
三种诱导剂对草地早熟禾低温抗性的影响
2024-11-05 00:00马福钦韩悦李文秀崔林雨张世林董文科
草地学报 2024年10期








摘要:为了分析不同类型诱导剂对草地早熟禾(Poa pratensis)低温抗性的影响,本试验以草地早熟禾‘巴润’为材料,研究低温胁迫(白天4℃/黑夜0℃,每天光照14 h,黑暗10 h,光强3500~4000 Lux,相对湿度60%)下喷施不同浓度的壳聚糖(0,20,50,100,200,300,400和500 μmol·L-1)、甜菜碱(0,20,40,60,80,100,120和140 μmol·L-1)和褪黑素(0,25,50,100,200,300,400和500 μmol·L-1)对草地早熟禾生理特性的影响。结果表明:喷施适宜浓度的诱导剂均能有效缓解草地早熟禾幼苗受低温胁迫危害,三种诱导剂处理的草地早熟禾幼苗相对电导率和丙二醛含量均低于低温对照幼苗,并呈先降后升的趋势;叶绿素、可溶性糖、可溶性蛋白和游离脯氨酸含量呈先升后降趋势,抗氧化酶活性显著提高;通过计算各指标抗寒隶属函数值并排序发现,三种诱导剂中以300 μmol·L-1浓度的褪黑素的处理效果最佳。
关键词:草地早熟禾;诱导剂;低温胁迫;生理特性
中图分类号:S945.79 文献标识码:A 文章编号:1007-0435(2024)10-3194-11
Effects of Three Inducers on Low Temperature Resistance of Poa pratensis
MA Fu-qin, HAN Yue, LI Wen-xiu, CUI Lin-yu, ZHANG Shi-lin, DONGWen-ke*
(College of Pratacultural Science, Gansu Agricultural University/Key Laboratory of Grassland Ecosystem/Ministry of Education/
Pratacultural Engineering Laboratory of Gansu Province/Sino-U.S. Center for Grassland Ecosystem Sustainability/Key Laboratory of Forage Gerplasm Innovationand Variety Breeding of the Ministry of Agriculture and Rural Affairs, Lanzhou, Gansu Province 730070,China)
Abstract:In order to analyze the effects of different types of inducers on the cold resistance of Kentucky bkuegrass (Poa pratensis),the experiment was conducted to study the effects of different concentrations of chitosan (0,20,50,100,200,300,400 and 500 μmol·L-1),betaine(0,20,40,60,80,100,120 and 140 μmol·L-1) and melatonin(0,25,50,100,200,300,400 and 500 μmol·L-1) on the physiological characteristics of Poa pratensis ‘Baron’ under low temperature stress(day 4℃/night 0℃,light 14 hours a day,darkness 10 hours,light intensity 3500-4000 Lux,relative humidity 60%). The results showed that spraying the appropriate concentration of inducers could effectively alleviate the damage of P. pratensis seedlings under low temperature stress. The relative conductivity and