
相同DLI下不同光照周期与强度对半结球生菜及上海青生长形态和营养品质的影响
2024-11-07 00:00:00何朋蔚阳树英卢安琪周嘉毅江怀军王汝旭陈勇良廖绪胤
江苏农业科学 2024年18期







摘要:以半结球生菜(Lactuca sativa L.)和上海青(Brassica chinensis L.)为试验材料,使用LED作为光源,在相同日累积光量(daily light integral,DLI)下,探究不同光照周期(8、12、16、20 h/d)与光照度[378、252、189、151 μmol/(m2·s)]组合对作物生长形态及营养品质的影响。结果表明:相同DLI下,合理延长光照周期、降低光照度可提升半结球生菜及上海青光合能力及电能利用效率,促进根系生长,缩短种植周期,增加生物量积累,并提升营养品质,但易增加作物硝酸盐、亚硝酸盐含量的积累。不同作物对光照组合变化的响应具有一定的差异性。本试验条件下,16 h/d、189 μmol/(m2·s)的光照组合利于增加半结球生菜及上海青产量,同时提升半结球生菜营养品质,而12 h/d、252 μmol/(m2·s)的光照组合更利于提高上海青营养品质。光照时长由8 h/d延长至16 h/d,半结球生菜的叶片数、株高、根长、单株干重、可溶性蛋白、游离氨基酸、可溶性糖、纤维素及维生素C含量分别增加20.3%、9.7%、148.1%、127.3%、41.5%、55.2%、83.2%、34.3%和3.4%,上海青分别增加26.0%、18.2%、117.3%、139.0%、21.3%、77.8%、74.2%、23.7%和3.5%。
关键词:半结球生菜;上海青;日累积光量;光照周期;光照度;生长形态;营养品质
中图分类号:S636.204;S634.304 文献标志码:A
文章编号:1002-1302(2024)18-0160-09
收稿日期:2023-10-30
基金项目:国家重点研发计划子课题(编号:2017YFD200803-01);国家自然科学基金面上项目(编号:31471446);中央引导地方科技发展基金(编号:2023ZYQ048)。
作者简介:何朋蔚(1999—),男,湖南长沙人,硕士,主要从事农业生态学方面的研究。E-mail:happyway1124@163.com。
通信作者:阳树英,博士,教授,从事农业生态学方面研究。E-mail:ysyalxh@126.com。
设施农业是在环境相对可控条件下,采用工程技术手段,进行高效生产的一种现代农业模式。相较于传统温室与露地栽培,设施农业可通过补光设备对作物进行光调控,增加作物产量,缩短种植周期以及提高作物品质,但在设施农业中使用LED灯进行补光,架设数量多,一次性投入成本高,加之光照调控技术的缺乏,极大限制了生产效率及经济收益[1-2]。
