
养分及水分对少花蒺藜草光合特性及抗氧化酶活性的影响
2024-11-05 00:00陈佳宁王舒琦罗亚勇周立业赵鑫宇夏咪王鹤松包金连
草地学报 2024年10期








收稿日期:2024-02-01;修回日期:2024-04-15
基金项目:内蒙古自治区重点研发和成果转化计划项目(2022YFDZ0028);内蒙古自然基金联合项目(2019LH03022);内蒙古自治区科技重大专项(2021ZD0041-005);国家自然科学基金资助项目(42142027);科尔沁沙地生态农业国家民委重点实验室开放基金(MDK2023097)资助
作者简介:陈佳宁(1998-),男,蒙古族,内蒙古通辽人,硕士研究生,主要从事生态修复与环境治理,E-mail:1120138440@qq.com;*通信作者Author for correspondence,E-mail:toni2002@126.com
摘要:为探究入侵植物少花蒺藜草(Cenchrus pauciflorus)在异质养分及干旱复水条件下的光合特性及生理变化,本研究以少花蒺藜草为试验对象,设置对照(CK)、低养分、中养分和高养分4个养分水平,在施肥的基础上设置2种水分处理(干旱后浇水与正常浇水),培育40 d后,开始3个周期的干旱复水试验(每个周期为7 d)。结果表明:在周期Ⅰ时,干旱组中养分的蒸腾速率(Transpiration rate,Tr)达到最大值2.950 mmol·m-2·s-1;在每次复水后,气孔导度(Stomatal conductance,Gs),胞间CO浓度(Intercellular carbon dioxide concentration,Ci),Tr都有明显的回升,瞬时水分利用效率(Instantaneous water use efficiency,WUE)在第3次干旱复水周期时,水平明显提高;少花蒺藜草高养分干旱组超氧化物歧化酶(Superoxide dismutase,SOD)活性达到最大值384.17 U·g-1FW。综合分析,在中养分水平,少花蒺藜草对水分胁迫的抗性更强,经历干旱复水后仍具有较强的干旱适应能力、较高的水分利用效率和复水恢复能力。
关键词:少花蒺藜草;净光合速率;瞬时水分利用率;干旱复水
中图分类号:S451 文献标识码:A 文章编号:1007-0435(2024)10-3091-12
Effects of Nutrients and Water on Photosynthetic Characteristics and Antioxidant Enzyme Activities of Cenchrus pauciflorus
CHEN Jia-ning1,2, WANG Shu-qi1, LUO Ya-yong2, ZHOU Li-ye1*, ZHAO Xin-yu2,3, XIA Mi2, WANG He-song1,2, BAO Jin-lian4
(1. College of Agriculture,Inner Mongolia Minzu University, Tongliao, Inner Mongolia 028000, China; 2. Northwest Institute of Eco-Environment and Resources, Chinese Academy of Sciences, Lanzhou,Gansu Province 730000, China; 3. Inner Mongolia Agricultural University,
Hohhot,Inner Mongolia 010018, China;4. Tongliao Market Inspection Center, Tongliao, Inner Mongolia 028000, China)
Abstract:In order to investigate the photosynthetic characteristics and physiological changes of invasive plant Cenchrus pauciflorus under heterogeneous nutrient and drought and rehydration conditions,this study took Cenchrus pauciflorus as the test object,and set four nutrient levels including CK,low nutrient,medium nutrient and high nutrient. On the basis of fertilization,two kinds of water treatments (post-drought watering and normal watering) were set up,and after 40-day cultivation,three cycles of drought rehydration experiment (each cycle was 7 days) were started. The results showed as follows:In cycle Ⅰ,the Transpiration rate (Tr) of nutrients in the drought group reached the maximum of 2.950 mmol·m-2·s-1; After each rehydration,stomatal conductance (Gs),intercellular carbon dioxide concentration (Ci) and Tr all showed significant recovery. Instantaneous water use efficiency (WUE) increased significantly in the 3rd drought rehydration cycle. Superoxide dismutase (SOD) activity reached the maximum value of 384.17 U·g-1FW in the high-nutrient drought group. Comprehensive analysis showed that Cenchrus pauciflorus had stronger resistance to water stress at the medium nutrient level,and still had stronger drought adaptation ability,higher water use efficiency and water recovery ability after drought rehydration.