malondialdehyde content of P. pratensis seedlings treated with three kinds of inducers were lower than that of control seedlings under low temperature stress,and showed a trend of first decreasing and then increasing. The contents of chlorophyll,soluble sugar,soluble protein and free proline increased first and then decreased,and the activity of antioxidant enzymes increased significantly. The cold resistance membership function of each index was calculated and sorted. It was found that 300 μmol·L-1 of melatonin had the best treatment effect.
Key words:Poa pratensis;Inducer;Low temperature stress;Physiological characteristics
收稿日期:2024-01-31;修回日期:2024-05-27
基金项目:国家自然科学基金(32201457);草业生态系统教育部重点实验室(甘肃农业大学)开放课题(KLGE202210)资助
作者简介:
马福钦(1998-),男,汉族,甘肃金昌人,硕士研究生,主要从事牧草及草坪草种质资源育种研究,E-mail:Mafq2023@163.com;*通信作者Author of corresponfence,E-mail:dongwk2021@163.com
随着社会的发展,草坪成为人类日常生活和现代文明不可或缺的一部分,具有维持生态系统、美化环境、休憩、游玩、运动等作用,是城市绿化中的重要成分[1]。草地早熟禾(Poa pratensis )是城市草坪建植中主要使用的冷季型草坪草之一,具有极深的蓝绿色,质地细嫩,具耐热、耐寒、耐旱,抗病性较好等优点[2],在草坪建植以及生态环境治理中广泛应用。
低温是一种最常见的逆境胁迫因子,影响植物的生长发育,限制植物在自然界中的地理分布。当植物受到低温胁迫时,其细胞膜系统受破坏,矿质营养吸收受到影响,生物合成速率降低[3],光能吸收减少,许多生理代谢功能受到阻碍,严重时可造成植株死亡[4]。喷施某些化学物质可以提高植物抗寒性,这类化学物质统称为抗寒诱导剂。壳聚糖(Chitosan,CTS)是甲壳质脱乙酞化后得到的一种聚氨基葡萄糖,壳聚糖及其衍生物能提高植物的抗寒能力,在低温条件下促进植物生长,提高种子的发芽率,降低细胞膜透性及丙二醛(Malondialdehyde,MDA)的积累,提高植物中渗透物质的含量,减小低温胁迫对幼苗伤害[5-6]。已有研究发现,将壳聚糖作为外源物质喷施于棉花(Gossypium)[7]、水稻(Oryza sativa)[8]和辣椒(Capsicum annuum)[9]等的叶片,均能提高其低温抗性。褪黑素(Melatonin,MT)是一种低分子量的吲哚类化合物,在植物中的主要作用是作为活性氧自由基(Reactive Oxygen Species,ROS)清除剂,起到抵御植物内部和环境氧化应激的第一层保护作用,能够增强植物对非生物胁迫的抵性,延缓植物叶片的衰老[10]。喷施外源褪黑激素使低温胁迫下玉米(Zea mays)幼苗渗透物质含量和抗氧化酶活性增加[11];减轻了低温胁迫对黄瓜(Cucumis sativus)[12]的光合机构的损伤,降低了低温造成的膜质过氧化程度,能够增强其抵抗低温的能力,恢复幼苗正常生长。甜菜碱(Betaine,BT)是一种季铵型水溶性生物碱,广泛存在于动植物及微生物中,是维持细胞稳定的重要渗透调节物质[13]。当植物处于不良环境条件下时,植物体内的甜菜碱会大量积累,用以减轻逆境胁迫对细胞膜的损伤,并保护主要酶活性以及稳定光合作用中多肽,从而提高植物适应不良环境的能力[14]。前人研究发现,叶面喷施外源甜菜碱能够缓解低温胁迫下番茄(Solanum lycopersicum )幼苗的叶绿素含量下降,增强渗透调节能力,提高其耐低温能力[15]。
