,如乳杆菌属在T2、T3和T4组的丰度分别为30.79%、16.80%和18.31%,高于T1组(10.18%)。气球菌属(Aerococcus)是T1组的优势菌属,其丰度为40.30%,高于T2组(7.10%)、T3组(9.07%)和T4组(22.61%)。随堆肥进行,各组的优势菌属出现变化,如乳杆菌属和气球菌属在升温期至高温期的丰度均快速降低。尿素芽孢杆菌属(Ureibacil-lus)的丰度在T1和T3组第1天急剧上升为35.16%和37.21%,远高于T2和T4组。一些菌属在堆肥高温期后出现,如norank_f_Bacillaceae、芽孢杆菌属(Bacil-lus)、八叠球菌属(Tepidimicrobium)、假纤细芽孢杆菌属(Pseudogracilibacillus)、unclassified_f_Bacillaceae、大洋芽孢杆菌属(Oceanobacillus)、双芽孢杆菌属(Am-phibacillus)等,均呈现先增加后降低规律。还有一些菌如漠河杆菌属(Moheibacter)、嗜热杆菌属(Caldi-coprobacter)、norank-f-Longimicrobiaceae菌属等在堆肥中后期出现。肠球菌属(Enterococcus)丰度相对较低,随堆肥进行丰度下降。堆肥结束各组低比例菌群合并后的占比上升,T1、T2、T3、T4组分别达到5 2.65%、41.23%、56.33%、47.31%。
图7C为发酵饲料中的粪肠球菌(E.faecalis)在堆肥中的变化情况。在堆肥初始期,各组粪肠球菌相对丰度高低依次为T4组>T1组>T2组>T3组。在升温期T1组丰度急剧降低,T2和T3组丰度增加,T4组丰度略降,在高温期除T3组丰度降低外,其余组丰度增加,而在降温期各组丰度均降低,在堆肥末期未检测到粪肠球菌。除粪肠球菌外,其余两种细菌丁酸梭菌(Clostridium butyricum)和凝结芽孢杆菌(Bacillus co-agulans)在堆肥初始期未检测到,说明其在蛋鸡肠道内与其他微生物产生竞争,未成为鸡粪中的优势菌。
2.5.3 环境因子与微生物群落的关系分析
从图8环境因子与微生物群落(门水平)的相关性关系可见,GI、总氮、硝态氮、EC、氨气与多种微生物的关系较密切,而pH和铵态氮与单一的微生物的关系较密切。厚壁菌门和互养菌门(Synergistota)与GI呈显著负相关(P≤0.05),而厚壁菌门与硝态氮(0.01<P≤0.05)、EC(0.001<P≤0.01)和氨气(P≤0.001)呈显著正相关,互养菌门与总氮和EC呈显著正相关(0.01<P≤0.05)。Patescibacteria是唯一与pH呈显著负相关的菌门。奇球菌门(Deinococcota)、变形菌门、芽单胞菌门、黏球菌门与总氮、硝态氮、氨气和EC呈显著负相关(P≤0.001),而与GI呈正相关(0.01<P≤0.05或0.001<P≤0.01)。脱硫杆菌门(Desulfobacte-rota)是唯一与铵态氮呈显著负相关的菌门(0.001<P≤0.01)。拟杆菌门、Sumerlaeota、绿弯菌门(Chloroflexi)与总氮、铵态氮、硝态氮和EC呈显著负相关(0.01<P≤0.05或0.001<P≤0.01或P≤0.001),放线菌门、unclassi-fied-k-norank-d-Bacteria与EC和氨气呈显著负相关(0.001<P≤0.01或P≤0.001)。隐杆菌门(Abditibacteri-ota)、浮霉菌门(Planctomycetota)、蛭弧菌门(Bdellovi-brionota)与硝态氮呈显著负相关(0.01<P≤0.05),与EC和氨气呈极显著负相关(0.001<P≤0.01或P≤0.001)。盐厌氧菌门(Halanaerobiaeota)、纤维杆菌门(Fibrobac-terota)与EC呈显著负相关(0.01<P≤0.05),蓝藻门(Cyanobacteria)与硝态氮呈显著负相关(0.01<P≤0.05)。