大量有关调节光强度与光周期来提高蔬菜生产效率的研究指出,光照度一定时,随着光照周期的延长,瓜类的产量和品质有效提高[3-6],生菜产量及叶绿素含量显著增加[7-9],大蒜的营养品质指标先升高后下降[10]。光照周期一定时,随着光照度的削弱,植物将具有荫蔽适应特征。例如株高和叶长的增加,光强度过高会降低植株叶片气孔开度,使水分及CO2的运输受到阻碍,减缓光合速率,抑制植株生长[11-14]。
尽管相关研究内容丰富,但大多仍驻足于FYFxkD+NmYwto0giMyfhBg==改变光照总量的维度,对于相同光照总量下,不同光照周期与强度对作物影响的探究则鲜有涉足。日累积光亮(daily light integral,DLI)指植物一天内光照时长与光照度的乘积,可为研究植物对光的响应机制提供较为全面的参数,常被用来代替光合光量子通量密度(photosynthetic photon flux density,PPFD)作为影响植物生长发育的光变量[15-16]。有研究表明,DLI不变时,改变光照周期与光照度组合可为作物的生长发育带来不同影响,在现行能耗水平上,可进一步通过光照时间与光照度间的反向调控来优化设施农业的生产效率和作物品质[7,17]。
半结球生菜(Lactuca sativa L.)为菊科莴苣属一年生或二年草本植物,上海青(Brassica chinensis L.)为十字花科芸薹属一年生或二年生草本植物。这2种蔬菜光照适应度广,种植周期短,营养丰富,在设施农业生产中大量种植,对二者进行光调控研究不仅具有较大的应用价值,还可比较不同作物对光环境响应机制的异同。因此,本试验立足低碳高效的农业发展趋势,以半结球生菜和上海青为例,探究日累计光亮相同时,不同光照周期及强度组合对作物生长发育的影响,为我国设施农业生产提供参考。
1 材料与方法
1.1 试验地点与材料
盆栽试验于2022年3—5月在湖南农业大学环境工程实训基地人工控制气候室进行。指标测定在洞庭湖区农村生态系统健康湖南省重点实验室进行。供试生菜种子由南京理想农业科技有限公司提供,品种名为理想401半结球型生菜;供试上海青种子由武汉市九头鸟种苗有限公司提供,品种名为金品上海青。试验光源为深圳市绿景光电有限公司生产的LED灯管,灯珠比为红光R(620~660 nm)∶蓝光B(440~470 nm)∶白光W(全光谱)=3∶2∶1,额定功率18 W。
1.2 试验设计
