
豫南地区菱斑食植瓢虫室内外种群抗寒性研究
2024-09-25 00:00:00陈俊华王小盼郭世保侯冰仪张唯一刘红敏
中国瓜菜 2024年9期


摘 要:为明确豫南地区菱斑食植瓢虫(Epilachna insignis)的抗寒能力与低温驯化反应,利用过冷却点测定仪测定其过冷却点和结冰点,并分析其频次分布。结果表明,菱斑食植瓢虫卵的过冷却点(-23.69 ℃)和结冰点(-21.31 ℃)最低;4龄幼虫的过冷却点(-6.77 ℃)和结冰点(-3.93 ℃)最高;菱斑食植瓢虫雌雄成虫过冷却点和结冰点差异显著,雄成虫过冷却点(-19.61 ℃)和结冰点(-17.67 ℃)均显著低于雌成虫过冷却点(-14.23 ℃)和结冰点(-11.49 ℃),且室内种群雌雄成虫过冷却点和结冰点均与自然种群无显著差异。同一虫龄、虫态不同个体间过冷却点与结冰点的变化均符合正态分布。抗寒驯化处理结果表明,随处理温度的降低,雌成虫的死亡率逐渐升高,各冷藏驯化处理过冷却点差异均不显著,以8 ℃冷藏驯化结冰点最低(-13.99 ℃)。
关键词:菱斑食植瓢虫;过冷却点;结冰点;抗寒性
中图分类号:S47 文献标志码:A 文章编号:1673-2871(2024)09-159-07
Study on the cold resistance of indoor and natural population of Epilachna insignis in southern Henan province
CHEN Junhua 1, 2, WANG Xiaopan1, GUO Shibao1, 2, HOU Bingyi1, ZHANG Weiyi1, LIU Hongmin1, 2
(1. Xinyang Agriculture and Forestry University, Xinyang 464000, Henan, China; 2. Xinyang Research Center of Engineering and Technology for Crop Pest Control and Prevention, Xinyang 464000, Henan, China)
Abstract: In order to clarify the cold-resistance and low-temperature acclimation of Epilachna insignis in southern Henan province, the supercooling point and freezing point were determined by using the supercooling point meter, and the frequency distribution of the supercooling point was analyzed. The results showed that the supercooling point(-23.69 ℃)and freezing point(-21.31 ℃)of eggs were the lowest. The supercooling point(-6.77 ℃) and freezing point(-3.93 ℃)of the 4th instar larva were the highest, and there were significant differences in the supercooling point and freezing point between male and female adults of E. insignis. The supercooling point (-19.61 ℃) and freezing point (-17.67 ℃) of male adults were significantly lower than those of female adults (-14.23 ℃) and(-11.49 ℃), and the supercooling and freezing points of male and female adults in the indoor population were not significantly different from those in the natural population. The changes in supercooling point and freezing point of different individuals of the same instar and different stages were in accordance with normal distribution. The results of cold acclimation showed that the mortality of female adults increased gradually with the decrease of the treatment temperature. There was no significant difference in the supercooling point among the treatments, and the lowest freezing point(-13.99 ℃) was at 8 ℃.