2.5.4 微生物细菌群落功能预测分析
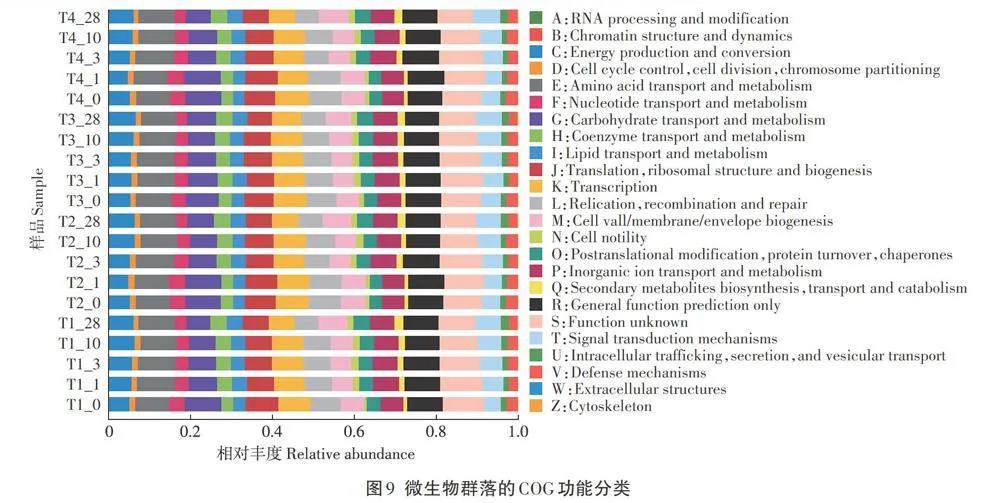
利用PICRUSt软件对堆肥过程中微生物群落进行COG功能分类预测,结果如图9所示。微生物群落COG功能主要集中在能量生产和转化、物质(氨基酸、碳水化合物、核苷酸、辅酶、脂质、无机离子等)的运输和代谢、信号转导机制、遗传物质的翻译修饰等方面。其中氨基酸和碳水化合物的转运与代谢是微生物功能集中度高的代谢途径。氨基酸的代谢途径在堆肥高温期第3天达到峰值,T1、T2、T3、T4组的丰度分别为9.22%、9.41%、9.22%和9.45%,随后在堆肥降温期略有降低。碳水化合物代谢途径在T1-T4组的初始丰度分别为8.94%、8.75%、7.94%和8.83%,且随堆肥进行逐渐降低,至堆肥末期各组分别减少了3 .07%、2.27%、0.82%、2.72%,T3组减少的比例最低。堆肥发酵的实质是围绕氮、碳元素的转化进行,随堆肥发酵期的变化,相应的微生物功能集中的丰度也发生变化。堆肥初始期信号转导机制在对照组的丰度为4.07%,在试验组(T2、T3、T4组)的丰度分别为4.54%、4.83%、4.69%,均高于对照组,随堆肥的进行,各组的信号转导机制进一步富集,试验组随发酵饲料添加量的增加,信号转导机制丰度增加,至堆肥末期T4组丰度达6.74%。此外,能量的产生和转化途径在堆肥第1天下降,而从第3天起至堆肥结束呈持续增加状态,表明能量的产生和转化伴随堆肥的整个过程。
3 讨论
3.1 发酵饲料对鸡粪堆肥腐熟度的影响
发酵饲料是一类含有活菌的生物饲料,在畜禽生产上已有较多应用。家禽粪便中含有较多的氮、钾等物质,将其进行堆肥腐熟转化为有机肥后可供植物吸收利用。未腐熟的堆肥具有植物危害性和环境风险性,与堆肥腐熟度相关的指标包括pH、EC和GI等。本试验中各组的鸡粪初始pH值在7.0左右,经过堆肥升温期,pH快速升高至9.1,呈碱性,表明鸡粪堆体中的有机氮转为了铵态氮。此外,本试验中pH值的快速提高与堆肥升温速率同步,这与姜新有等的研究得到的pH值的变化与堆肥温度有关的结论相似。EC值反映腐熟粪肥对植物生长抑制或生物毒性作用的程度,有研究表明粪肥EC高于4mS·cm-1不利于植物生长,而本研究中各组EC值均低于2.1 mS·cm-1,表明其对植物生长无抑制作用。GI直接反映堆肥腐熟度,本试验中各组CI均高于50%,且5.0%和10.0%发酵饲料组的鸡粪经过28 d堆肥腐熟,GI均远高于70%,达到有机肥标准,说明添加5.0%或10.0%的发酵饲料饲喂蛋鸡可以促进鸡粪堆肥腐熟。盛凯清等报道饲喂发酵饲料的猪粪易腐熟,其堆肥过程中纤维素酶、中性蛋白酶、脲酶等活性变化与猪粪添加外源菌剂堆肥发酵的酶活性变化类似,说明发酵饲料影响猪粪有机肥的生产。本试验中饲喂发酵饲料促进鸡粪堆肥GI升高的机制是否与堆肥时添加外源菌剂促进GI升高的机制相似还有待于进一步研究,如陈倩倩等报道在羊粪与鸡粪混合堆肥中,添加嗜热复合菌使堆肥的GI升高,促进堆肥腐熟。
3.2 发酵饲料对鸡粪堆肥过程中氨气排放的影响