本试验于3月初开始育苗,3月30日开始进行处理,5月2日结束。种子经过24 h温水浸泡催芽后,每孔1至2颗播种于装有基质(V草炭∶V珍珠岩∶V蛭石=3∶1∶1)的50孔育苗盘中。试验开始时,选取具有3张真叶且长势一致菜苗移栽于直径0.15 m,深度0.2 m的土基盆栽中,每盆1株,每隔7 d浇灌0.2%浓度的磷酸二氢钾及尿素水溶液。土壤基质为红壤,取自湖南农业大学环境与生态学院实训基地(28°18′40″N,113°09′12″E),理化性质:pH值6.7,有机质含量19.22 g/kg,总氮含量1.13 g/kg,总磷含量1.36 g/kg,总钾含量14.5 g/kg,速效钾含量53.7 mg/kg。
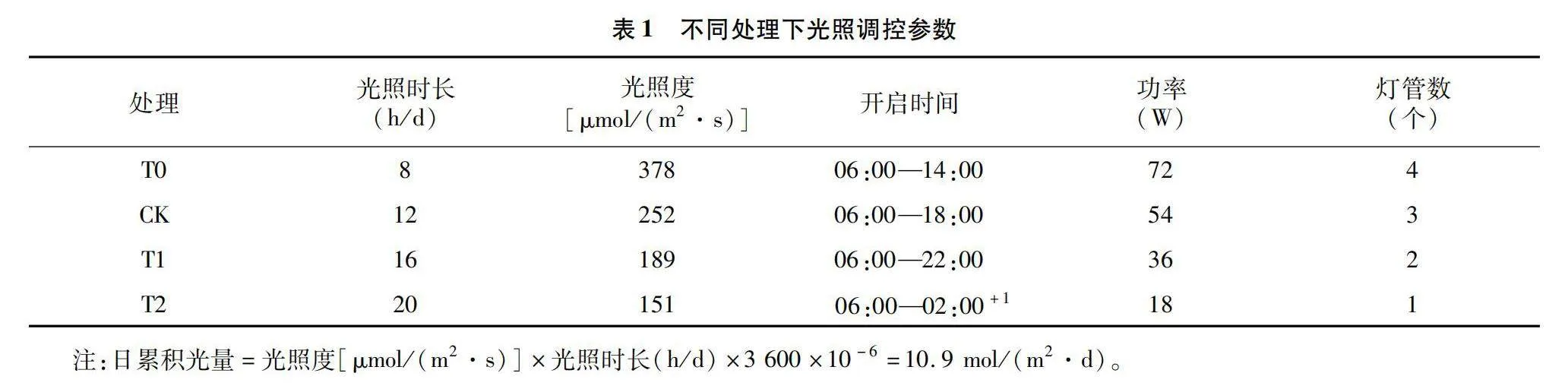
在相同DLI下设置不同光照周期与强度处理:T0[8 h/d、378 μmol/(m2·s)]、T1[16 h/d、189 μmol/(m2·s)]及T2[20 h/d、151 μmol/(m2·s)],以现行补光参数为CK[12 h/d、252 μmol/(m2·s)],各处理具体参数见表1。
1.3 测定指标及方法
1.3.1 生长形态指标的测定
分别于试验处理当天、第8天、第16天、第24天和第32天,于各处理组选择6株进行形态测定。从基质表面向上5 mm为起始点,记录到作物生长最高点的距离为株高,统计每株叶片数,并使用叶面积仪CI-202(美国CID公司)扫描最大叶面积。第32天将植株地上部分和地下部分分开,分别测量鲜重,使用根系扫描仪EPSON 1680(SEC Inc,JPN)测定根系指标,烘干后测定地上部和根系干重,并计算根冠比及含水率。
1.3.2 生理指标及营养品质的测定
维生素C含量的测定采用二甲苯萃取比色法[18];叶绿素含量测定采用95%乙醇比色法[18];游离氨基酸含量的测定采用茚三酮溶液显色法[18];可溶性蛋白含量的测定采用考马斯亮蓝G-250染色法[18];可溶性糖含量的测定采用蒽酮比色法[18];丙二醛(MDA)含量的测定采用硫代巴比妥酸(TBA)显色法[18];硝酸盐及亚硝酸盐含量的测定采用苯酚分光光度计法[19]。
1.3.3 能源效率计算
结合作物产量、补光成本等信息计算得出作物电能利用效率,公式如下[3]。
电能利用效率=单株干重×Wche×种植密度×种植面积光源功率×总光照时长。
式中:Wche为1 g干重代表的化学能,取值为2×104 J/g[20]。种植面积和密度在本研究中分别为1.08 m2和25.9株/m2。
1.4 数据处理
运用Microsoft Excel 2010软件对数据进行计算和处理,利用Origin 2022软件进行数据整理及图表绘制,使用IBM SPSS Statistics软件进行主成分分析和方差分析。