Key words: Epilachna insignis, Supercooling point; Freezing point; Cold resistance
收稿日期:2024-01-05;修回日期:2024-02-25
基金项目:河南省重大专项创新示范专项(201111310900);河南省卓越农林人才教育基地建设项目
作者简介:陈俊华,女,副教授,主要从事害虫生态防控研究。E-mail:junhua993@163.com
通信作者:刘红敏,女,教授,主要从事害虫绿色防控研究。E-mail:liuhongm8@sina.com
菱斑食植瓢虫(Epilachna insignis)属瓢甲科食植瓢虫亚科食植瓢虫属Epilachna。成虫虫体中型,虫体两鞘翅末端略尖削,背面隆起呈半球形,体被黄色细毛,前胸背板有1个黑色的斑点,一侧鞘翅上有7个黑色斑点,各斑点呈菱形或近似心形[1-3]。幼虫共4龄,体黄色,体背布满枝刺,每龄蜕皮后枝刺黄色,后逐渐变为黑色[4-6]。
菱斑食植瓢虫在我国分布广泛,它的自然寄主包括栝楼、茄子和龙葵等多种茄科和瓜类植物[1,4,7],近些年来在河南、四川、山东、陕西、福建、广东、安徽、云南等地都有发生,幼虫喜食瓜蒌叶片,严重时幼虫3~5 d可食尽叶肉,使叶片呈网络状,严重影响瓜蒌的生长[6-7]。国内外已有研究多集中在菱斑食植瓢虫的形态特征识别[3]、生物学观察[5]和综合防控等方面[5-6]。
昆虫属于变温动物,温度对其生长、发育、成活率、扩散和分布有重要的影响,当遇到低温时昆虫会自动调节自身的温度来抵御低温环境[8]。其中,在昆虫的耐寒性研究中,过冷却点和结冰点是反映昆虫耐寒性强弱的主要指标[7-9]。昆虫体液含有糖、蛋白质和脂肪等物质,使其能承受0 ℃以下的低温,这种现象称为过冷却现象。过冷却理论认为当昆虫体温下降到0 ℃时(N1),体液并不结冰,当体温继续下降到某一温度(T1)时,因体液开始结冰而释放少量热量,虫体温度会出现短暂上升以接近0 ℃(N2),随之虫体温度持续下降,昆虫体液开始结冰,此时,N2称之为体液结冰点,T1称为过冷却点[10]。温带地区昆虫体温会受季节温度上下变动,一般低温来临前,一些昆虫转入特殊越冬场所或进入某一特定发育阶段来对抗低温的不利影响,同时很多昆虫可通过调整自身生理生化状态或能量重塑储存形式,如昆虫身体颜色改变、减小体型、减轻体重、降低体内游离水比例、增加脂肪类和糖类等物质含量来适应不良低温环境[7-9,11]。