目前已有低温胁迫下喷施外源壳聚糖、褪黑素和甜菜碱在狗牙根(Cynodon dactylon)[16]、小麦(Triticum aestivum)[17]、水稻[18]等植物的研究,并获得了较好的效果。为此,本试验选择壳聚糖、甜菜碱、褪黑素为诱导剂,探索喷施不同浓度诱导剂对草地早熟禾幼苗低温抗性的影响,旨在筛选出最佳诱导剂及其最适浓度,为提高草地早熟禾低温抗性提供理论依据。
1 材料和方法
1.1 试验材料
供试材料草地早熟禾品种‘巴润’种子由北京克劳沃草业技术开发中心提供、三种诱导剂壳聚糖、甜菜碱、褪黑素由上海源叶生物科技有限公司提供。
1.2 试验方法
1.2.1 幼苗种植与培养 选取颗粒饱满、完整的草地早熟禾‘巴润’种子,先用70%乙醇消毒30 s,再用20%的次氯酸钠消毒15 min后用蒸馏水冲洗5~6次,将种子晾干,均匀的撒在盛有清洗干净并高温烘干杀菌的沙土的育苗钵中,并用干沙覆盖,将其表面喷至湿润,每天喷水2次,保证沙子湿润度,并移至光照培养室[每天光照14 h,黑暗10 h,昼夜温度分别为(23±1)℃和(20±1)℃,相对湿度60%]。待幼苗长至2 cm左右时,需在沙基中浇灌Hoagland's营养液。
1.2.2 喷施诱导剂处理 待幼苗生长30 d时,选取形态和长势基本一致的草地早熟禾幼苗,用小型喷壶分别往植株叶片喷施不同浓度的三种诱导剂,壳聚糖设置为0,20,50,100,200,300,400和500 μmol·L-1,甜菜碱设置为0,20,40,60,80,100,120和140 μmol·L-1,褪黑素设置为0,25,50,100,200,300,400和500 μmol·L-1,并设置常温对照组(CK),每个处理设置三次重复;喷施程度以叶面均匀布满雾状水滴为宜;每24 h喷洒叶片1次,连续喷3次;缓苗1天后将幼苗转入4℃/0℃(白天/黑夜,每天光照14 h,黑暗10 h,光强3500~4000 Lux,相对湿度60%)光照培养箱中低温胁迫处理,72 h后采样测定相关生理指标。
1.3 测定指标及方法
1.3.1 普通生理指标的测定 叶片相对电导率参照魏臻武等[19]的方法;叶绿素含量采用丙酮法[20]测定;丙二醛(Malondialdehyde,MDA)含量采用硫代巴比妥酸法[21]测定;游离脯氨酸(Proline,Pro)含量采用磺基水杨酸提取法[22]测定;可溶性糖(Soluble sugar,SS)含量采用蒽酮法[23]测定;可溶性蛋白(Soluble protein,SP)含量采用考马斯亮蓝G-250比色法[24]测定。
1.3.2 抗氧化酶活性测定 超氧化物歧化酶(Superoxide dismutase,SOD)、过氧化物酶(Peroxidase,POD)和过氧化氢酶(Catalase,CAT)酶的活性,参照贺佳圆[25]的方法测定。
1.4 数据分析
SPSS 26.0软件进行统计分析,采用单因素ANOVA进行分析处理,Duncan’s新复极差法进行显著性方差分析;Microsoft Excel 2010用于绘图和数据处理。用模糊数学隶属函数法进行低温抗性综合评价[26],指标性状与低温抗性呈正相关时(Pro、SS、SP和叶绿素含量以及SOD、POD、CAT活性)使用正隶属函数隶属函数,指标性状与低温抗性呈负相关时(相对电导率和丙二醛含量)使用反隶属函数。公式如下:
正隶属函数公式为:U(X)=(X-X)/(X-X)
反隶属函数公式为:U(X) =1- (X-X)/(X-X)
式中,U(X)是隶属函数值;X为某一处理水平某指标测定值;X和X为所有参试水平系中某一指标中的最大值和最小值;最后将每个处理各抗寒指标的隶属函数值相加,求出平均值(D值)进行抗寒性排序。
2 结果与分析
2.1 低温胁迫下三种诱导剂对草地早熟禾幼苗相对电导率的影响
如图1所示,低温胁迫下,草地早熟禾幼苗的相对电导率较常温对照有显著上升,最大上升幅度为220.3%;叶面喷施不同类型诱导剂后,草地早熟禾幼苗的相对电导率均较低温对照(0 μmol·L-1)下降,但均显著高于CK(P<0.05)。其中与低温对照相比,叶面喷施200 μmol·L-1的壳聚糖、80 μmol·L-1的甜菜碱和300 μmol·L-1的褪黑素处理下草地早熟禾幼苗的相对电导率达到最小值,分别较低温对照降低了34.5%,32.2%,38.9%,300 μmol·L-1褪黑素处理的降低幅度最大。