2 结果与分析
2.1 不同光照周期与强度对半结球生菜及上海青生长形态的影响
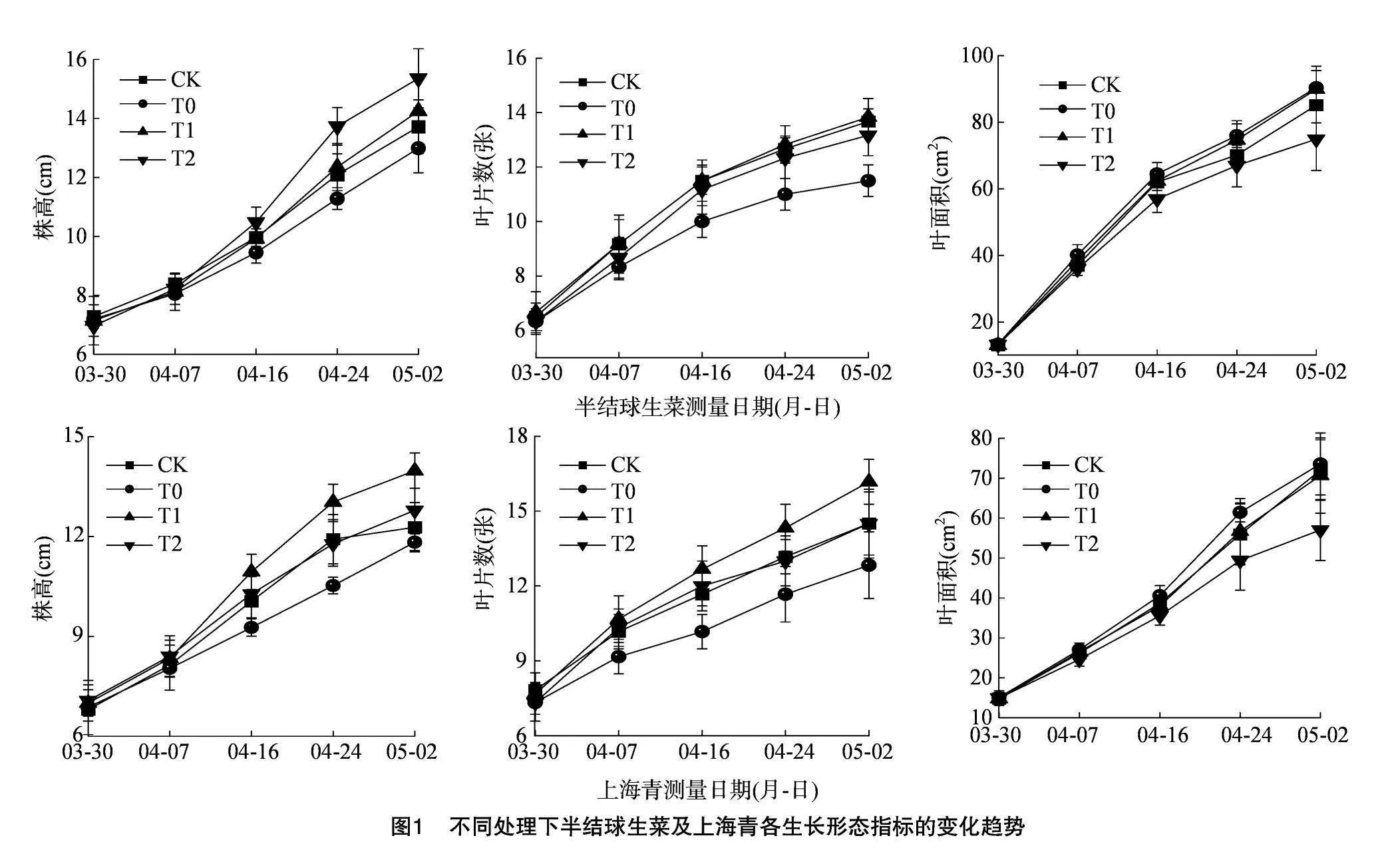
由图1可知,8 h/d光照周期下的半结球生菜及上海青植株的株高及叶片数生度情况均不及同期其他处理组,而20 h/d光照周期下的半结球生菜及上海青植株的叶面积均小于同期其他处理组。由图2可知,光照时长由8 h/d延长至16 h/d,半结球生菜的叶片数及株高分别提升20.3%和9.7%,上海青叶片数及株高分别提升26.0%和18.2%。光照时长由16 h/d延长至20 h/d,半结球生菜株高提升7.8%,叶片数及叶面积分别减小4.8%和16.7%,上海青株高、叶片数及叶面积均分别减小8.5%、10.3%和19.3%。由图3可知,12 h/d及16 h/d光照周期下的半结球生菜及上海青植株在生长表型上体现出更为健康的特征。
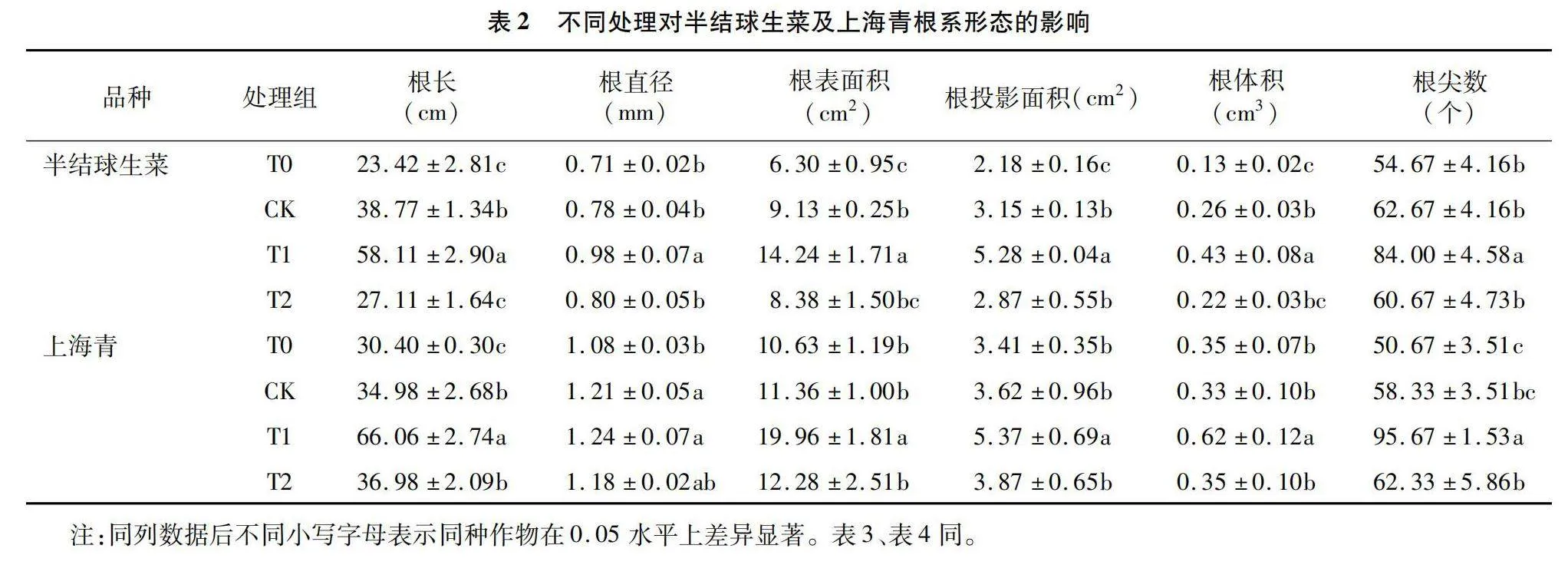
2.2 不同光照周期与强度对半结球生菜及上海青根系形态的影响
根系是植株吸收水分和营养物质的重要器官,其生长情况影响肥料吸收利用效率和自身抗逆性。由表2可知,16 h/d光照周期下的半结球生菜及上海青具有最优根系发育水平,光照周期由8 h/d延长至16 h/d,半结球生菜的根长、根体积与根尖数分别增长148.1%、230.8%和53.6%,上海青分别增长117.3%、77.1%和88.8%。但继续延长光照周期则使得二者的根系生长受到抑制。
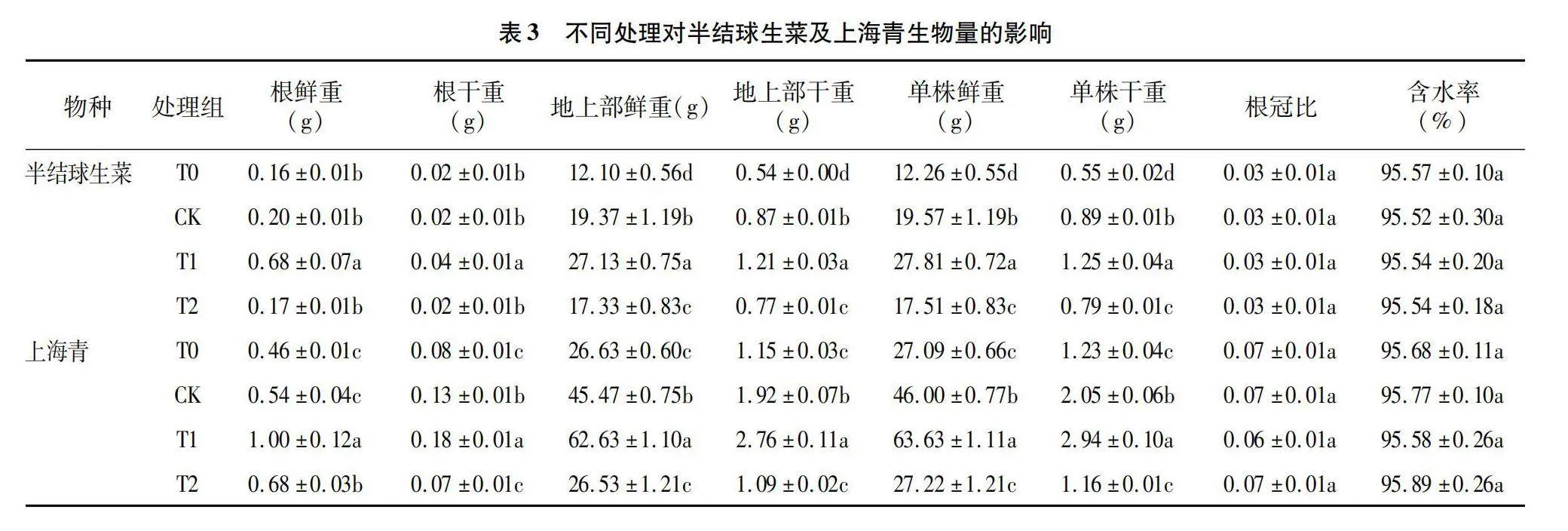
2.3 不同光照周期与强度对半结球生菜及上海青生物量的影响
由表3可知,与根系形态指标的变化趋势一致,16 h/d光照周期下的半结球生菜及上海青根干、鲜重及地上部干、鲜重均显著高于其他处理(P<0.05),而二者各组间根冠比及含水率无显著性差异。相同DLI下,16 h/d光照周期下半结球生菜的单株干重相较8、12、20 h/d分别增长127.3%、40.4%和58.2%,而16 h/d光照周期下上海青的单株干重较8、12、20 h/d分别增长139.0%、43.4%和153.4%。
2.4 不同光照周期与强度对半结球生菜及上海青电能利用效率的影响
由图4可知,相同DLI下,一定范围内延长光照周期、降低光照度可有效提升半结球生菜及上海青电能利用效率。当光照周期由8 h/d延长至16 h/d,半结球生菜及上海青的电能利用效率分别提升126.3%和139.3%,而随着光照周期由16 h/d延长至20 h/d,二者的电能利用效率均分别下降8.9%和36.1%。