Key words:Cenchrus pauciflorus;Net photosynthetic rate;Water use efficiency;Drought rehydration
养分和水分是植物生长、发育和维持所需的重要资源,对植物的生长发育有重要的影响[1]。土壤养分直接影响植物体内养分含量,并通过影响养分在叶片中的分配来调节植物的光合及生理作用[2]。同时,水分亏缺是影响植物光合和生理诸多环境因素中最主要的[3]。植物自身对干旱胁迫所导致的伤害有补给作用,在生理生态方面均能得到不同程度的恢复,展现出一定的生理补偿效应,是植物在抵御外界短期干旱胁迫的一种体内调节机制[4]。有研究表明,植物对干旱复水的响应方式是在干旱胁迫解除后通过快速生长或提高蒸腾速率(Transpiration rate,Tr)和净光合速率(Net photosynthetic rate,Pn),增加叶绿素(Chlorophyl,chl)含量,提高逆境时的抗氧化酶活性,增强其抗氧化能力等来补偿部分或全部因水分亏缺对自身造成的伤害[5-6]。
V养分添加一定程度上可以改善干旱胁迫对植株的危害。研究表明,养分添加可以增加在水分亏缺下植物叶肉细胞的光合活力,进而增强单叶的净光合速率[7]。养分添加优化了毛竹(Phyllostachys edulis)自身有限资源的分配,促进毛竹根部形成,减少了干旱对毛竹发育的抑制作用[8]。在不同水分胁迫条件下紫苏(P. frutescens)叶片Chl含量随着养分含量的提高而增加[9]。
由此可见,养分及水分对植物生长发育,光合及生理都有着十分复杂的影响。禾本科少花蒺藜草(Cenchrus pauciflorus)属一年生草本,现如今已经成为了科尔沁沙地最为严重的入侵杂草之一[10-11]。在水分亏缺的情况下,少花蒺藜草通过调整生物构件如降低穗生物量,以及减少分株高度等的生存方式来适应当地自然环境[12]。对于在半干旱沙地生态系统的入侵植物少花蒺藜草,它是如何适应异质养分和干旱环境的光合及生理响应机制尚不了解。因此,本研究以入侵植物少花蒺藜草为研究对象,在设置不同养分和水分的基础上对少花蒺藜草进行干旱复水处理,分析它在不同养分与水分作用下的光合及生理响应差异,以期明确少花蒺藜草异质养分条件下独特的光合和生理适应机制,为减少和控制少花蒺藜草的扩散和蔓延提供科学依据。
1 材料与方法
1.1 供试材料及试验地概况
本研究依托于中国科学院西北生态环境资源研究院奈曼沙漠化研究站开展,研究区位于科尔沁沙地中南部内蒙古自治区通辽市奈曼旗(42°55′N~42°57′N,120°40′E~120°43′E;海拔约为360 m)境内。科尔沁沙地属于温带大陆性半干旱季风气候,春季干旱而多风,夏季炎热、雨热同期,冬季寒冷。该区域年平均降雨量约360 mm,年均蒸发量约1972.8 mm,其中降水量的70%~80%集中在6—8月,年平均气温6.4℃,无霜期约150 d,年平均风速3.5 m·s-1,全年≥10℃的有效积温在3000℃以上。该区域的土壤类型以栗钙土和风沙土为主,而地貌类型则表现出流动沙丘、半固定沙丘、固定沙丘、平缓草地交错分布的特征[13-14]。供试少花蒺藜草刺苞种子于2022年5月采集于奈曼沙漠化研究站附近孟和嘎查道路两旁(42°57′N,120°40′E;海拔约为350 m)。
1.2 试验设计
试验以入侵植物少花蒺藜草为研究对象,在种植前设置了4个不同的施肥处理:一是不施加任何肥料的纯沙(CK);二是施加2.1 g磷酸二铵(N∶P∶K=18∶46∶0)与0.4 g复合肥(N∶P∶K=14∶16∶15)(低养分);三是施加7.8 g磷酸二铵与1.6 g复合肥(中养分);四是施加28.1 g磷酸二铵与5.6 g复合肥(高养分),均与纯沙混合后放置在花盆(上内径18.5 cm,下内径16 cm,高21 cm)内,填装至距离盆口5 cm处,养分水平相当于未发生沙漠化土地的营养水平[15]。将籽粒饱满、大小一致的刺苞置于去离子水中浸泡24 h,浸泡后的种子用浓度为2%的次氯酸钠溶液灭菌30 min,再用蒸馏水洗涤5—6遍,吸干水分,种在预先准备好的花盆中,每盆播种20粒少花蒺藜草种子,覆土2~3 cm后压实浇水。