2.2 低温胁迫下三种诱导剂对草地早熟禾幼苗丙二醛(MDA)含量的影响
如图2所示,低温胁迫下,低温对照的草地早熟禾幼苗MDA含量较CK显著上升,但随着不同类型诱导剂浓度的升高,草地早熟禾幼苗的MDA含量均呈先下降后上升的趋势。在200 μmol·L-1的壳聚糖、60 μmol·L-1的甜菜碱和100 μmol·L-1的褪黑素处理下草地早熟禾幼苗的MDA含量达到最小值,分别较低温对照降低了44.2%、27.2%、43.4%,其中200 μmol·L-1壳聚糖处理下的MDA含量降低幅度最大,但在褪黑素浓度为100 μmol·L-1时草地早熟禾幼苗的MDA含量最小。
2.3 低温胁迫下三种诱导剂对草地早熟禾幼苗叶绿素含量的影响
如图3所示,低温胁迫下,草地早熟禾幼苗的叶绿素含量变化幅度不显著,但较CK有显著的降低;且随着不同类型诱导剂浓度的升高,草地早熟禾幼苗的叶绿素含量均呈先上升后下降的趋势。在300 μmol·L-1的壳聚糖、100 μmol·L-1的甜菜碱和300 μmol·L-1的褪黑素处理下草地早熟禾幼苗的叶绿素含量达到最大值,分别较低温对照升高了3.9%,3.7%,3.5%,其中300 μmol·L-1壳聚糖处理的升高幅度最大。
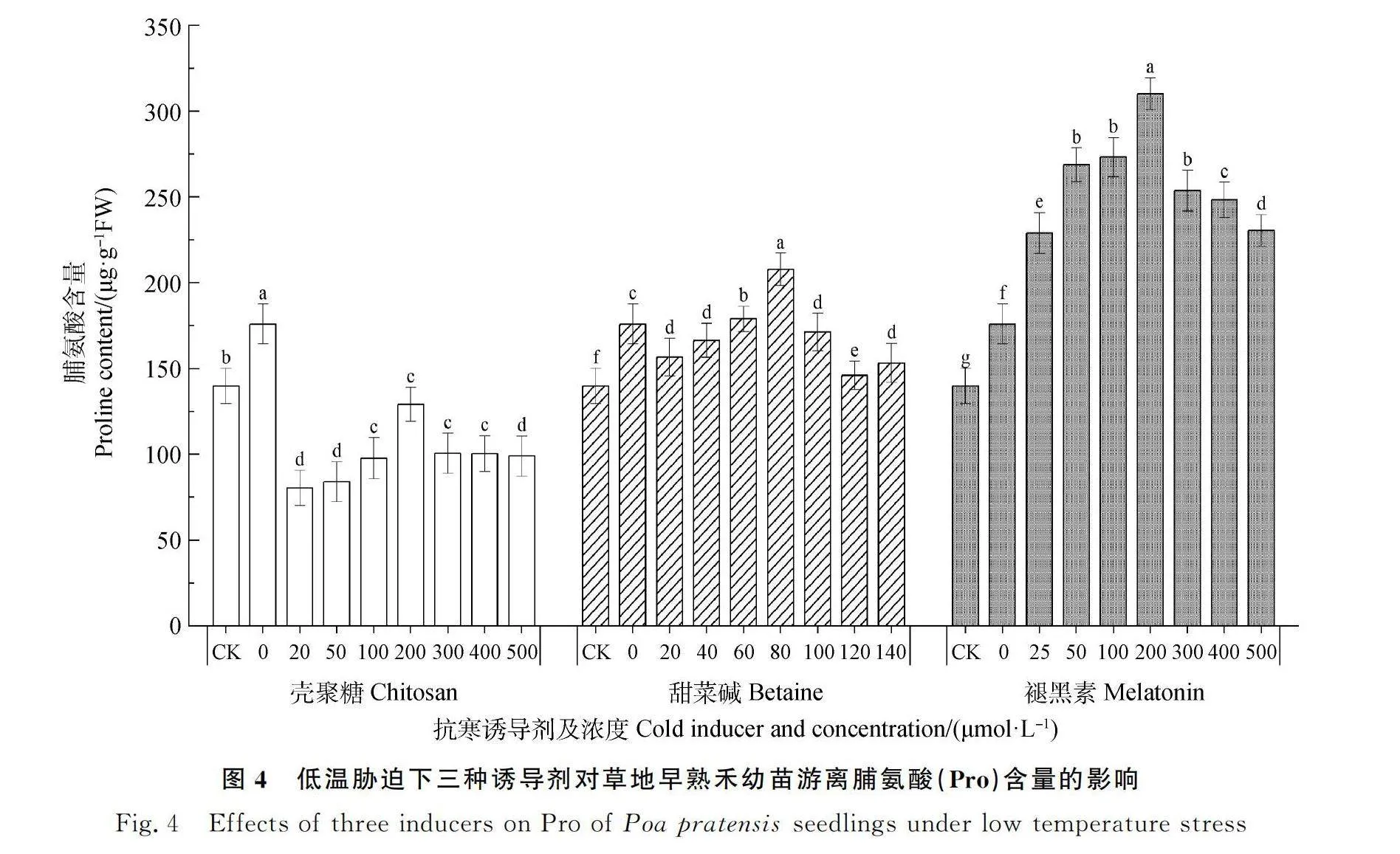
2.4 低温胁迫下三种诱导剂对草地早熟禾幼苗游离脯氨酸(Pro)含量的影响
如图4所示,低温胁迫下,随着不同类型诱导剂浓度的增加,草地早熟禾幼苗的Pro含量均呈先上升后下降的趋势。喷施壳聚糖处理的幼苗Pro含量均显著低于低温对照(P<0.05),甜菜碱处理的幼苗Pro含量在浓度为60~80 μmol·L-1时显著高于低温对照(P<0.05),褪黑素处理下幼苗Pro的含量均显著高于低温对照(P<0.05);壳聚糖抑制低温胁迫下草地早熟禾幼苗Pro的合成,而适宜浓度的甜菜碱和褪黑素能促进幼苗Pro的合成;且在80 μmol·L-1的甜菜碱和200 μmol·L-1的褪黑素处理下草地早熟禾幼苗的Pro含量达到最大值,分别较低温对照升高了18.1%、76.2%,其中200 μmol·L-1褪黑素的处理升高幅度最大,幼苗的Pro含量最高。
2.5 低温胁迫下三种诱导剂对草地早熟禾幼苗可溶性糖(SS)含量的影响
如图5所示,三种诱导剂处理对低温胁迫下草地早熟禾幼苗的SS含量有显著的影响,整体表现为随着不同类型诱导剂浓度的增加,草地早熟禾幼苗的SS含量均呈先上升后下降的趋势,但变化差异又因诱导剂浓度的差异而不尽相同。其中,低浓度的诱导剂促进低温下草地早熟禾幼苗SS含量的升高,而高浓度诱导剂抑制草地早熟禾幼苗的SS含量的升高,在200 μmol·L-1的壳聚糖、80 μmol·L-1的甜菜碱和300 μmol·L-1的褪黑素处理下草地早熟禾幼苗的SS含量达到最大值,分别较低温对照升高了14.4%,15.6%,29.2%,其中300 μmol·L-1褪黑素处理的升高幅度最大。
2.6 低温胁迫下三种诱导剂对草地早熟禾幼苗可溶性蛋白(SP)含量的影响
如图6所示,低温胁迫下,经不同类型诱导剂处理后的草地早熟禾幼苗SP含量显著高于常温对照和低温对照(P<0.05),同一类型诱导剂处理后的SP含量在不同浓度下存在差异,在300 μmol·L-1的壳聚糖、100 μmol·L-1的甜菜碱和300 μmol·L-1的褪黑素处理下草地早熟禾幼苗的SP含量达到最大值,分别较低温对照升高了18.5%,27.4%,43.6%,其中300 μmol·L-1褪黑素处理的升高幅度最大,但随着不同类型诱导剂浓度的增加,草地早熟禾幼苗的SP含量均呈先上升后下降的趋势,且褪黑素处理后SP含量显著高于其他两组。
2.7 低温胁迫下三种诱导剂对草地早熟禾幼苗超氧化物歧化酶(SOD)活性的影响
如图7所示,低温胁迫下三种诱导剂对草地早熟禾幼苗SOD活性有显著的影响,随着不同类型诱导剂浓度的增加,草地早熟禾幼苗的SOD活性均呈先上升后下降的趋势。无低温胁迫时草地早熟禾幼苗SODXfwmlcprIa64PpWfUs2y7A==活性显著低于低温胁迫后;在200 μmol·L-1的壳聚糖、80 μmol·L-1的甜菜碱和300 μmol·L-1的褪黑素处理下草地早熟禾幼苗的SOD活性达到最大值,且显著高于低温对照(P<0.05),分别较低温对照升高了16.0%,14.8%,45.2%,其中褪黑素处理后SOD活性上升最为显著。
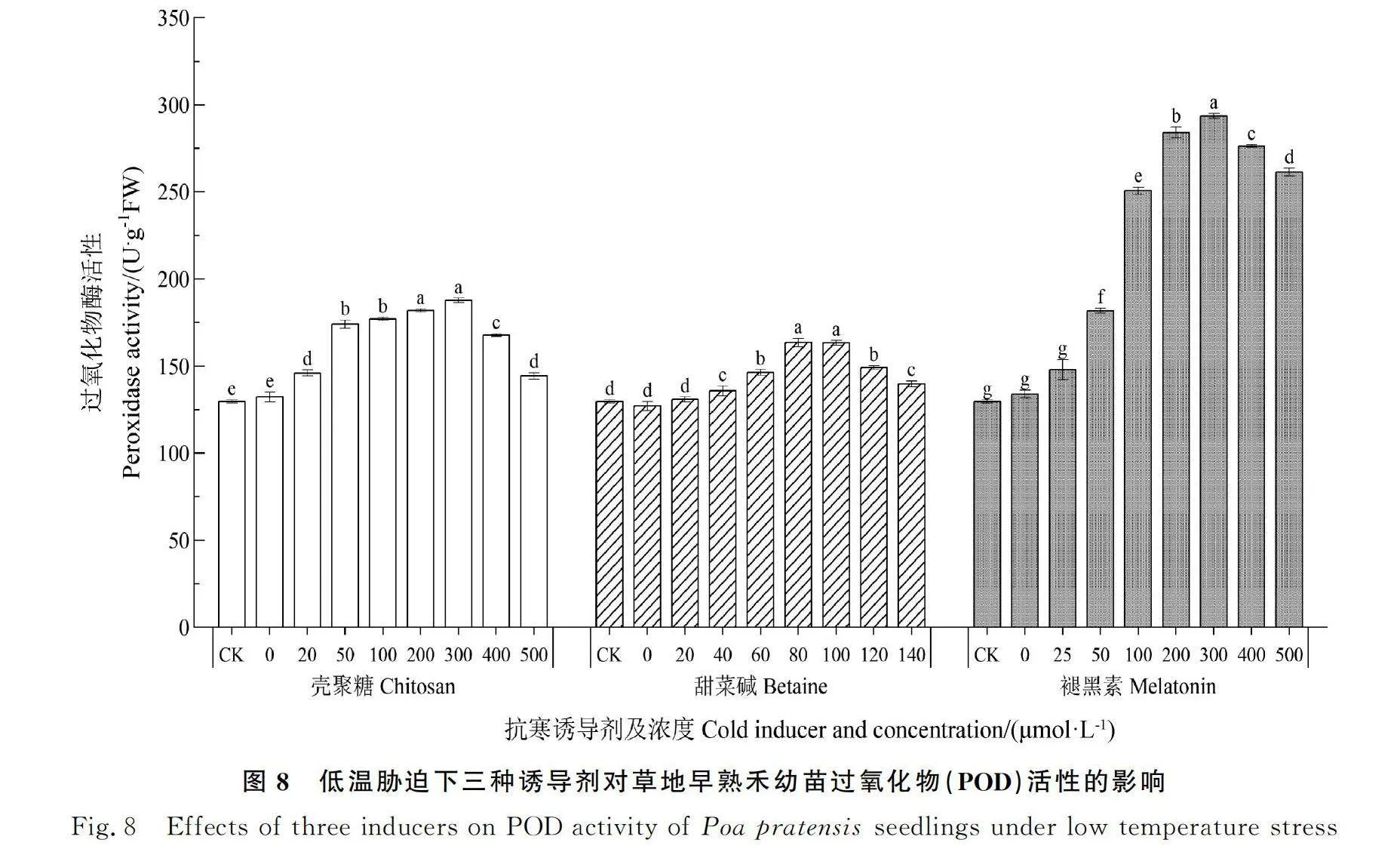