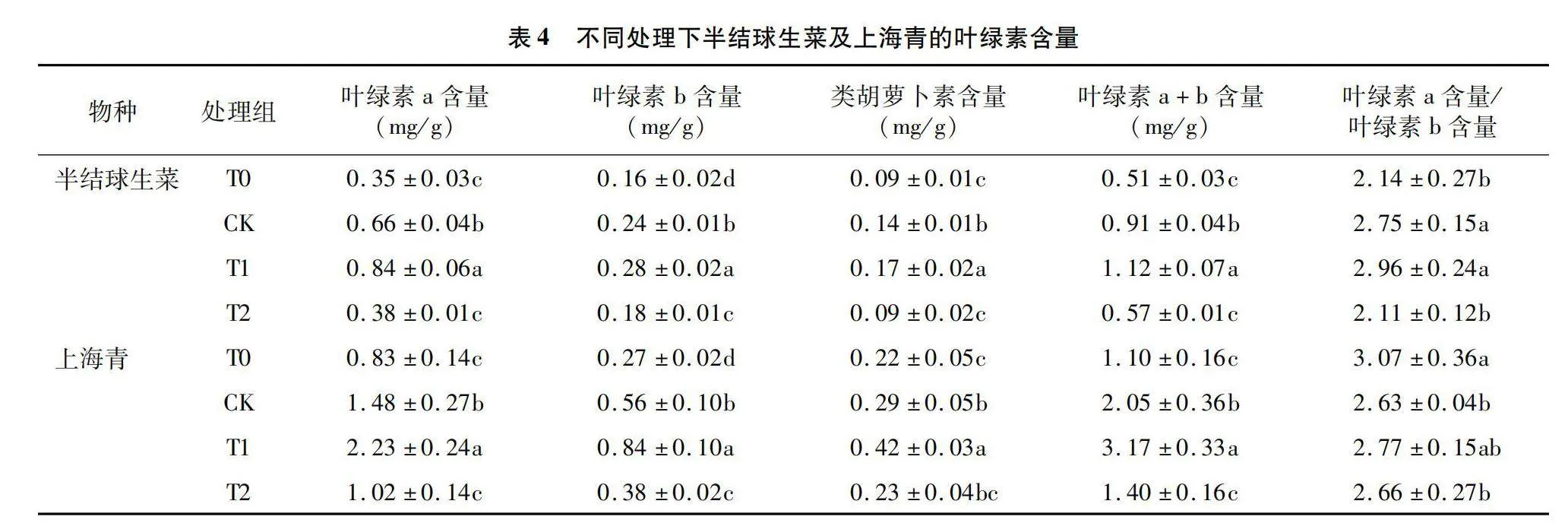
2.5 不同光照周期与强度对半结球生菜及上海青叶绿素含量的影响
由表4可知,16 h/d光照周期下的半结球生菜及上海青叶绿素a、叶绿素b、类胡萝卜素和叶绿素a+b含量在各组中具有最高水平。随着光照周期由8 h/d延长至16 h/d,二者的叶绿素a+b含量分别增加119.6%和188.2%,且12 h/d和16 h/d光照周期下的半结球生菜叶绿素a含量/叶绿素b含量值较高,可知在当前光照条件下能较好地满足其进行光合作用的条件。随着光照度的削弱,上海青的叶绿素a含量/叶绿素b含量值降低,其弱光状态下的光能利用能力得到提高。
2.6 不同光照周期与强度对半结球生菜及上海青生理生化指标的影响
2.6.1 对品质相关性状的影响
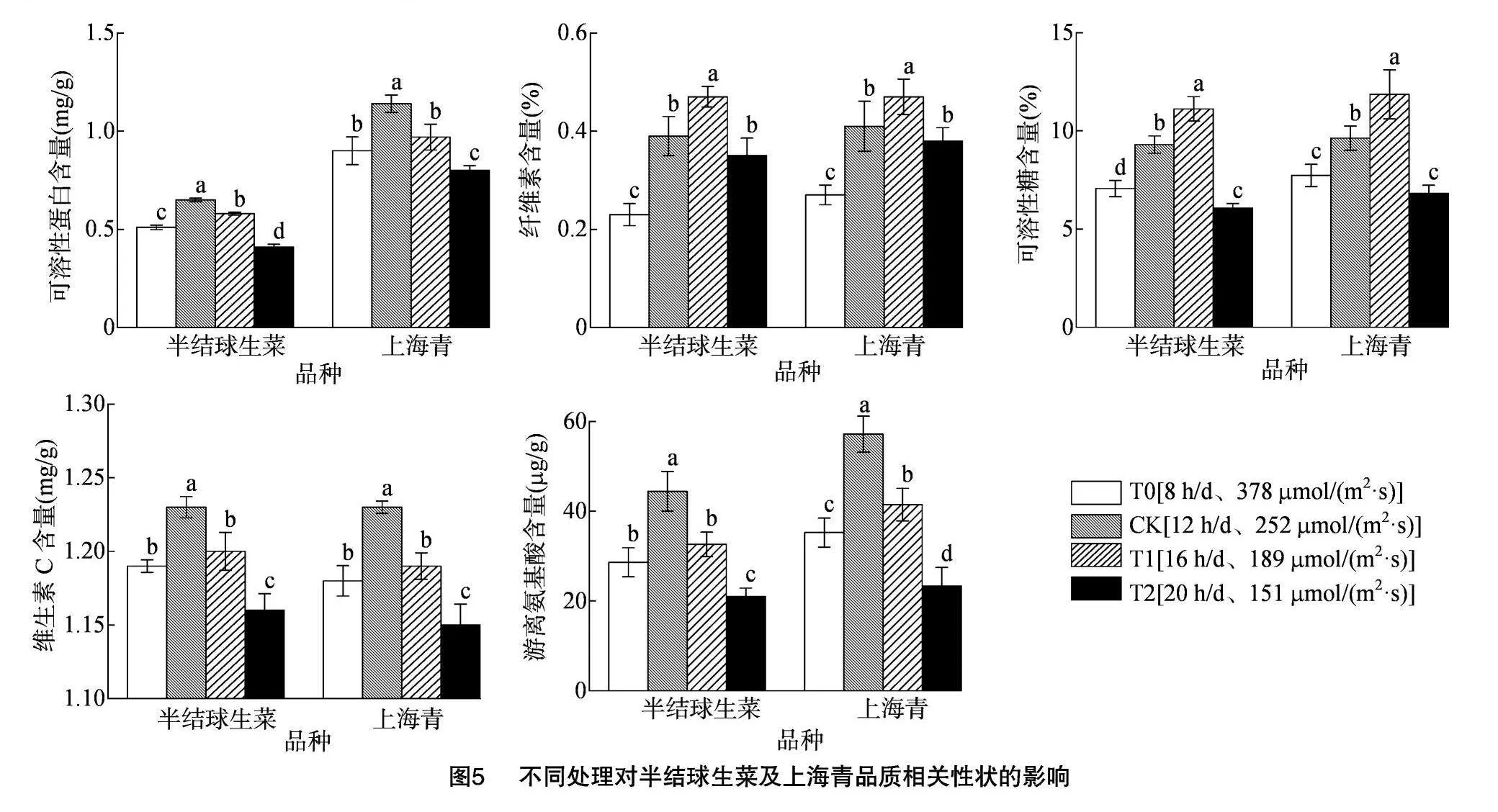
由图5可知,不同光照周期与强度下半结球生菜与上海青品质指标的变化趋势具有一致性。12 h/d光照周期下的半结球生菜可溶性蛋白、游离氨基酸及维生素C含量在各组中显著最高(P<0.05),相较8 h/d光照周期分别提升58.5%、111.3%和6.0%,而16 h/d相较8 h/d光照周期分别提升41.5%、55.2%和3.4%。与之相似,12 h/d光照周期下的上海青可溶性蛋白、游离氨基酸及维生素C含量同样在各组中显著最高,较8 h/d光照周期分别提升42.5%、145.1%和7.0%,而16 h/d相较8 h/d光照周期分别提升21.3%、77.8%和3.5%。而16 h/d光照周期下二者的可溶性糖和纤维素含量均显著高于其余各组,相较8 h/d光照周期,半结球生菜分别提升83.2%和34.3%,上海青分别提升74.2%和23.7%。
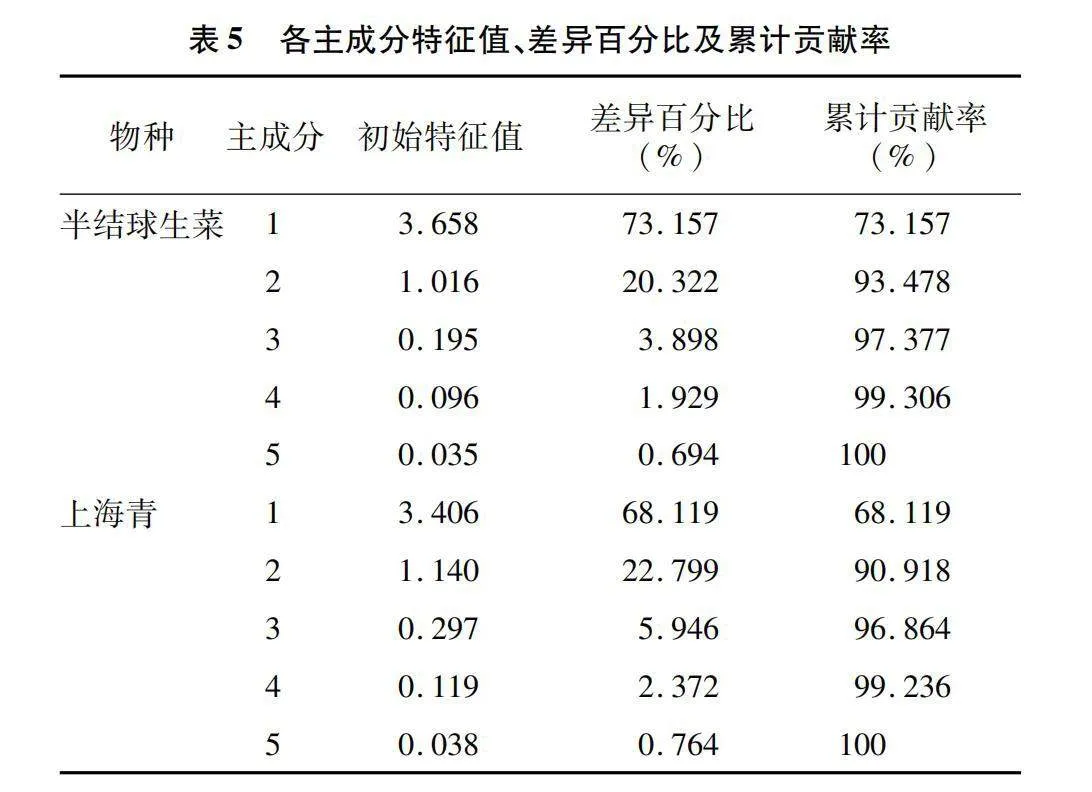
2.6.2 半结球生菜及上海青营养品质的主成分分析
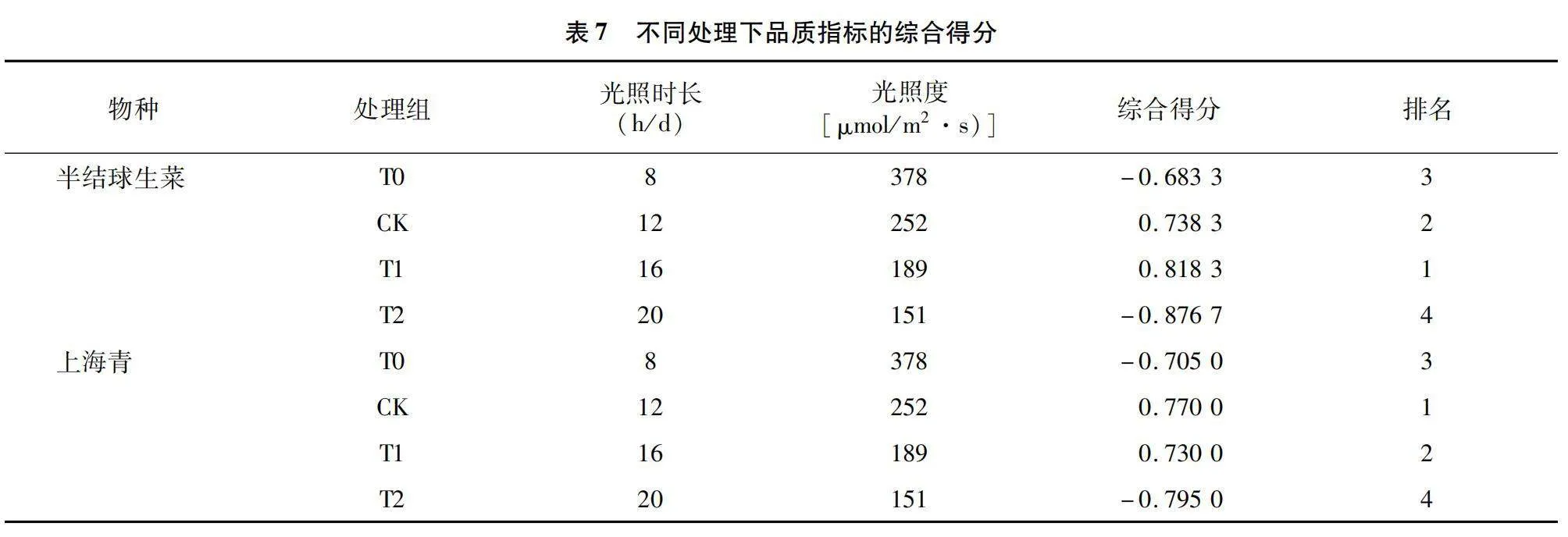
本研究将可溶性糖、维生素C、可溶性蛋白、游离氨基酸及纤维素含量的积累水平作为评价半结球生菜及上海青营养品质的影响因素,并对其进行主成分分析。由表5可知,半结球生菜主成分1(PC1)和主成分2(PC2)累计贡献率达到93.478%,上海青累计贡献率达到90.918%,可足够代表品质指标的信息。结合表6各品质指标成分矩阵信息可知,二者的维生素C、可溶性蛋白、游离氨基酸及可溶性糖在PC1上的载荷系数较大,与PC1具有较强的正向关系;而纤维素含量在PC2上荷载系数大,与PC2有正向关系。结合图6-a、图6-b可知,12 h/d与16 h/d光照周期下的半结球生菜与上海青品质水平均具有较好的得分趋势。由表7可知,通过量化分析,相同DLI下,半结球生菜在16 h/d、189 μmol/(m2·s)光照组合下具有最优营养品质,上海青则在12 h/d、252 μmol/(m2·s)光照组合下具有最佳营养品质。