此外,为了解施肥后少花蒺藜草对土壤干旱及旱后复水的响应,本研究设置了2种不同水分处理:一是保持土壤水分较好(接近田间持水量的对照);二是在自然条件下干旱,极端干旱后恢复土壤水分。5月27开始试验,经过40 d的培养后开始干旱复水试验(每个干旱复水周期为7 d),干旱组在每个干旱周期结束时进行复水(田间持水量),浇水组在干旱周期内每间隔3日浇一次水(50%田间持水量)。干旱复水过程进行了3个周期,在每个干旱复水周期的第3 d及第6 d测定叶片光合指标,在第4 d及第7 d时,从CK、中养分及高养分处理中已做好标记的植株上采集约10 g完全展开的绿色叶片,放置于液氮罐中保存,用于测定生理指标。
养分及水分处理共4(土壤养分)×2(土壤水分)=8个处理,每个处理8个重复,共计64盆,所有花盆均放置在智能温室大棚内,温室内空气温度23~37℃,湿度26%~44%,光照强度0~27 412 Lux,CO浓度390~552 mg·L-1,平均风速0.7 m·s-1左右。除了避免降水进入盆中,尽量保持与自然环境条件一致。
1.3 指标测定及其方法
用便携式光合分析系统(LI-6400,LI-COR Inc,Lincoln,NE,USA)测定植物的光合特性指标,在上午8:00—11:00进行。测定时为了保持在整个干旱复水期间光强和温度条件一致,采用人工光源(Li-6400-02B,Li-Cor,USA)提供1500 μmol·m-2·s-1光合有效辐射,该光强值达到各样点植物的光饱和点以上,且对植物的生理过程不产生抑制作用[16]。测定时使用开放式气路,空气流速为0.5 dm3·min-1,待数值稳定2 min后读数。选取植物上部充分展开的叶片进行活体测定,测定不同植株上的3~5片叶子,每片叶子读取仪器稳定后的5个数据,计算平均值。根据Pn与Tr的比值计算植物叶片的瞬时水分利用效率(Instantaneous water use efficiency,WUE)[17]。叶绿素含量(Chl)采用95%乙醇浸提比色法、过氧化氢酶活性(Catalase,CAT)采用紫外吸收法测定、过氧化物酶活性(POD)采用愈创木酚比色法、超氧化物歧化酶活性(SOD)采用NBT光化还原法测定[18]。
1.4 数据处理
所有数据均使用Microsoft Office Excel 2013进行图表制作,使用SPSS 26.0进行单因素方差分析(One-way ANOVA)和多因素方差分析(Multi-way ANOVA),显著度水平为P<0.05。
2 结果与分析
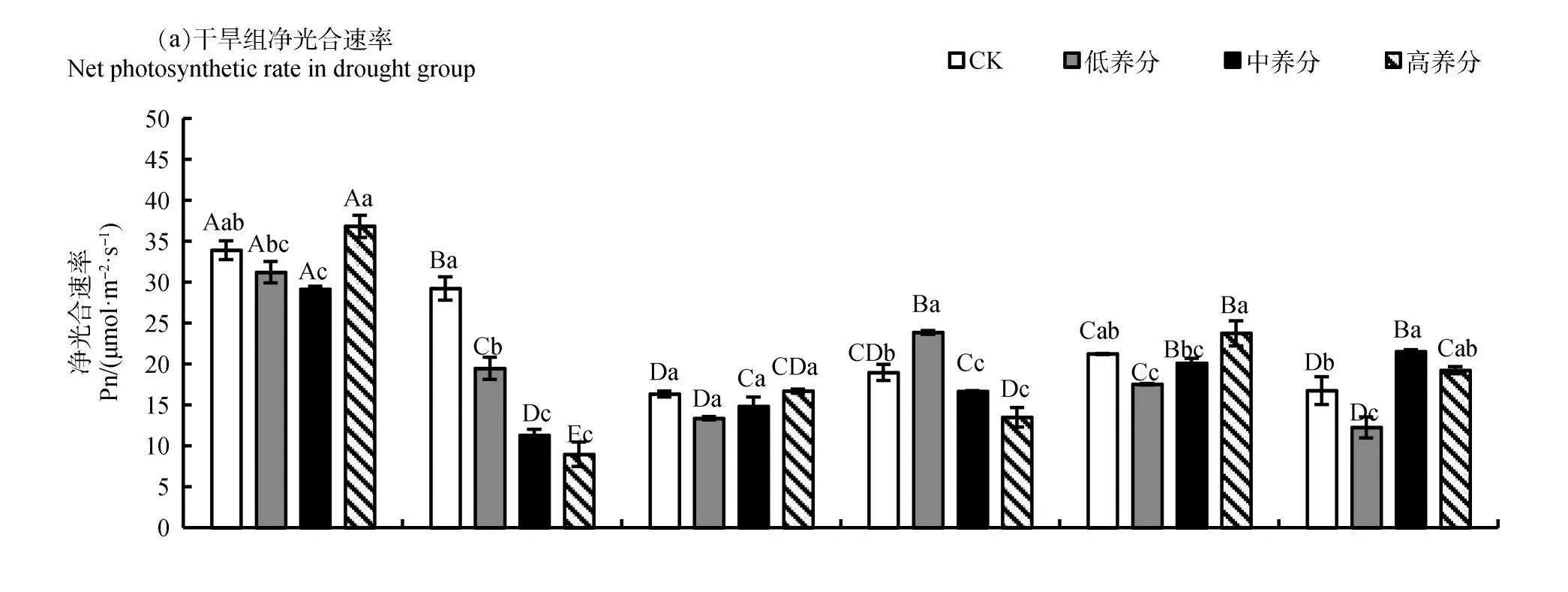
2.1 养分及水分对少花蒺藜草叶片净光合速率的影响
由图1所示,在周期Ⅰ时,干旱组第3 dCK、低养分、中养分、高养分的少花蒺藜草Pn均显著高于其他时期处理(P<0.05),浇水组第3 d低养分处理的Pn均显著高于其他时期低养分处理(P<0.05),两组处理第3 d时,CK、低养分、中养分、高养分的少花蒺藜草Pn均达到最大值,第6 d时,干旱组中养分、高养分的Pn达到最小值;在周期Ⅱ时,干旱组第6 d低养分处理的Pn显著高于同一时间其他处理(P<0.05),浇水组第3 d中养分处理的Pn显著高于同1 d的低养分与高养分处理(P<0.05);在周期Ⅲ时,干旱组第3 d高养分的Pn显著高于低养分、中养分(P<0.05),浇水组高养分的少花蒺藜草Pn呈现为先下降后上升再下降的变化趋势,且CK、低养分、中养分、高养分处理的Pn在第6 d时达到最小值。
2.2 养分及水分对少花蒺藜草叶片气孔导度的影响
由图2所示,在整个干旱复水周期内,干旱组低养分与中养分的少花蒺藜草Gs呈先下降后上升再下降的趋势;在周期Ⅰ时,干旱组第3 d低养分、中养分、高养分的少花蒺藜草Gs均达到最大值,分别为0.172 mol·m-2·s-1,0.155 mol·m-2·s-1,0.169 mol·m-2·s-1,干旱组第6 d的低养分与其他处理差异显著(P<0.05),浇水组第3 d的低养分处理显著高于其他时期低养分处理(P<0.05);第6 d的低养分处理与其他处理差异显著(P<0.05),且中养分、高养分的Gs达到最小值,分别为0.049 mol·m-2·s-1,0.038 mol·m-2·s-1;在周期Ⅱ时,干旱组第6 d高养分处理的Gs显著低于同一天的其他处理(P<0.05),浇水组,中养分处理的Gs在第6 d达到最小值;在周期Ⅲ时,浇水组低养分、高养分在周期Ⅲ的第6 d达到最小值。