氮元素是有机肥的主要营养元素之一,畜禽粪便中氮元素主要以有机氮的形式存在。在堆肥过程中氮的转化较为复杂,主要通过矿化、挥发、硝化、固定化和反硝化作用进行转化。与家畜粪便相比,鸡粪中的氮含量较高,在堆肥过程中总氮的损失高达31.8%,牛羊粪总氮损失约为22.5%,猪粪便总氮损失为28.6%。堆肥氮损失主要来自氨气、一氧化二氮、氮气和NOx化合物的气体排放,其中氨气排放占主导地位,可占到总氮损失的68.2%。氮损失不仅带来肥料价值的损失,同时其难闻的气味还带来环境和健康问题。影响氨气产生的因素包括初始铵态氮浓度、温度等。堆肥初始期的低铵态氮浓度使初始投入的铵态氮减少,限制氨气的产生,而在堆肥过程中,氨气的排放集中在堆肥高温期,降低高温期氨气的挥发是提高粪肥中氮含量的关键因素。本试验中氨气主要在堆肥升温期和高温期产生,试验组在此阶段的氨气总排放量低于对照组,这可能与堆肥初始期试验组的铵态氮含量均低于对照组有关。本试验的结果表明蛋鸡饲喂含发酵饲料的日粮,可抑制鸡粪在堆肥前期和中期氨气的挥发,降低氮损失,有助于堆肥氮元素的保留,这与刘闰等报道的蛋鸡饲喂菌酶协同发酵饲料可显著降低蛋鸡粪便氨气产生的结果有相似之处。
3.3 发酵饲料对鸡粪堆肥微生物群落结构的影响
发酵饲料作为含有活菌的生物饲料,不仅改善动物肠道对营养物质的吸收利用,同时也影响动物肠道微生物群落构成。畜禽粪便中的微生物主要源自肠道微生物,随堆肥进行,微生物群落组成也发生相应的变化。本试验中饲喂发酵饲料的蛋鸡粪便在堆肥过程中,其细菌群落多样性均低于对照组,表明发酵饲料改变蛋鸡粪便的微生物群落构成,且对微生物群落多样性的影响贯穿堆肥过程。畜禽粪便堆肥腐熟的实质是粪便中微生物分解利用有机质的结果,粪便中微生物群落的变化对粪肥的堆肥转化有直接的影响作用。本试验中厚壁菌门、拟杆菌门、放线菌门和变形菌门是蛋鸡粪便的主要菌门,且随堆肥进行厚壁菌门的丰度下降,而其余3种菌门的丰度则增加,这与前人的一些研究结果相似,如于静等冈、王秀红等报道指出鸡粪好氧堆肥中厚壁菌门、拟杆菌门、放线菌门、变形菌门是主要的菌门。堆肥过程中细菌的丰度变化与其结构特征相关,厚壁菌门的细菌能够产生耐热芽孢,因而在堆肥升温期和高温期的丰度高,且厚壁菌门的细菌对纤维素、多糖等有机物的降解有促进作用,与氮的转化和氨气的挥发关系密切。陶兴玲等研究发现厚壁菌门对有机质降解的影响最强。Wang等在蛋鸡粪好氧堆肥中添加嗜热芽孢杆菌改变了堆肥升温和高温期的细菌群落结构,增加了乳酸菌和硝化菌的相对丰度,缩短高温期,降低pH值,促进了硝化作用,降低了氨气的挥发。本试验中饲喂发酵饲料的蛋鸡粪便中厚壁菌门的乳杆菌属和魏斯氏菌属的丰度在堆肥初始期均高于对照组,这可能与发酵饲料中添加的丁酸梭菌和凝结芽孢杆菌有关。凝结芽孢杆菌属兼性厌氧菌,在蛋鸡肠道中消耗游离氧,利于厌氧微生物乳酸菌的生长,而丁酸梭菌为厌氧菌,在肠道内可抑制有害菌如大肠杆菌的生长等,促进有益菌如乳酸菌、双歧杆菌等的增殖,促进纤维素等有机物降解和堆肥腐熟。本试验饲喂的发酵饲料中的粪肠球菌在蛋鸡肠道内体现出量的优势,且在蛋鸡粪便中以10.0%添加量的相对丰度最高,在堆肥过程中保持优势至堆肥后期。堆肥过程中主要菌群的丰度变化也反映了堆肥反应的连续性和持续性。
参与堆肥的微生物除细菌外,还包括真菌和放线菌,其中鸡粪堆肥以细菌占主导地位,基于本试验主要研究细菌的组成变化情况,未探究真菌和放线菌的组成变化,故未对发酵饲料中包含的酿酒酵母真菌的变化情况做分析。
3.4 发酵饲料对环境因子与微生物群落的关系影响