2.6.3 对抗氧化指标的影响
由图7可知,当光照周期为8 h/d,光照度为378 μmol/(m2·s)时,半结球生菜及上海青的MDA含量均显著大于其他光照处理组(P<0.05)。随着光照度由151 μmol/(m2·s)增加至378 μmol/(m2·s),半结球生菜及上海青的MDA含量逐渐增加,分别提升41.3%和71.4%。
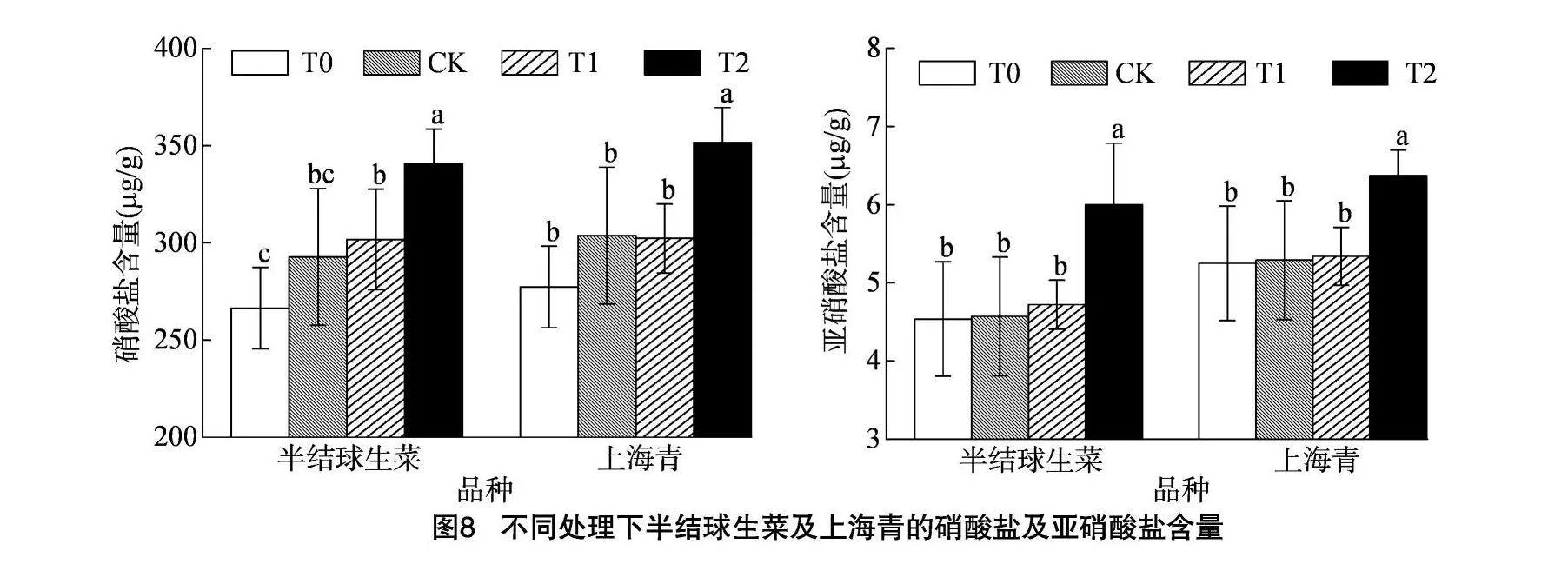
2.6.4 对硝酸盐及亚硝酸盐积累的影响
由图8可知,相同DLI下,半结球生菜及上海青植株内硝酸盐和亚硝酸盐的含量均随光照周期的延长而升高,20 h/d光照周期下的硝酸盐及亚硝酸盐含量均显著高于其他光照处理组(P<0.05)。随着光照周期由8 h/d延长至20 h/d,半结球生菜硝酸盐和亚硝酸盐含量分别升高27.8%和32.2%,上海青硝酸盐和亚硝酸盐含量分别升高26.7%和21.3%。
3 讨论
有研究表明,DLI一定时,16 h/d光照周期下的草莓幼苗相较12 h/d具有更快的生长速度,且植株生长形态对不同光照周期的响应机制因品种而异[17,21]。本试验同样发现相同DLI下,将光照时间由8 h/d延长至16 h/d,半结球生菜及上海青植株生长态势更佳,有助于缩短其种植周期;当光照周期延长至20 h/d,即光照度削弱至151 μmol/(m2·s)时,半结球生菜出现徒长现象,而上海青株高则呈下降趋势,考虑是二者生长发育中光照需求差异所致。与Yan等对黄瓜幼苗的研究结论[22]相似,本试验发现相同DLI下,随着光照时间由8 h/d延长至16 h/d,半结球生菜和上海青的根系生长更加旺盛,利于增强作物的肥料汲取效率,提高抗逆性,但继续延长光照周期致使光照度降低到一定水平时,根系生长受到抑制。前人研究指出,相同DLI下,21 h/d光照周期下的生菜单株干重较12 h/d光照周期增加28%[9]。本试验同样发现将光照周期由8 h/d延长至16 h/d,可显著促进半结球生菜及上海青的生物量积累,实现增产。但二者在20 h/d光照周期下的单株干重均显著小于12 h/d,考虑与本试验DLI值下光照度过低有关。