2.3 养分及水分对少花蒺藜草叶片胞间CO浓度的影响
在周期Ⅰ时,干旱组第3 d中养分处理的少花蒺藜草Ci显著高于同一天其他处理(P<0.05),浇水组CK、低养分、中养分、高养分处理Ci达到最小值,分别为20.747 μmol·m-2·s-1,12.192 μmol·m-2·s-1,26.753 μmol·m-2·s-1,22.875 μmol·m-2·s-1,第6 d,干旱组低养分处理的Ci显著高于同一天其他处理(P<0.05);在周期Ⅱ时,干旱组第6 d低养分的少花蒺藜草Ci达到最小值60.041 μmol·m-2·s-1,浇水组第3 d高养分处理的少花蒺藜草Ci显著高于同一d其他处理(P<0.05);在周期Ⅲ时,干旱组第3 d的低养分显著高于其他时期低养分处理(P<0.05),且CK、低养分处理的少花蒺藜草Ci达到最大值。浇水组第3和第6 d的中养分均显著高于其他时期中养分处理(P<0.05)(图3)。
2.4 养分及水分对少花蒺藜草叶片蒸腾速率的影响
由图4所示,在周期Ⅰ时,干旱组第3 d的高养分显著高于其他时期高养分处理(P<0.05),且低养分、中养分、高养分的少花蒺藜草Tr达到最大值分别为3.380 mmol·m-2·s-1,2.950 mmol·m-2·s-1,3.431 mmol·m-2·s-1;在周期Ⅱ时,干旱组第3 d,低养分、中养分的少花蒺藜草Tr达到最小值,浇水组第3 d中养分处理的Tr显著高于其他处理(P<0.05);在周期Ⅲ时,浇水组CK、低养分、中养分、高养分处理的Tr在第3 d均达到最小值;在整个干旱复水周期内,干旱组低养分、中养分、高养分的少花蒺藜草Tr呈先下降后上升再下降的趋势。
2.5 养分及水分对少花蒺藜草叶片瞬时水分利用率的影响
在周期Ⅰ时,干旱组第6 d,CK、低养分、高养分的少花蒺藜草WUE达到最小值,分别为7.124 μmol·mmol-1,6.500 μmol·mmol-1,7.853 μmol·mmol-1,浇水组第3 d与周期Ⅲ第6 d内低养分、高养分条件下的WUE显著高于其他时期不同处理(P<0.05),浇水组CK、中养分在第3 d达到最大值;在周期Ⅱ时,干旱组第3 d与周期Ⅲ第6 d内中养分条件下的WUE显著高于其他时期不同处理(P<0.05),干旱组第3 d,CK、低养分、中养分的少花蒺藜草的WUE达到最大值,分别为11.851 μmol·mmol-1,10.716 μmol·mmol-1,13.058 μmol·mmol-1,浇水组CK、中养分、高养分在第6 d达到最小值,分别为4.144 μmol·mmol-1,7.482 μmol·mmol-1,7.431 μmol·mmol-1;在周期Ⅲ时,干旱组高养分少花蒺藜草的WUE在第6 d达到最大值13.043 μmol·mmol-1,浇水组低养分、高养分在周期Ⅲ的第6 d达到最大值;在整个干旱复水周期,干旱组低养分、中养分、高养分的少花蒺藜草WUE呈W型的变化趋势,浇水组CK、低养分、中养分、高养分均表现为N型的变化趋势(图5)。
2.6 养分及水分对少花蒺藜草叶绿素含量的影响
如图6所示,在干旱复水周期Ⅰ内的第4 d时,同一养分和水分处理的少花蒺藜草Chl含量均分别存在显著差异,且中养分、高养分浇水组的Chl含量均显著高于干旱组(P<0.05);在同一水分条件下,干旱组中养分的Chl含量与CK、高养分的Chl含量均存在显著差异(P<0.05),浇水组中养分的Chl含量均显著高于CK和高养分(P<0.05),浇水组中养分的Chl含量达到最高为3295.88 mg·g-1。在周期Ⅰ的第7 d时,同一养分条件下,高养分的少花蒺藜草Chl含量存在显著差异,且干旱组显著高于浇水组(P<0.05);在同一水分条件下,干旱组中养分、高养分的Chl含量均显著高于CK(P<0.05),浇水组中养分的Chl含量显著高于CK、高养分(P<0.05)。
在干旱复水周期Ⅲ内的第4 d时,同一养分条件下,CK与高养分的少花蒺藜草Chl含量存在显著差异,且浇水组显著高于干旱组(P<0.05);在同一水分条件下,干旱组高养分的Chl含量与CK存在显著差异(P<0.05),浇水组高养分的Chl含量均显著高于CK和中养分(P<0.05)。在周期Ⅲ的第7 d时,同一养分条件下,干旱组少花蒺藜草Chl含量均显著高于浇水组(P<0.05);在同一水分条件下,干旱组与浇水组中养分、高养分的Chl含量均显著高于CK(P<0.05)。