2.8 低温胁迫下三种诱导剂对草地早熟禾幼苗过氧化物酶(POD)活性的影响
如图8所示,随着不同类型诱导剂浓度的增加,草地早熟禾幼苗的POD活性均呈先上升后下降的趋势,但各类型诱导剂处理下差异较大。低温对照(0 μmol·L-1)和常温对照(CK)差异不显著,随着诱导剂浓度的升高,300 μmol·L-1的壳聚糖、80 μmol·L-1的甜菜碱和300 μmol·L-1的褪黑素处理下草地早熟禾幼苗的POD活性达到最大值,分别较低温对照升高了42.0%,28.7%,119.2%。在褪黑素浓度为400 μmol·L-1和500 μmol·L-1时POD活性仍显著高于CK和低温对照处理(P<0.05)。
2.9 低温胁迫下三种诱导剂对草地早熟禾幼苗过氧化氢酶(CAT)活性的影响
如图9所示,低温对照的CAT活性显著低于CK的活性(P<0.05);随着不同类型诱导剂浓度的增加,草地早熟禾幼苗的CAT活性均呈先上升后下降的趋势。在300 μmol·L-1的壳聚糖、100 μmol·L-1的甜菜碱和300 μmol·L-1的褪黑素处理下草地早熟禾幼苗的CAT活性达到最大值,分别较低温对照(0 μmol·L-1)升高了17.5%,12.8%,42.7%,其中300 μmol·L-1褪黑素处理的升高幅度最大。
2.10 草地早熟禾抗寒性综合评价
通过计算各指标抗寒隶属函数均值,对三种不同浓度诱导剂对草地早熟禾低温抗性的诱导效果进行评价,发现300 μmol·L-1的褪黑素处理下草地早熟禾抗寒评价值最大为0.880,抗寒性最强,其次是褪黑素浓度为200 μmol·L-1时;500 μmol·L-1的壳聚糖处理,抗寒评价值最小为0.200,抗寒性最弱(表1)。
3 讨论
低温胁迫使植物细胞膜脂过氧化,质膜透性改变或失去透性,MDA大量积累,胞内物质泄露,从而导致电解质升高,因此植物细胞膜脂过氧化程度及电解质外渗程度常用MDA含量和相对电导率来表现,代表胁迫对植物的损失程度。本研究发现,低温胁迫下未进行三种外源诱导剂(0 μmol·L-1)处理的草地早熟禾幼苗相对电导率、MDA相对于常温对照(CK)有明显phrnBwj3QI4IQrAg0PFGgQ==上升,而三种外源诱导剂处理后草地早熟禾幼苗的相对电导率和MDA有显著降低的趋势。这与练华山等[27]在壳聚糖缓解低温处理对丝瓜(Luffa aegyptiaca)幼苗的影响研究和尉欣荣等[28]在低温胁迫下褪黑素对多年生黑麦草(Lolium perenne)幼苗生长和抗氧化系统的作用中的研究结果一致。壳聚糖、甜菜碱和褪黑素三种诱导剂能缓解低温对草地早熟禾幼苗细胞膜脂的过氧化,保证细胞膜系统结构的完整性,从而提高了草地早熟禾幼苗的低温抗性。
植物在应对不利于其生长的环境胁迫时,细胞渗透物质增加,来维持低温环境下细胞的渗透平衡[29]。Pro、SS、SP作为重要的渗透调节物质和营养物质,其作用是提高细胞的保水能力,抑制了电解质外流,防止活性氧对膜脂和蛋白质的过氧化作用,保护生物膜,减轻细胞损伤程度[30]。朱云林等[31]研究发现,低温胁迫下壳聚糖可以使水稻幼苗渗透物质的含量升高,维持细胞正常酸碱度和稳定生物膜渗透性。本研究表明,低温胁迫增加了草地早熟禾幼苗体内Pro,SS和SP的含量,并且叶面喷施壳聚糖后Pro等渗透物质的含量随壳聚糖浓度的升高呈先升高后降低的趋势。这可能是由于适当浓度的壳聚糖可以破坏细胞膜的完整性,而正是由于这种破坏,才会诱发一系列有利于增强抵抗力的生理变化,激活酶促抗氧化系统和非酶促抗氧化系统[32]。本研究也表明,低温胁迫下草地早熟禾幼苗的渗透物质含量显著高于常温对照,并且在叶面喷施甜菜碱后这些物质的含量进一步的上升,说明甜菜碱可以提高草地早熟禾幼苗体内渗透物质的含量。其原因可能是,外源甜菜碱在提高渗透物质合成途径中关键酶的活性,促进酶的合成方面具有一定作用。白如意等[33]研究发现,低温胁迫下,100 μmol·L-1的褪黑素处理显著提高了南瓜幼苗中SS等物质的含量,减轻了低温胁迫对南瓜幼苗的伤害。本研究也发现叶面喷施褪黑素后,草地早熟禾幼苗的Pro、SS和SP含量明显上升;其中,200~300 μmol·L-1褪黑素处理效果最为显著,喷施外源褪黑素可以通过诱导草地早熟禾幼苗渗透调节物质的积累,维持渗透势平衡,从而提高草地早熟禾抵抗低温的能力。