叶绿素含量是反映植株光合性能的重要指标,直接影响到光合作用的强弱[23]。本试验发现相同DLI下,光照周期由8 h/d延长至16 h/d,可提升半结球生菜及上海青叶绿素含量,从而提高叶片光合同化效率,缓解光抑制现象,这与李志鑫等的研究结果[24]相似。延长光照时间有助于光合色素的积累和电子传递速率的提高,当DLI相同时,延长光照时间在弥补光照度削弱对植物造成影响的同时,能进一步提高其光能利用率,促进其生长[7,25]。
前人在对生菜的研究中发现,DLI一定时,22 h/d光照周期下生菜的电子传输速率较7 h/d增加74%[26]。本试验发现,光照周期由8 h/d延长至16 h/d后,半结球生菜及上海青的电能利用效率均显著升高,这与周璇的研究结果[16]相似。考虑是延长光照周期并削弱光照度促使了更多的电子经由光系统Ⅱ(PSⅡ)集成,加之光合活动时间与光照时间的同步延长,从而实现光能输入的提高[8,27-28]。但当光照周期延长至20 h/d时,则可能由于光照度过低而开始下降。
有研究指出,茄子幼苗中可溶性糖含量及番茄中的可溶性蛋白含量随着光照时间的延长而升高[29-30],而生菜中的维生素C含量则随着光照度的削弱而提高[31]。本试验结果与之相似,并发现相同DLI下,随着光照周期的进一步延长,光照度逐渐降低于植株光补偿点,半结球生菜及上海青的品质指标变化由升高转为下降。通过主成分分析发现,相同DLI下,随着光照组合的变化,半结球生菜与上海青的生理品质指标变化趋势虽具相似性,但相同光照组合下二者的综合品质排名不同,考虑是二者间各品质指标在不同光照条件下变化幅度的差异所致,即不同作物的综合营养品质对光照组合的响应具有差异性。植株中MDA含量的高低常被用来衡量其受氧化损伤的程度[32]。本试验发现光照周期为8 h/d时,半结球生菜和上海青因高光照度而产生氧化损伤,并因此降低生长水平,这与Zheng等的研究结果[33]相似。蔬菜硝酸盐及亚硝酸盐具有强光下积累低而弱光下积累较多的特性[34]。本试验发现,光照周期为20 h/d时,生菜及上海青的硝酸盐及亚硝酸盐大量积累,对作物品质产生一定影响。
4 结论
综上,在完全可控的环境中,相同DLI下,合理延长光照周期、降低光照度利于促进半结球生菜及上海青植株的生长发育,缩短种植周期,提高作物产量,并提升营养品质,但需要注意弱光条件下作物硝酸盐、亚硝酸盐含量的积累。本试验条件下,16 h/d、189 μmol/(m2·s)的光照组合可帮助提高半结球生菜及上海青产量,同时提升半结球生菜营养品质,而12 h/d、252 μmol/(m2·s)的光照组合利于上海青营养品质的提高。
参考文献:
[1]孙 锦,高洪波,田 婧,等. 我国设施园艺发展现状与趋势[J]. 南京农业大学学报,2019,42(4):594-604.