2.7 养分及水分对少花蒺藜草抗氧化物酶活性的影响
由图7可知,在干旱复水周期Ⅰ内的第4 d时,同一养分条件下,中养分、高养分干旱组少花蒺藜草的CAT活性均显著高于浇水组(P<0.05)。中养分浇水组少花蒺藜草的POD活性均显著高于干旱组(P<0.05)。少花蒺藜草SOD活性均存在显著差异,且干旱组显著高于浇水组(P<0.05);在同一水分条件下,高养分干旱组少花蒺藜草的CAT活性与CK相比存在显著差异(P<0.05);在同一水分条件下,干旱组CK、中养分、高养分少花蒺藜草的CAT活性均存在显著差异(P<0.05),且高养分的CAT活性达到最大值为15.56 U·g-1FW。干旱组高养分少花蒺藜草的CAT活性与CK相比存在显著差异(P<0.05)。干旱组中养分、高养分的SOD活性显著高于CK(P<0.05),浇水组均存在显著差异(P<0.05)。在周期Ⅰ的第7 d时,同一养分条件下,中养分、高养分的少花蒺藜草CAT活性存在显著差异,且干旱组显著高于浇水组(P<0.05)。中养分、高养分的少花蒺藜草POD活性存在显著差异,且浇水组显著高于干旱组(P<0.05)。高养分干旱组的SOD活性显著高于浇水组(P<0.05);在同一水分条件下,干旱组CK、中养分、高养分的CAT活性差异显著,且中养分、高养分均显著高于CK(P<0.05),浇水组CK、中养分、高养分的CAT活性差异显著,且高养分显著高于CK、中养分(P<0.05),其CAT活性达到最大值22.89 U·g-1FW。干旱组与浇水组中养分的POD活性显著高于CK、高养分(P<0.05)。干旱组CK、中养分、高养分的SOD活性差异显著(P<0.05),浇水组高养分的SOD活性显著高于CK、中养分(P<0.05),其SOD活性达到最大值343.53 U·g-1FW。
在干旱复水周期Ⅲ内的第4 d时,同一养分条件下,少花蒺藜草的CAT活性均存在显著差异,且中养分干旱组显著高于浇水组,高养分浇水组显著高于干旱组(P<0.05)。少花蒺藜草的POD活性均存在显著差异,且干旱组显著高于浇水组(P<0.05)。少花蒺藜草的SOD活性均存在显著差异,且干旱组显著高于浇水组(P<0.05);在同一水分条件下,干旱组中养分、高养分的CAT活性均显著高于CK(P<0.05),浇水组CK、中养分、高养分的CAT活性差异显著(P<0.05)。干旱组中养分的POD活性均显著高于CK、高养分(P<0.05)。干旱组中养分、高养分的SOD活性均显著高于CK(P<0.05),浇水组CK、中养分、高养分的SOD活性均存在显著(差异P<0.05)。在周期Ⅲ的第7 d时,同一养分条件下,中养分干旱组少花蒺藜草CAT活性显著高于浇水组(P<0.05)且达到最大值12.00 U·g-1FW。少花蒺藜草POD活性均存在显著差异(P<0.05)。高养分干旱组少花蒺藜草SOD活性显著高于浇水组且SOD活性达到最大值384.17 U·g-1FW(P<0.05);在同一水分条件下,干旱组中养分、高养分的CAT活性均显著高于CK(P<0.05),浇水组高养分的CAT活性均显著高于CK、中养分(P<0.05)。干旱组与浇水组中养分的POD活性均显著高于CK、高养分(P<0.05)。干旱组与浇水组高养分的CAT活性显著高于CK、中养分(P<0.05)。
3 讨论
3.1 养分及水分对少花蒺藜草光合特性及水分利用效率的影响
光合作用是植物最为重要的生命活动,也是植株生长发育的基础,而养分和水分是影响光合作用的关键因子[19-20]。养分差异和干旱胁迫都能直接影响植物的光合作用,进而影响植物的生长发育,其作用强度取决于养分含量多少,干旱程度和持续时间[21]。植物的干旱适应性可以分为耐旱能力和复水恢复能力两个方面,复水恢复能力既是植物适应干旱环境的主要内容,又是植物应对干旱胁迫的重要机理[22]。水分亏缺时,植物主动降低Gs,减少水分流失,进而降低Pn,减少对水分的需求,以增强对干旱胁迫的适应。引起植物光合速率降低的因素,除了气孔限制以外,还有非气孔限制,Gs和Ci是判断光合速率下降原因的主要依据[23-24]。当Pn、Gs和Ci变化趋势一致时,光合速率下降主要由气孔限制引起,而当它们变化不一致时,光合速率降低主要由叶肉细胞活性所决定,即非气孔影响[25]。