低温驯化是将昆虫置于较低温度(亚致死低温)一段时间以提高其在低温下存活率的过程[12],一般合适的低温驯化可以不同程度地提高昆虫抵抗低温的能力,低温驯化一般可分为快速冷驯化(RCH)和长时冷驯化(ACC)两类[11-12]。快速冷驯化的处理温度一般较低,可在十几小时或几小时甚至更短时间内提高昆虫的耐寒能力,是昆虫适应自然状态中温度骤变或短期低温环境的一种有效策略[13]。随着全球气温升高、异常气候频现,昆虫对季节、环境等变化的适应或应对能力引起广泛关注[14]。低温驯化是通过不同环境温度突变的模拟从而增强昆虫应对不良环境的适应性,是昆虫关键的越冬策略[15-16]。
为探索菱斑食植瓢虫在豫南地区的越冬状况和抗寒性,通过测定菱斑食植瓢虫各虫态、虫龄的过冷却点和结冰点,以及不同低温抗寒驯化对其过冷却点和结冰点的影响,初步确定其抗寒能力以及秋冬季温度变动对菱斑食植瓢虫抗寒性的影响,分析不同虫龄、虫态的抗寒能力强弱,为菱斑食植瓢虫越冬区划分、虫害防控和发生期预测等提供理论依据。
1 材料与方法
1.1 材料
供试虫源:于2023年6月中旬至7月上旬自河南省信阳市信阳农林学院内栝楼上采集菱斑食植瓢虫卵1200粒左右,置于25 cm×25 cm×25 cm的养虫盒中,盒下部铺浸湿润的滤纸保湿,放入温度为(25±2) ℃、相对湿度为(70%±10%)、光周期L/D为14 h/10 h的养虫室待其孵化,放入新鲜的栝楼叶片饲养,每天及时补充新鲜叶片并吸走养虫盒盒壁水珠,待其化蛹后挑出放入高40 cm的圆柱形养虫笼待羽化,羽化后放入新鲜栝楼叶片、枝条任其自由交配,收集虫卵作为供试虫源。栝楼叶片均在河南省信阳市信阳农林学院校内采摘,于7-11月春夏秋生长季分批采摘。
1.2 测量方法
室内种群:从菱斑食植瓢虫养虫盒中随机选取个体发育一致的菱斑食植瓢虫卵、1~4龄幼虫、蛹、雄成虫和雌成虫各30头以上个体,采用昆虫过冷却点测定仪及其配套测定软件测定其过冷却点和结冰点。测定时虫体较小的卵置于10 μL容量移液器枪头底部,将感温探头与之充分接触,多余空隙处塞紧脱脂棉。1~4龄幼虫、蛹和成虫单只置于1.5 mL离心管底部,将探头插入离心管与虫体接触,用脱脂棉塞满离心管内多余的空隙。将探头置于超低温冷冻储存箱中,一端与计算机相连的过冷却点测试仪记录虫体温度变化,每0.5 s显示1次虫体温度,出现过冷却点和结冰点折线拐点后试验停止。
室外种群:于11月上中旬采集室外雌雄成虫带回实验室测定其过冷却点和结冰点,测量方法同室内种群。
1.3 低温驯化方法
豫南地区冬季室外的日最低平均温度为5 ℃左右,设置冷藏驯化温度分别为2、4、8 ℃。将羽化后菱斑食植瓢虫雌成虫2日龄种群分别置于冷藏温度下12 h,超低温冷冻储存箱温度以1 ℃·min-1 的速率下降至2、4、8 ℃,以 25 ℃ 常温下不冷藏的菱斑食植瓢虫作为对照。每个处理30头成虫,3次重复。处理结束后取出放于室温下24 h后测量其过冷却点和结冰点,同时计算不同处理菱斑食植瓢虫的存活率。
存活率/%=处理后存活头数/每个处理总头数×100。