植物能够通过光合作用合成有机物,是植物生长发育的基础。叶绿素是植物光合作用中重要的物质之一,也是反映叶片受损程度的一个直接指标[34]。低温胁迫会造成叶绿体和线粒体电子传递系统紊乱,是导致过量ROS产生的主要原因之一,也是破坏光合膜和其他细胞膜的主要因素之一,从而引发胁迫破坏。低温胁迫会破坏植物叶片细胞中叶绿体的结构,导致叶绿素发生降解,影响光能的吸收、转化和分配,阻碍光合电子传递和破坏光合机制,抑制碳同化作用,最终导致植物叶片的光合能力下降[35]。杨文飞等[36]研究发现,低温处理下水稻幼苗叶片中叶绿素含量下降,且随着低温时间的延长,呈逐渐下降的趋势,与叶绿素的合成和降解有关。本试验结果也表明,低温下草地早熟禾幼苗叶片中叶绿素的含量显著低于常温对照。练华山等[27]研究发现,壳聚糖可以显著提高丝瓜幼苗叶片中叶绿素的含量,其原因可能是壳聚糖可有效抑制低温下叶绿体类囊体片层结构紊乱、被膜内陷或破裂,促进叶绿体形成和叶绿素合成。此外,甜菜碱还能维持逆境条件下植物叶绿体体积,增加叶绿素a/b的值,降低类胡萝卜素和叶绿素的比值,保持Ca2+-ATPase活性和希尔反应活力,以维持植物的光合能力[39]。已有的研究表明,褪黑素可以通过上调冷响应途径的关键转录因子CBFs/DREBs和低温应答基因COR15a的表达水平,诱导转录激活因子CAMTA1,ZAT10和ZAT12的表达,从而提高拟南芥(Arabidopsis thaliana)的冷冻耐受性[38]。本试验中,叶面喷施壳聚糖、甜菜碱和褪黑素三种诱导剂能显著减缓低温下叶绿素降低的趋势,表明壳聚糖、甜菜碱和褪黑素可以通过增强叶绿素合成相关基因的表达或抑制低温下叶绿体结构的紊乱,维持草地早熟禾叶片内叶绿素含量的稳定,增强草地早熟禾对低温的抗性。
在正常环境条件下,植物体内抗氧化系统足以保护植物体免受活性氧的伤害,而低温胁迫下,由于植物细胞膜脂过氧化,并产生大量的ROS,使植物细胞膜受到伤害;但抗氧化酶活性也相应升高,来增强植物的抗氧化能力,减缓低温对植物细胞造成的伤害[39]。SOD具有催化歧化反应生成O和HO的作用,因此,被认为是植物逆境抗性强弱的重要指标[40]。植物的低温抗性强弱与植物体内是否能够维持较高的SOD活性水平有关[41]。POD和CAT是植物体内常见的氧化还原酶,具有分解植物体内HO防止细胞受损的作用,并与SOD一起在减少生物氧自由基浓度上共同作用,与植物低温损伤和耐寒性密切相关[42]。徐婷等[43]研究发现,低温胁迫下外源甜菜碱处理的玉米幼苗SOD、POD、CAT活性显著提高,能有效缓解低温对玉米幼苗的危害,其原因可能是甜菜碱能与膜外蛋白质的羧基阴离子相互作用,使低温下细胞膜的结构不被破坏[44]。张嘉雯等[45]研究表明,低温胁迫下,随着胁迫时间的延长,烟草(Nicotiana tabacum)幼苗的SOD和CAT活性先降低后升高,而POD活性呈持续降低的趋势,其可能是低温胁迫初期ROS大量积累,消耗了更多的抗氧化酶,褪黑素处理后这三种酶的活性均上升。低温胁迫下草地早熟禾SOD活性显著升高,POD和CAT的活性上升幅度较小,而叶面喷施壳聚糖、甜菜碱和褪黑素处理后这三种酶的活性显著升高,说明壳聚糖、甜菜碱和褪黑素能够提高抗氧化酶的活性,在低温胁迫时增强草地早熟禾幼苗清除ROS的能力,提高草地早熟禾幼苗的低温抗性。
4 结论
喷施三种诱导剂均能有效缓解低温对草地早熟禾幼苗造成的伤害,但三种诱导剂对草地早熟禾低温抗性的增强效果不同,抗低温效果由强到弱依次为褪黑素、甜菜碱、壳聚糖,且同种诱导剂不同浓度之间存在浓度效应。通过抗低温综合评价值相关性分析表明,300 μmol·L-1褪黑素喷施处理对提高草地早熟禾幼苗低温抗性的作用效果最强。
参考文献
[1]任继周,张自和. 草地与人类文明[J]. 草原与草坪,2000(1):5-9
[2]方强恩,孙英,白小明,等. 甘肃早熟禾属野生植物资源分布研究[J]. 中国草地学报,2010,32(6):39-45
[3]KONG R S,HENRY H A L.Does cross-acclimation between drought and freezing stress persist over ecologically relevant time spans? A test using the grass Poa pratensis[J]. Plant Biol (Stuttg),2018,20(2):280-287
[4]董文科,马祥,张玉娟,等. 低温胁迫对不同早熟禾品种糖酵解代谢及其相关基因表达的影响[J]. 草地学报,2019,27(6):1503-1510