本试验结果表明,在经历频繁干旱复水周期后,两组处理中,低养分,中养分,高养分的少花蒺藜草Pn、Gs和Ci变化趋势基本趋于一致,呈现倒S的发展趋势,但干旱处理组较浇水处理组,后者所变化的趋势更为平缓,可能的原因是,少花蒺藜草在经历干旱复水前期,正常的水分供应让少花蒺藜草的叶肉细胞储存了足够多的水分,从而降低了少花蒺藜草在经历频繁干旱复水周期过程中受到的损伤,同时,少花蒺藜草通过主动降低Gs,使得Pn下降,减少水分流失,吸收土壤养分等方式,弥补了一定程度叶片受到的损伤。蒸腾是植株体内的水分以气体状态向外消耗的过程,是植株水分代谢至关重要的环节之一[26],蒸腾速率反映了植株水分的代谢能力[27]。由图(4)可以看出,在反复干旱复水周期内,少花蒺藜草的Tr在每个干旱复水周期的3—6 d,Tr都有明显的下降,而在第6 d进行复水后,Tr又有明显的回升,可能的原因是少花蒺藜草在遭受干旱胁迫时,通过关闭气孔,降低叶片蒸腾速率,从而减少水分的消耗,在复水后,叶片得到水分补充,又开始正常的光合作用进行生长发育。少花蒺藜草在复水后,Gs,Ci,Tr都有明显的回升,一定程度上说明了少花蒺藜草拥有较好的干旱适应性特别是复水恢复能力,也体现了少花蒺藜草在经历频繁干旱复水时的光合生理适应策略。
水分利用效率(WUE)直接反映植物蒸散消耗单位质量水所产生的干物质量,可提供有关水分代谢功能的重要信息[28-29]。干旱胁迫下,植物首先关闭气孔以降低Tr,减少水分消耗,并通过限制气体交换影响Pn。WUE在一定程度上可以反映植物的耗水特征和对干旱的适应,提高WUE是植物应对干旱的主要响应机理之一[30]。本试验结果表明,在整个干旱复水周期内,浇水组中养分处理的少花蒺藜草Pn、WUE较干旱组低养分、高养分处理的Pn、WUE高,说明浇水组的中养分处理是少花蒺藜草能更好抵抗干旱胁迫的适宜施肥量,弥补了干旱胁迫对叶肉细胞造成的损伤。两组处理中,少花蒺藜草的WUE在整个干旱复水周期内,始终都保持在相对较高的水平,在经历第3次干旱复水周期时,少花蒺藜草的WUE水平显著提高,可能的原因是少花蒺藜草具有较高的水分利用效率,叶片保水能力强,一定程度上限制了叶片表面的水分蒸发,对水分输出节约,减弱了频繁干旱胁迫对光合作用的限制,提高了少花蒺藜草对干旱胁迫的抗性。
叶绿素含量的改变一定程度上能体现植物光合作用的变化规律,并作为植物对水分胁迫敏感性的指示性指标[31]。当植株遭到水分亏缺时,Chl含量首先表现为增加,随着胁迫程度的加深则表现为降低[32]。姜野[33]研究表明,随着养分含量的提高,两种土壤基质中Chl含量差异显著,且少花蒺藜草幼苗Chl含量呈现为显著提高。本试验结果表明,当少花蒺藜草经历干旱第1个周期时,在同一水分条件时,干旱组中养分与高养分的叶绿素含量明显升高,而浇水组的叶绿素有所降低,在中养分条件下,叶绿素含量显著高于其他两组养分处理。可能的原因是,中养分条件下,提高了少花蒺藜草的叶绿素含量,增强了其光合作用,加快了根系对土壤水分的吸收,在经历干旱胁迫时,叶绿素浓缩效应导致叶片含水量降低,少花蒺藜草叶片出现了萎蔫卷曲现象,正常施水时,叶片含水量相对较好,有效减缓了叶绿素的降解。经过复水恢复后,当少花蒺藜草干旱第3个周期时,第4 d,干旱组与浇水组的少花蒺藜草Chl含量相较于干旱第1个周期的Chl含量有明显升高,而到第7 d时,叶绿素含量骤然降低,可能的原因是,复水后,少花蒺藜草的特有的生理补偿效应影响了叶片的含水量,增强了少花蒺藜草的光合作用,当少花蒺藜草经历多次干旱复水周期后,水分亏缺可能使得叶片的叶绿体受到了不可逆的损伤,养分也不能得到相应的补充,使少花蒺藜草生长受到限制,衰减了光合作用,减少了Chl含量,从而影响到生物量的调配。
3.2 养分及水分对少花蒺藜草抗氧化酶活性的影响
干旱胁迫往往会诱发植物体内产生更多活性氧,引起膜脂过氧化,蛋白质和核酸的分解,从而产生氧化物质[34]。抗氧化物酶是植物体细胞内清除系统的主要保护酶,能够有效的控制高浓度氧的持续积累,提高自身的抗逆性[35]。大量研究发现,适量的养分及水分条件对植物生长有一定促进作用,而少量或过量则会产生抑制作用,这些促进或抑制通常由植物形态、生理性状所表现[36]。本试验结果表明,少花蒺藜草在经历第一个干旱复水周期的第4 d时,同一养分条件下,中养分、高养分干旱组少花蒺藜草的CAT、SOD活性均显著高于浇水组,当干旱胁迫到达第7 d时,CK、中养分、高养分条件下,干旱组与浇水组的抗氧化物酶活性都有明显的提高,且干旱组较浇水组提高得更多。