1.4 数据分析
采用IBM SPSS Statistics 22单因素方差分析(one-way ANOVA)法对菱斑食植瓢虫各虫龄、虫态、雌蛹、雄蛹及雌成虫、雄成虫之间的过冷却点和结冰点进行分析,运用K-S法检验其是否符合正态分布,采用Duncan新复极差法进行多重比较。频次分布图采用Microsoft Excel 2019软件绘制。
2 结果与分析
2.1 菱斑食植瓢虫各虫龄、虫态的过冷却点和结冰点
由表1可知,菱斑食植瓢虫以卵的过冷却点最低,为-23.69 ℃,最低个体达-28.14 ℃,与其他各虫龄、虫态均差异显著;4龄幼虫的过冷却点最高,为-6.77 ℃,过冷却点最高个体达-3.02 ℃。在幼虫发育阶段,菱斑食植瓢虫随着虫龄增大,其过冷却点逐渐升高,直至幼虫化蛹后过冷却点开始下降;雄成虫、雌成虫室内种群的过冷却点均与自然种群差异不显著,且雄成虫的过冷却点显著低于雌成虫。表明随着菱斑食植瓢虫幼虫的生长,其抗寒能力逐渐减弱,进入蛹期阶段后,菱斑食植瓢虫抗寒性逐步增强。
菱斑食植瓢虫不同虫龄、虫态间结冰点存在差异,且菱斑食植瓢虫结冰点高低的变化趋势与过冷却点近似,菱斑食植瓢虫卵的体液结冰点最低,推测其耐寒性最强,4龄幼虫的体液结冰点最高,推测其耐寒性最弱。不同发育阶段菱斑食植瓢虫的体液结冰点普遍高于过冷却点。结冰点由高到低依次为:4龄幼虫>3龄幼虫>蛹>2龄幼虫>1龄幼虫>自然种群雌成虫>雌成虫>自然种群雄成虫>雄成虫>卵。蛹的结冰点高于1龄与2龄幼虫,但均差异不显著;雌成虫的最高结冰点为-4.87 ℃,雄成虫的最高结冰点为-15.78 ℃;雌雄成虫的室内种群的结冰点均与自然种群无显著差异。
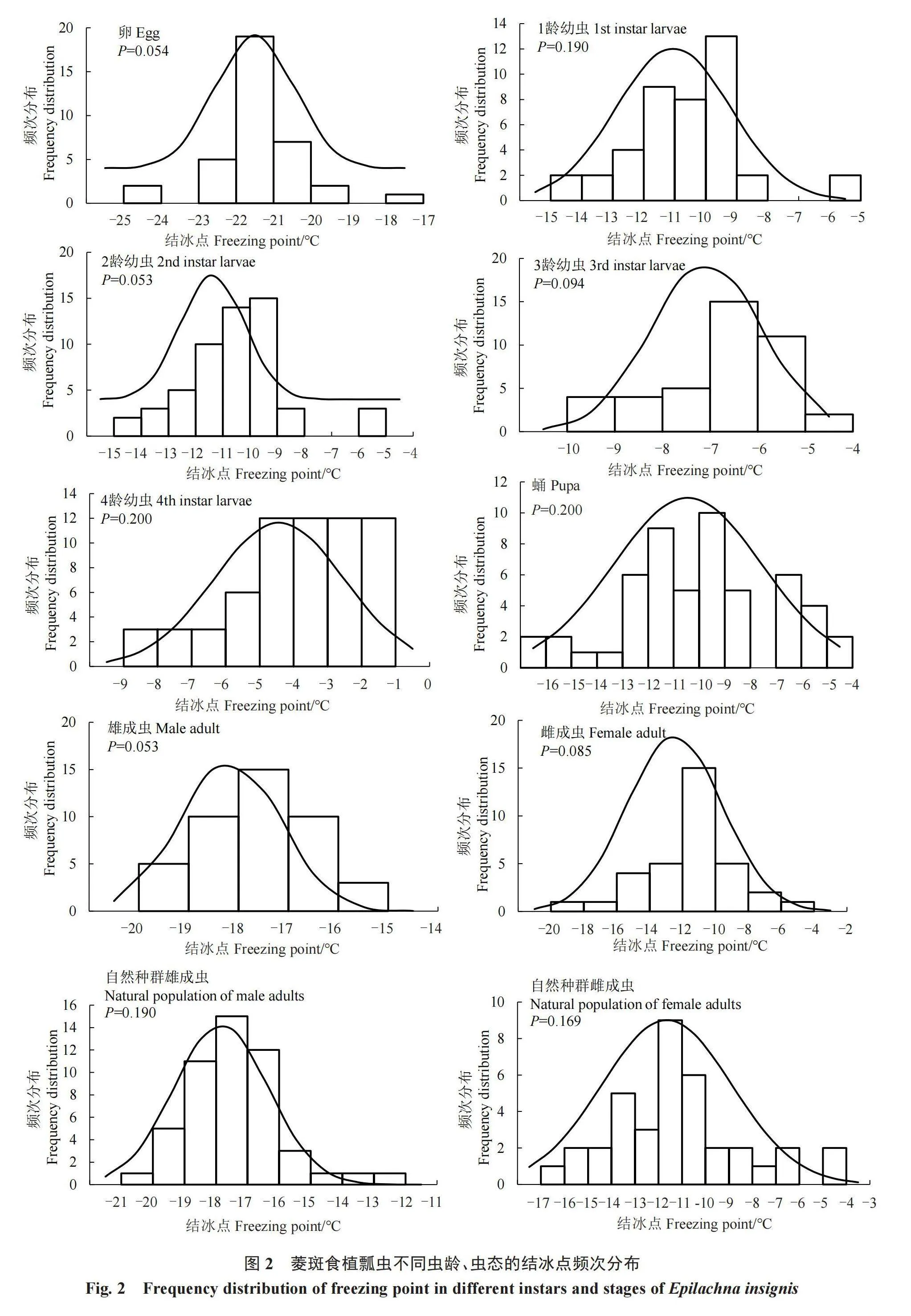
2.2 菱斑食植瓢虫不同虫龄、虫态过冷却点和体液结冰点的频次分布
不同虫龄、虫态的菱斑食植瓢虫过冷却点的分布情况存在一定范围的宽度,表明不同个体间存在一定程度的差异,分布均符合正态分布(图1)。过冷却点分布以蛹的分布范围最广,分布于-21.82~-7.67 ℃;以雄成虫的过冷却点分布范围最窄,为-22.01~-16.61 ℃。不同虫龄、虫态的菱斑食植瓢虫的过冷却点范围分布宽度由大到小依次为:蛹(14.15 ℃)>自然种群雌成虫(11.65 ℃)>2龄幼虫(10.44 ℃)>1龄幼虫(10.43 ℃)>雌成虫(10.01 ℃)>卵(8.93 ℃)>自然种群雄成虫(8.70 ℃)>3龄幼虫(8.19 ℃)>4龄幼虫(7.66 ℃)>雄成虫(5.40 ℃)。结果表明,蛹、自然种群雌成虫对低温的适应能力相对较强,判断其能适应冬季低温变化的幅度较宽。