[5]滕泽,张玉霞,陈卫东,等. 壳聚糖对苜蓿抗寒性及抗寒保护物质含量的影响[J]. 中国农业科技导报,2023,25(2):192-198
[6]谭毛,段亚玲,周梦春,等. 壳聚糖的分析方法及其在农业领域中的应用进展[J]. 农药,2021,60(1):6-10
[7]辛慧慧. 棉花对低温胁迫的生理响应及调控研究[D]. 石河子:石河子大学,2015:1-36
[8]JAN M,SHINWARI K I,HAFEEZ M,et al. Consequences of short term low temperature stress on physiological and biochemical aspects of rice (Oryza sativa)[J]. Scientia Agriculture, 2015,10(1):1-14
[9]姜秀梅. 外源物质处理对低温胁迫下辣椒种子萌发及幼苗抗冷性的影响[D]. 乌鲁木齐:新疆农业大学,2014:1-59
[10]DUAN W J,MENG Y J,JIANG D,et al. Effects of exogenous melatonin on the morphology and antioxidant enzyme activities of cotton seedlings under salt stress[J]. Chinese Journal of Eco-Agriculture,2022,30(1):92-104
[11]刘新宇,褚敏,张苗苗,等. 外源褪黑素对玉米幼苗低温冷害响应的影响[EB/OL]. http://kns.cnki.net/kcms/detail/46.1068.S.20230323.1054.005.html,2023-03-24/2024-06-13
[12]QINGHAI G,YAKUN W,XIAOMIN L U,et al. Effects of exogenous melatonin on growth and antioxidant system of leaves in cucumber seedlings under low temperature and weak light stress[J]. Acta Botanica Boreali-Occidentalia Sinica,2014,34(8):1608-1613
[13]ALLARD,FRANCE,HOUDE,et al. Betaine improves freezing tolerance in wheat[J]. Plant & Cell Physiology,1998,39(11):1194-1202
[14]刘思露. 外源甜菜碱对匍匐翦股颖抗旱性的影响[D]. 北京:北京林业大学,2015:1-12
[15]NAM,JUN,KANG,et al. Effects of foliar application of glycine betaine on the growth and contents of osmolyte in tomato seedling[J]. Journal of Bio-Environment Control,2006,15(4):390-395
[16]舒必超,杨勇,刘雪勇,等.低温胁迫对狗牙根生理及基因表达的影响[J].草业学报,2018,27(11):106-119
[17]KAMATA T,UEMURA M. Solute accumulation in heat seedlings during cold acclimation:contribution to increased freezing tolerance[J]. Cryo Letters,2004,25(5):311-322
[18]刘陈晨,李旭,任士伟,等. 低温胁迫下外源甜菜碱对水稻秧苗期生长的影响[J]. 江苏农业科学,2019,47(11):100-102
[19]魏臻武,范占炼,王槐三. 不同类型草坪草的抗寒锻炼[J]. 草业科学,1997(3):61-66
[20]张志良,翟伟箐. 植物生理学实验指导[M]. 第3版. 北京:高等教育出版社,2003:151-153
[21]李玲. 植物生理学模块实验指导[M]. 北京:科学出版社,2009:96-97