随着养分含量的增加,少花蒺藜草在经历干旱胁迫时,抗氧化酶活性也随之增强,而SOD酶活性明显升高恰恰说明少花蒺藜草对活性氧的歧化能力增强了,将活性氧转化为HO,CAT和POD酶活性显著高于CK,说明CAT和POD有效清除了由SOD歧化活性氧后产生的HO,有效缓解了少花蒺藜草的膜脂过氧化,减轻了干旱对少花蒺藜草的损伤,有助于其在恢复土壤水分后尽快恢复生理活性。经过复水后,少花蒺藜草有明显的恢复特征,如卷曲的叶片开始舒展,耷拉下来的叶片开始挺立,因此,土壤水分对少花蒺藜草叶片中抗氧化酶活性的影响占有主导地位,干旱胁迫会导致抗氧化酶活性显著升高。当少花蒺藜草经历干旱周期的第4 d时,少花蒺藜草的CAT、SOD酶活性相比第一个周期时明显下降,可能干旱对少花蒺藜草细胞造成的损伤仍未完全解除。第7 d时,中养分条件下的少花蒺藜草CAT、SOD酶活性有所升高,可能少花蒺藜草需要保持较高的CAT和SOD活性,高效清除HO,从而更好地保护和修复干旱胁迫下受到的伤害。在两次干旱周期内,少花蒺藜草的POD活性始终保持在较高水平,且中养分条件下,相较其他两组处理,酶活性更高,可能是少花蒺藜草在适宜的养分补给条件下,通过维持较高的POD酶活性来抵御干旱环境,保证植株的正常生长发育。本研究中养分、水分及其两因素间的交互作用均对少花蒺藜草的抗氧化酶活性有一定的影响,但不同的酶对水肥响应情况不同,原因是当植物处于不同的水肥环境和时间点时,参与保护机制的主力酶不同,此消彼长。因此,在未来的研究中,对于少花蒺藜草在不同环境条件下抗氧化酶活性的变化规律,还需要从分子应答机制的角度,结合植物体内活性氧浓度的改变进行综合研究。
4 结论
少花蒺藜草在中养分条件下能更好的抵御水分胁迫。在整个干旱复水周期内,少花蒺藜草的WUE始终都保持在较高的水平,说明其具有较强的干旱适应能力以及较高的水分利用效率和复水恢复能力。在养分差异及干旱复水条件下少花蒺藜草通过增加叶绿素含量,提高净光合速率,提高过氧化物酶活性等方式,维持植株的正常生长发育。体现了少花蒺藜草在经历频繁干旱复水时的光合及生理适应策略,是少花蒺藜草能在科尔沁沙地干旱环境成功入侵的原因之一。
参考文献
[1]杜忠毓,安慧,王波,等. 养分添加和降水变化对荒漠草原植物群落物种多样性和生物量的影响[J]. 草地学报,2020,28(4):1100-1110
[2]陆姣云,杨惠敏,田宏,等. 水分对不同生育时期紫花苜蓿茎叶碳、氮、磷含量及化学计量特征的影响[J]. 中国草地学报,2021,43(6):25-34
[3]王方琳,柴成武,赵鹏,等. 3种荒漠植物光合及叶绿素荧光对干旱胁迫的响应及抗旱性评价[J]. 西北植物学报,2021,41(10):1755-1765
[4]刘吉利,赵长星,吴娜,等. 苗期干旱及复水对花生光合特性及水分利用效率的影响[J]. 中国农业科学,2011,44(3):469-476
[5]曾文芳,李亚姝,崔晓宁,等. 施氮对紫花苜蓿光合作用及抗蓟马的影响[J]. 草原与草坪,2021,41(1):61-66,75
[6]孙明伟,徐月乔,王贵,等. 松嫩草地两种生态型羊草根际效应和光合生理对干旱胁迫的响应[J]. 中国草地学报,2021,43(5):8-17
[7]张珍贤,王华,蔡传涛,等. 施肥对干旱胁迫下幼龄期小粒咖啡光合特性及生长的影响[J]. 中国生态农业学报,2015,23(7):832-840
[8]高歌,李正才,葛晓改,等. 施氮对干旱胁迫下毛竹幼苗生物量和根系形态的影响[J]. 生态学杂志,2022,41(5):858-864
[9]赵永平,惠亚云,朱亚,等. 施氮对干旱胁迫紫苏叶绿素含量和光合作用的影响[J]. 广东农业科学,2019,46(10):63-68
[10]周立业,张玉霞,杨秀梅,等. 科尔沁沙地典型人工固沙林群落中少花蒺藜草入侵地土壤特性[J]. 草地学报,2014,22(6):1381-1384
[11]曲婷,周立业. 入侵植物少花蒺藜草异型种子萌发策略及其幼苗生长特性[J]. 草业学报,2022,31(6):91-100
[12]马金宝,张永莉,田迅,等. 科尔沁沙地少花蒺藜草不同生育时期生理适应性的研究[J]. 草原与草坪,2020,40(6):52-57,64