不同虫龄、虫态的菱斑食植瓢虫体液的结冰点存在一定程度的差异,但均符合正态分布(图2)。菱斑食植瓢虫的体液结冰点分布均有一定的范围宽度,结冰点分布范围波动最小的是雄成虫,为3.74℃;结冰点分布范围波动最大的是雌成虫,为14.65 ℃。体液结冰点频次分布范围宽度由大到小排列顺序为:雌成虫(14.65 ℃)>蛹(11.87 ℃)>自然种群雌成虫(11.60 ℃)>1龄幼虫(8.33 ℃)>2龄幼虫(8.32 ℃)>自然种群雄成虫(7.89 ℃)>4龄幼虫(7.20 ℃)>卵(6.95 ℃)>3龄幼虫(5.14 ℃)>雄成虫(3.74 ℃)。雌成虫、蛹和自然种群雌成虫的个体间结冰点变化幅度比较大,推测其对低温的调节适应能力较强。从体液结冰点频次分布图来看,雌成虫无论自然种群还是室内种群,其体液结冰点的变化范围均大于雄成虫。
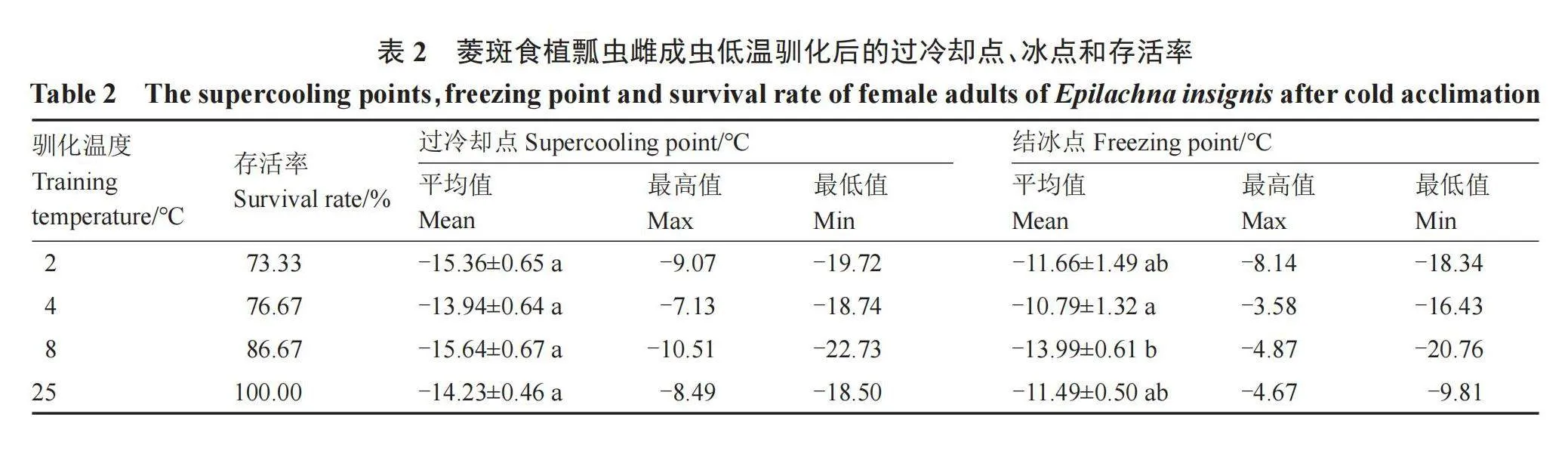
2.3 菱斑食植瓢虫雌成虫低温驯化后的过冷却点、结冰点和存活率
由表2可知,随着处理温度的降低,菱斑食植瓢虫雌成虫存活率逐渐升高。各冷藏处理12 h菱斑食植瓢虫雌成虫过冷却点处理间差异不显著;各冷藏处理以8 ℃驯化12 h结冰点最低,为-13.99 ℃,但与对照之间差异不显著。低温驯化结冰点由高到低依次是:4 ℃(-10.79 ℃)>25 ℃(-11.49 ℃)>2 ℃(-11.66 ℃)>8 ℃(-13.99 ℃)。
3 讨论与结论
昆虫冬季忍受极端低温情况决定了昆虫种群的扩散分布与翌年的发生情况[17]。菱斑食植瓢虫作为一种重要的植食性昆虫,对栝楼等造成严重危害,对其进行耐寒性研究是有重要意义的[5-6]。本研究结果表明,菱斑食植瓢虫以卵过冷却点和结冰点最低,4龄幼虫过冷却点和结冰点最高。刘奎等[18]研究了叶甲科椰心叶甲(Brontispa longissima)各虫态的过冷却点、结冰点,结果表明,椰心叶甲各虫态均以卵的过冷却点和结冰点最低,以4龄幼虫的过冷却点和结冰点最高,与本研究结果一致。韩光荣[19]研究了瓢甲科端尖食植瓢虫(Epilachna quadricollis)不同发育阶段的过冷却点与结冰点,结果表明最高为1龄幼虫,最低为3龄幼虫,与本研究结果存在差异。耿书宝等[20]研究表明,茶银尺蠖(Sopula subpunctaria)的幼虫随着龄期的增加,其过冷却点逐渐升高,而本研究结果随着菱斑食植瓢虫幼虫龄期的增加,其过冷却点和结冰点升高。笔者调查表明,菱斑食植瓢虫在豫南地区主要以成虫越冬,越冬成虫多隐蔽于阳面建筑、杂草等缝隙处,早春成虫出蛰早,出蛰成虫即取食补充营养,在寄主枝蔓或叶片背面产卵,卵裸露,早春豫南气温波动大,推测是卵期过冷却点低的原因之一。昆虫的过冷却点还与体内含水量、体内脂类物质及体内糖原物质等有关[14,21],菱斑食植瓢虫的抗寒性是否与其体内物质含量等有关仍需进一步研究。此外,Zhang等[22]研究表明,miRNA介导的蛔苷信号能够调控松墨天牛(Monochamus alternatus)的抗寒性。