[2]孙祥瑞,张尚卿,岳艳丽,等. 越冬蔬菜植物生长灯补光措施效果评价[J]. 中国蔬菜,2013(9):45-47.
[3]王 君,杨其长,仝宇欣. 红蓝光下光强对生菜电能、光能利用效率及品质的影响[J]. 中国农业大学学报,2016,21(8):59-66.
[4]宋 羽. 株间LED补光与温室番茄光能利用特性及生长发育关系的研究[D]. 北京:中国农业大学,2017:88-89.
[5]薛国萍,姜 伟,付崇毅,等. LED光源对草莓生长发育影响的研究进展[J]. 北方农业学报,2017,45(5):108-114.
[6]祁娟霞,韦 峰,董 艳,等. 不同补光时间对日光温室甜瓜生长发育的影响[J]. 浙江农业学报,2016,28(6):979-983.
[7]MaoH P,Hang T,Zhang X D,et al. Both multi-segment light intensity and extended photoperiod lighting strategies,with the same daily light integral,promoted Lactuca sativa L. growth and photosynthesis[J]. Agronomy,2019,9(12):857.
[8]Lefsrud M G,Kopsell D A,Kopsell D E,et al. Kale caroteff7ntBh2bNUJbROYHYkwGA==noids are unaffected by,whereas biomass production,elemental concentrations,and selenium accumulation respond to,changes in selenium fertility[J]. Journal of Agricultural and Food Chemistry,2006,54(5):1764-1771.
[9]WeaverG,van Iersel M W. Longer photoperiods with adaptive lighting control can improve growth of greenhouse-grown ‘Little Gem’ lettuce (Lactuca sativa)[J]. HortScience,2020,55(4):573-580.
[10]靳志勇,王硕硕,刘 娜,等. 不同补光时间和方式对设施大蒜鳞茎膨大和品质的影响[J]. 园艺学报,2014,41(12):2411-2418.
[11]Ji F,Wei S Q,Liu N,et al. Growth of cucumber seedlings in different varieties as affected by light environment[J]. International Journal of Agricultural and Biological Engineering,2020,13(5):73-78.
[12]Yan Z N,He D X,Niu G H,et al. Evaluation of growth and quality of hydroponic lettuce at harvest as affected by the light intensity,photoperiod and light quality at seedling stage[J]. Scientia Horticulturae,2019,248:138-144.
[13]BowesG,Ogren W L,Hageman R H. Light saturation,photosynthesis rate,RuDP carboxylase activity,and specific leaf weight in soybeans grown under different light intensities[J]. Crop Science,1972,12(1):77-79.
[14]ChowW S. Photoprotection and photoinhibitory damage[M]//Advances in molecular and cell biology. Amsterdam:Elsevier,1994,10:151-196.
[15]Faust J E,Heins R D. Modeling leaf development of the African violet (Saintpaulia ionantha Wendl.)[J]. Journal of the American Society for Horticultural Science,1993,118(6):747-751.
[16]周 璇. 光照时长与光照强度对叶用莴苣生长及生理特性的影响[D]. 泰安:山东农业大学,2022:49-50.
[17]Kelly N,Choe D,Meng Q W,et al. Promotion of lettuce growth under an increasing daily light integral depends on the combination of the photosynthetic photon flux density and photoperiod[J]. Scientia Horticulturae,2020,272:109565.
[18]李合生. 植物生理生化实验原理和技术[M]. 北京:高等教育出版社,2000:134-211.
[19]叶瑞洪,林 谦. 苯酚分光光度法测定食品中硝酸盐与亚硝酸盐[J]. 闽江学院学报,2010,31(2):111-114.
[20]Kozai T. Resource use efficiency of closed plant production system with artificial light:concept,estimation and application to plant factory[J]. Proceedings of the Japan Academy(Series B,Physical and Biological Sciences),2013,89(10):447-461.
[21]Tsuruyama J,Shibuya T. Growth and flowering responses of seed-propagated strawberry seedlings to different photoperiods in controlled environment chambers[J]. HortTechnology,2018,28(4):453-458.
[22]Yan Z N,Wang L,Wang Y F,et al. Morphological and physiological properties of greenhouse-grown cucumber seedlings as influenced by supplementary light-emitting diodes with same daily light integral[J]. Horticulturae,2021,7(10):361.
[23]Causin H F,Jauregui R N,Barneix A J. The effect of light spectral quality on leaf senescence and oxidative stress in wheat[J]. Plant Science,2006,171(1):24-33.
[24]李志鑫,李 欣,蔡冼峰,等. 相同DLI下不同光照周期对水培罗勒风味物质的影响[J]. 青岛农业大学学报(自然科学版),2023,40(2):84-90,111.
[25]Zhou J,Wang J Z,Hang T,et al. Photosynthetic characteristics and growth performance of lettuce (Lactuca sativa L.) under different light/dark cycles in mini plant factories[J]. Photosynthetica,2020,58(3):740-747.
[26]Elkins C,van Iersel M W. Longer photoperiods with the same daily light integral improve growth of Rudbeckia seedlings in a greenhouse[J]. HortScience,2020,55(10):1676-1682.
[27]Shen Y Z,Guo S S,Ai W D,et al. Effects of illuminants and illumination time on lettuce growth,yield and nutritional quality in a controlled environment[J]. Life Sciences in Space Research,2014,2:38-42.
[28]Liu Y,Wang X H,Fang Y,et al. The effects of photoperiod and nutrition on duckweed (Landoltia punctata) growth and starch accumulation[J]. Industrial Crops and Products,2018,115:243-249.
[29]陈 敏,李海云. 不同光周期对茄子幼苗生长的影响[J]. 北方园艺,2010(16):53-55.
[30]刘 露. 不同留果穗数和不同补光时间对番茄品质及产量的影响[D]. 杨凌:西北农林科技大学,2018:48-49.
[31]刘 杰,胡笑涛,王文娥,等. 光强和光周期对水培生菜光合及叶绿素荧光特性的影响[J]. 西南农业学报,2019,32(8):1784-1790.
[32]Mittler R. Oxidative stress,antioxidants and stress tolerance[J]. Trends in Plant Science,2002,7(9):405-410.
[33]Zheng Y J,Zhang Y T,Liu H C,et al. Supplemental blue light increases growth and quality of greenhouse pak choi depending on cultivar and supplemental light intensity[J]. Journal of Integrative Agriculture,2018,17(10):2245-2256.
[34]钱 和,蒋 将,陈正行.蔬菜中硝酸盐与亚硝酸盐的积累规律与控制方法[J]. 食品科技,2007,32(1):64-67.