堆肥是通过微生物的代谢作用转化粪肥中的营养物质,其活动受到堆肥环境因子(温度、pH、氨气浓度等)的影响,微生物的优势菌群发生相应改变,而优势菌群的代谢活动同时也对环境因子产生影响。本试验中厚壁菌门在堆肥升温期和高温期均占绝对优势,代谢活跃,分解有机物,产生大量的氨气,因而与氨气的产量呈极显著正相关,而随着降温期的延长厚壁菌门丰度下降,氨气的产量降低,大部分的有机质已降解,对植物的毒性也降低,因而厚壁菌门与GI值呈显著负相关。于静等研究认为温度和pH是堆肥过程中微生物群落结构改变的主要驱动因子。本试验中pH仅与Patescibacteria菌门呈显著负相关,而硝态氮、氨气产量、EC与多数菌门呈显著负相关,GI则与多数菌门呈显著正相关,表明这些环境因子与堆肥中细菌菌门的关系更密切。
3.5 发酵饲料对微生物细菌功能预测的影响
16S rRNA基因序列测序分析得到微生物细菌物种组成,通过PICRUSt软件结合已知微生物基因组信息,预测微生物的功能,与宏基因组测序结果相比,其细菌功能预测准确性较高,目前PICRUSt较多应用于微生物细菌和古菌的功能预测研究。本研究中PICRUSt软件预测的细菌功能主要集中在物质的代谢转化上,其中氨基酸和碳水化合物的转运与代谢丰度相对较高,在高温期氨基酸的代谢丰度达到顶点,这与堆肥的进程相吻合,高温期氮代谢活跃,有机氮在微生物的氨化作用下转化为无机态的铵态氮。崔鹏等利用PICRUSt2软件预测超高温和常温堆肥微生物群落的功能,发现两种堆肥方式的微生物功能代谢通路都包括氨基酸类物质代谢、碳水化合物代谢、能量代谢和遗传信息复制、翻译等。本试验中还发现发酵饲料促进信号转导机制的富集。
4 结论
(1)综合本试验结果,在蛋鸡日粮中添加5.0%和10.0%的发酵饲料,有利于蛋鸡粪便的堆肥腐熟,经过28 d堆肥,种子发芽指数分别为99.4%和87.1%,达到有机肥堆肥标准。添加5.0%发酵饲料时蛋鸡粪便堆肥的氨气总排放量最低,为14.43 mg·kg-1。
(2)添加发酵饲料降低蛋鸡粪便的细菌多样性,厚壁菌门、拟杆菌门、放线菌门和变形菌门是蛋鸡粪便的主要菌门,随堆肥进行,厚壁菌门的丰度降低,其余菌门的丰度升高。添加发酵饲料提高蛋鸡粪便中乳杆菌属的相对丰度。堆肥环境因子硝态氮、氨气、电导率与厚壁菌门呈显著负相关,而与拟杆菌门、变微生物信号转导机制的富集。
(责任编辑:李丹)
基金项目:江苏省重点研发(现代农业)计划项目(BE2022379);江苏现代农业产业技术体系建设专项资金项目(JATS[2023]069);常州市科技项目(CE20222025);山东省科技型中小企业创新能力提升工程项目(2022TSGC1224);潍坊市科技型中小企业创新能力提升工程计划项目(2022TS1122)
猜你喜欢
湖南饲料(2022年6期)2022-12-20 07:10:16
当代水产(2021年2期)2021-03-29 02:57:16
吉林农业(2016年12期)2017-01-06 19:17:43
广东饲料(2016年4期)2016-12-01 03:43:18
广东饲料(2016年1期)2016-12-01 03:43:02
中小企业管理与科技·下旬刊(2016年11期)2016-11-18 08:19:35
天津农业科学(2016年9期)2016-11-17 11:15:56
中国科技博览(2016年19期)2016-10-19 11:47:05
科技视界(2016年9期)2016-04-26 12:23:48
科技视界(2016年7期)2016-04-01 09:39:11