本研究结果表明,菱斑食植瓢虫各发育阶段的过冷却点和结冰点的频次分布均有一定的范围,不同个体间存在一定的变化幅度,这种现象反映出昆虫对自然环境变化的适应策略[23]。在研究室内种群和自然种群中,雌成虫的过冷却点和结冰点均高于雄成虫,频次分布图显示变化幅度雌成虫高于雄成虫,表明雌成虫越冬期间存在超过10 ℃的温度变化范围,可增加其越冬存活量。耿书宝等[24]研究灰茶尺蠖(Ectropis grisescens)雌雄成虫之间的过冷却点,结果为雄成虫低于雌成虫,且存在显著差异,与本研究中菱斑食植瓢虫雄成虫显著低于雌成虫的结果一致。魏淑花等[25]研究白纹雏蝗(Glyptobothrus albonemus)雌雄成虫体内的自由水与结合水比值以及亚油酸甲酯、油酸甲酯和亚麻酸甲酯等的含量也不一样。Fan等[26]研究温度可影响柑橘木虱(Diaphorina citri)的体色、体型,体重等,低温下多出现体小体轻个体,与能量转换积累有关。笔者研究中也发现菱斑食植瓢虫的雄性个体略小于雌性个体,温度越低表现越明显,其生理机制需进一步探究。
此外,研究证明很多昆虫在低温的刺激下启动抗寒基因的表达,合成抗冻蛋白、热激蛋白、甘油、山梨醇、海藻糖等物质[11,27]。室内种群经2、4和8 ℃抗寒暴露处理,结果表明,8 ℃低温处理过冷却点和结冰点与对照有所下降,均无显著差异,但个体间过冷却点和结冰点的变化幅度增大,尤其结冰点愈加明显。有研究表明,分别冷藏于不同变温条件下的食蚜瘿蚊(Aphidoletes aphidimyza)表现出不同的抗寒能力[28];不同地理种群甜菜夜蛾(Spodoptera exigua)的抗寒能力也不同[29]。
本研究中菱斑食植瓢虫雌成虫经低温驯化处理后存活率均随着温度的降低而逐渐降低,且都高于50%,表明菱斑食植瓢虫多数成虫可在豫南地区安全越冬。此外,笔者测定了自然条件下菱斑食植瓢虫越冬虫态的过冷却点和结冰点,结果与室内种群无显著差异,表明室内种群过冷却点和结冰点能反映菱斑食植瓢虫野外自然种群的抗寒能力。此外,通过对菱斑食植瓢虫越冬虫态雌雄成虫过冷却点和结冰点的测定,结果表明,菱斑食植瓢虫在适生植物充足的条件下,在我国南北方多数地区均有严重发生危害的可能性。
参考文献
[1] 张巍巍,李元胜.中国昆虫生态大图鉴[M].重庆:重庆大学出版社,2019.
[2] 曾涛,庞雄飞.食植瓢虫属Epilachna中国新记录种及两种幼虫的形态记述(鞘翅目:瓢虫科)[J].昆虫分类学报,2001,23(2):114-119.
[3] 耿云冬.山西省瓢虫科分类研究[D].北京:首都师范大学,2007.