[22]BATES L S,WALDREN R P,TEARE I D. Rapid determination of free proline for water stress studies[J]. Plant and Soil,1973,39(1):205-207
[23]YEMM E W,WILLIS A J. The estimation of carbohydrates in plant extracts by anthrone[J]. Biochemical Journal,1954,57(3):508-514
[24]李合生. 植物生理生化实验原理和技术[M]. 北京:高等教育出版社,2000:134-261
[25]贺佳圆. 9个野生早熟禾种质材料抗寒性研究[D]. 兰州:甘肃农业大学,2012:11-24
[26]孙景宽,张文辉,张洁明,等. 种子萌发期4种植物对干旱胁迫的响应及其抗旱性评价研究[J]. 西北植物学报,2006,26(9):1811-1818
[27]练华山,周艾玲,梁乐,等. 叶面喷施壳聚糖缓解低温处理对丝瓜幼苗的影响[J]. 云南农业大学学报(自然科学),2023,38(2):298-305
[28]尉欣荣,张智伟,周雨,等. 褪黑素对低温和干旱胁迫下多年生黑麦草幼苗生长和抗氧化系统的调节作用[J]. 草地学报,2020,28(5):1337-1345
[29]HEIDARVAND L,AMIRI R M. What happens in plant molecular responses to cold stress?[J]. Acta Physiologiae Plantarum,2010,32(3):419-431
[30]李贺. 褪黑素对大豆苗期低温胁迫抗性的调控作用[D]. 大庆:黑龙江八一农垦大学,2021:55-57
[31]朱云林,顾大路,王伟中,等. 壳聚糖对水稻幼苗抗冷性的影响[J]. 江苏农业科学,2017,45(8):66-68
[32]孙巧峰,于贤昌,高俊杰,等. 羧甲基壳聚糖对黄瓜幼苗抗冷性的影响[J]. 中国农业科学,2004(11):1660-1665
[33]白如意,宋希梅,沈健,等. 叶面喷施褪黑素对低温胁迫下南瓜幼苗生长和生理特性的影响[J]. 西北植物学报,2023,43(5):805-813
[34]廖浩钦,谢文刚. 牧草应对低温胁迫机制研究进展[J]. 中国草地学报,2023,45(12):99-111
[35]郝辉芳,范月仙,李生泉. 低温锻炼对冷胁迫下棉苗叶片细胞超微结构的影响[J]. 棉花学报,2017,29(3):268-273
[36]杨文飞,杜永林,顾大路,等. 4种调节物质对水稻耐低温能力的影响[J]. 江苏农业学报,2017,33(4):739-746
[37]于郑娇,李书昆,褚鹏飞,等.外源褪黑素对低温胁迫下番茄幼苗光合荧光及生化特性的影响[J]. 山东农业科学,2023,55(8):48-55
[38]BAJWA V S,SHUKLA M R,SHERIF S M,et al. Role of melatonin in alleviating cold stress in Arabidopsis Thaliana[J]. Journal of Pineal Research,2014,56(3):238-245
[39]董文科,马祥,周学文,等. 外源甜菜碱对低温胁迫下紫花苜蓿幼苗生理特性的影响[J]. 草地学报,2019,27(1):130-140
[40]陈鸿鹏,谭晓风. 超氧化物歧化酶(SOD)研究综述[J]. 经济林研究,2007,25(1):59-65
[41]英潘. 干旱胁迫对不同甘蔗品种宿根蔗抗旱生态生理特性及产量品质的影响[D]. 南宁:广西大学,2014:2-5
[42]周瑞莲,张承烈,金巨和. 水分胁迫下紫花苜蓿叶片含水量、质膜透性SOD、CAT活性变化与抗旱性关系研究[J]. 中国草地,1991(2):20-24
[43]徐婷,王俊强,韩业辉,等. 外源甜菜碱浸种对低温胁迫下玉米种子萌发及幼苗生长的影响[J]. 黑龙江农业科学,2023(8):8-12
[44]裴海荣,郭宗端,李伟,等. 外源物质提高作物抗寒性研究进展[J]. 湖北农业科学,2017,56(3):409-412
[45]张嘉雯,卢绍浩,赵喆,等. 外源褪黑素对低温胁迫下烟草幼苗生理指标的影响[J]. 中国农业科技导报,2020,22(9):78-86
(责任编辑 彭露茜)