[13]周欣,左小安,赵学勇,等. 科尔沁沙地中南部34种植物叶功能性状及其相互关系[J]. 中国沙漠,2015,35(6):1489-1495
[14]吕朋,左小安,岳喜元,等. 科尔沁沙地封育过程中植被特征的动态变化[J]. 生态学杂志,2018,37(10):2880-2888
[15]HUANG Y,ZHAO X,ZHANG H,et al. Allometric effects of agriophyllum squarrosum in response to soil nutrients,water,and population density in the horqin sandy land of China[J]. Journal of Plant Biology,2009,52(3):210-219
[16]白文娟,郑粉莉,董莉丽,等. 黄土高原水蚀风蚀交错带不同生境植物的叶性状[J]. 生态学报,2010,30(10):2529-2540
[17]吕国利,王进鑫,冯树林,等. 不同类型抗蒸腾剂对4种草本植物叶片水分利用效率的影响[J]. 草地学报,2020,28(3):712-719
[18]邹琦. 植物生理学试验指导[M]. 北京:中国农业出版社:2003:163-166
[19]姜瑞芳. 珙桐幼苗生长与光合特性的主要影响因子[D]. 北京:北京林业大学,2016:14-15
[20]孙延亮,赵俊威,刘选帅,等. 施氮对苜蓿初花期光合日变化、叶片形态及干物质产量的影响[J]. 草业学报,2022,31(9):63-75
[21]南思睿,罗永忠,于思敏,等. 干旱胁迫后复水对新疆大叶苜蓿幼苗光合和叶绿素荧光的影响[J]. 草地学报,2022,30(5):1141-1149
[22]刘婷婷,陈道钳,王仕稳,等. 不同品种高粱幼苗在干旱复水过程中的生理生态响应[J]. 草业学报,2018,27(6):100-110
[23]安玉艳,郝文芳,龚春梅,等. 干旱-复水处理对杠柳幼苗光合作用及活性氧代谢的影响[J]. 应用生态学报,2010,21(12):3047-3055
[24]阎腾飞,黄玉杰,李文杨,等. 干旱胁迫条件下信阳五月鲜桃光合特性和生理生化指标变化规律研究[J]. 节水灌溉,2019(1):45-48,56
[25]韩吉梅,张旺锋,熊栋梁,等. 植物光合作用叶肉导度及主要限制因素研究进展[J]. 植物生态学报,2017,41(8):914-924
[26]王海珍,梁宗锁,韩蕊莲,等. 辽东栎(Quercus liaotungensis)幼苗对土壤干旱的生理生态适应性研究[J]. 植物研究,2005(3):311-316
[27]解婷婷,张希明,梁少民,等. 不同灌溉量对塔克拉玛干沙漠腹地梭梭水分生理特性的影响[J]. 应用生态学报,2008(4):711-716
[28]王庆伟,于大炮,代力民,等. 全球气候变化下植物水分利用效率研究进展[J]. 应用生态学报,2010,21(12):3255-3265
[29]刘吉利,赵长星,吴娜,等. 苗期干旱及复水对花生光合特性及水分利用效率的影响[J]. 中国农业科学,2011,44(3):469-476
[30]杨文权,顾沐宇,寇建村,等. 干旱及复水对小冠花光合及叶绿素荧光参数的影响[J]. 草地学报,2013,21(6):1130-1135
[31]谢贤健,兰代萍,白景文. 三种野生岩生草本植物的抗旱性综合评价[J]. 草业学报.2009,18(4):75-80
[32]ADITI G,ANDRÉS R,CAO A I. The physiology of plant responses to drought[J]. Science,2020,368(6488):266-269
[33]姜野. 干旱胁迫和氮素添加对不同土壤基质少花蒺藜草生理生态特性的影响[D]. 沈阳:辽宁大学,2017:31-32
[34]王慧,王冬梅,张泽洲,等. 外源褪黑素对干旱胁迫下黑麦草和苜蓿抗氧化能力及养分吸收的影响[J]. 应用生态学报,2022,33(5):1311-1319
[35]时丽冉,刘志华. 干旱胁迫对营英菜抗氧化酶和渗透调节物质的影响[J]. 草地学报,2010,18(5):673-677
[36]傅洁. 氮素和水分添加对油蒿群落土壤及植物养分状况的影响[D]. 北京:北京林业大学,2019:13-15
(责任编辑 刘婷婷)