[4] 尹健,熊建伟,孙万慧,等.信阳栝楼2种主要害虫的初步研究[J].河南农业科学,2007(1):51-53.
[5] 尹健.信阳栝楼的人工栽培及主要害虫的发生、防治技术研究[D].武汉:华中农业大学,2006.
[6] 王胜宝,张先平.菱斑食植瓢虫危害栝楼研究初报[J].陕西农业科学,1996(5):29-30.
[7] 张润志.菱斑食植瓢虫Epilachna insignis Gorham幼虫[J].应用昆虫学报,2014,51(2):399.
[8] 景晓红,康乐.昆虫耐寒性的测定与评价方法[J].昆虫知识,2004,41(1):7-10.
[9] 崔双双,贺一原.昆虫的耐寒性及其影响因素[J].生命科学研究,2011,15(3):273-276.
[10] 雷朝亮,荣秀兰,韩召军,等.普通昆虫学[M].北京:中国农业出版社,2003:441-442.
[11] 唐斌,林青青,邬梦静,等.抗寒类蛋白与冷驯化诱发昆虫耐寒的生理调节研究[J].环境昆虫学报,2014,36(5):805-813.
[12] 岳雷,郭建英,周忠实,等.冷驯化对昆虫耐寒性及其适合度的影响[J].中国生物防治学报,2013,29(2):286-293.
[13] LEE R E.Insect cold-hardiness:To freeze or not to freeze:How insects survive low temperatures[J].BioScience,1989,39(5):308-313.
[14] 巴都木才茨克.低温驯化对斑须蝽耐寒性的影响[D].呼和浩特:内蒙古师范大学,2022.
[15] 王宪辉,齐宪磊,康乐.昆虫的快速冷驯化现象及其生态适应意义[J].自然科学进展,2003,13(11):1128-1133.
[16] SAKAI M,SUZUKI K,GOTO M.Factors affecting carbohydrate and free amino acid content in overwintering larvae of Enosima leucotaeniella[J].Journal of Insect Physiology,1997,44(1):87-91.
[17] LI C C,PEI J H,LI J L,et al.Overwintering larval cold tolerance of Sirex noctilio (Hymenoptera:Siricidae):Geographic variation in northeast China[J].Insects,12(2):116.
[18] 刘奎,彭正强,李文德,等.椰心叶甲过冷却点的测定[J].植物检疫,2005(1):24-26.
[19] 韩光荣.端尖食植瓢虫和柳蓝叶甲的过冷却点测定[J].安徽农业科学,2017,45(18):135-136.
[20] 耿书宝,侯贺丽,洪枫,等.不同虫态茶银尺蠖过冷却点和结冰点的测定[J].植物保护学报,2022,49(4):1180-1186.
[21] 任金龙,赵莉,葛婧,等.意大利蝗卵的抗寒能力和含水量的季节变化[J].植物保护学报,2021,48(1):179-186.
[22] ZHANG B,ZHAO L L,NING J,et al.MiR-31-5p regulates cold acclimation of the wood-boring beetle Monochamus alternatus via ascaroside signaling[J].BMC Biology,2020,18:184.
[23] 张智,郑乔,张云慧,等.草地贪夜蛾室内种群抗寒能力测定[J].植物保护,2019,45(6):43-49.
[24] 耿书宝,侯贺丽,乔利,等.灰茶尺蠖过冷却点与结冰点的测定[J].中国植保导刊,2023(4):5-10.
[25] 魏淑花,吴秀芝,王颖,等.白纹雏蝗抗寒特性和高温耐受力及其机制[J].植物保护学报,2021,48(1):172-178.
[26] FAN J Y,SHANG F,PAN H M,et al.Body color plasticity of Diaphorina citri reflects a response to environmental stress[J].Insect Science,2024,31(3):937-952.
[27] 段小凤,王晓庆,李品武,等.冷驯化对茶尺蠖抗寒性生理指标的影响[J].应用昆虫学报,2015,52(6):1413-1420.
[28] 袁清婷,苟剑渝,黄纯杨,等.不同冷藏条件对食蚜瘿蚊过冷却点和冰点的影响[J].应用昆虫学报,2022,59(2):399-405.
[29] 韩兰芝,翟保平,张孝羲.甜菜夜蛾的抗寒力研究[J].植物保护学报,2005,32(2):169-173.
