
牧草盲蝽越冬成虫出蛰后飞行能力研究
2024-06-24 08:42:45郑艺翔李太隆罗棚冯宏祖
中国棉花 2024年5期
关键词:越冬
郑艺翔 李太隆 罗棚 冯宏祖
收稿日期:2023-11-07 第一作者简介:郑艺翔,硕士研究生,研究方向为农业昆虫与害虫治理,946957451@qq.com。 *通信作者:冯宏祖,博士,教授,研究方向为农业昆虫与害虫治理,fhzzky@163.com
基金项目:国家自然科学基金(32272539)
A study on the flight ability of overwintering adults of Lygus pratensis after hibernation
Zheng Yixiang, Li Tailong, Luo Peng, Feng Hongzu*
摘要:明确牧草盲蝽Lygus pratensis越冬成虫出蛰后不同时间的飞行能力,为牧草盲蝽越冬后的扩散区域预测研究和防控策略的制定提供参考。采用JIADUO昆虫飞行信息系统,在人工气候箱内对越冬后5个日期的牧草盲蝽进行飞行能力测试,比较种群飞行率、飞行速率、总飞行时间和总飞行距离。结果表明,3月24日刚出蛰的成虫基本不能飞行;4月13日的飞行能力最强,种群飞行率为63.89%,平均飞行速率为1.687 km·h-1,总飞行距离为1.609 km,总飞行时间为0.970 h;4月3日的飞行能力次之;4月23日和5月3日飞行能力逐渐减弱。综上,牧草盲蝽越冬成虫的飞行能力与出蛰后的时间关系密切,随出蛰后时间的推移呈现先增强后逐渐减弱的趋势。
关键词:牧草盲蝽;越冬;飞行能力;飞行磨;出蛰
牧草盲蝽Lygus pratensis是新疆重要的农业害虫。近年来由于新疆农作物种植结构、耕作制度和栽培模式的变化,再加上气候因素等影响,牧草盲蝽再度暴发成灾[1-2]。它大发生时,棉田蕾铃被害率达到56%;同时,有的会转移至香梨、苹果、红枣、核桃等果园危害花蕾及幼果,轻者会造成黑色的坏死斑,导致果实畸形,重者造成大量落花落果。牧草盲蝽成虫对开花植物有明显的趋好性,春夏季节从越冬场所、早期虫源地向大田迁飞扩散,也会在大田寄主作物之间扩散[3-4],这与其飞行能力有重要联系,但飞行能力随出蛰后时间的变化的研究未见报道。本研究借助飞行磨系统在实验室条件下测定了牧草盲蝽越冬后成虫出蛰后的飞行能力,以期为牧草盲蝽越冬后扩散范围的预测和防控策略的制定提供参考。
1 材料与方法
1.1 供试虫源
供试牧草盲蝽采自新疆阿拉尔市9团小麦地、苜蓿地,均为2023年早春自然越冬出蛰后的成虫。将其放入自制养虫盒内,在温度24 ℃、光周期(光暗比,下同)15∶9、相对湿度70%±10%、光照强度(480±12) lx的RXZ型智能人工气候箱内饲养,以供试验使用。
1.2 牧草盲蝽越冬成虫的飞行能力测定
本试验使用的飞行测试系统为JIADUO牌昆虫飞行信息系统(鹤壁嘉多卫农农林科技有限责任公司),由飞行磨主机(FXM-Z,24路)和昆虫小飞行磨(FXM-X,1个2路)组成。昆虫飞行传动装置由吊臂(直径0.1 mm、长5 cm铜丝)、2号昆虫针、黑色遮光纸(长0.5 cm、宽0.1 cm)制作。将采集到的牧草盲蝽在养虫盒内使用花椰菜饲养24 h补充能量,在第2日9:00开始飞行测试,测试前先用CO2处理牧草盲蝽25 s使其昏迷,再将吊臂顶端用502胶(编号:4167)与牧草盲蝽背板粘连。待牧草盲蝽苏醒后,将飞行传动装置放置在昆虫小飞行磨的2块磁铁中间,再将小飞行磨放入RXZ型智能人工气候箱内进行测试,测试条件:温度24 ℃、光周期15∶9、相对湿度70%±10%、光照强度(480±12) lx,测试时长为24 h。
试验从3月份第1次采集到田间自然出蛰的越冬成虫起,每隔10 d采集1次雌虫进行飞行能力测试,即于3月24日、4月3日、4月13日、4月23日、5月3日共采集5次,即为5个日期处理,每个处理20个重复(20头)。测定各处理种群飞行率(能飞个体比例)、总飞行距离、总飞行时间、飞行速率等参数。能飞个体的判断参考Stewart等[5]提出的标准,即总飞行时间>10 min为能飞个体。
1.3 数据分析
用SPSS 25.0对总飞行距离、总飞行时间、飞行速率数据进行分析处理。先对原始数据进行正态检验,若不符合正态分布,对数据进行对数转换。符合正态分布时,再采用邓肯多重范围检验(新复极差法)比较差异性,差异显著水平为0.05。
2 结果与分析
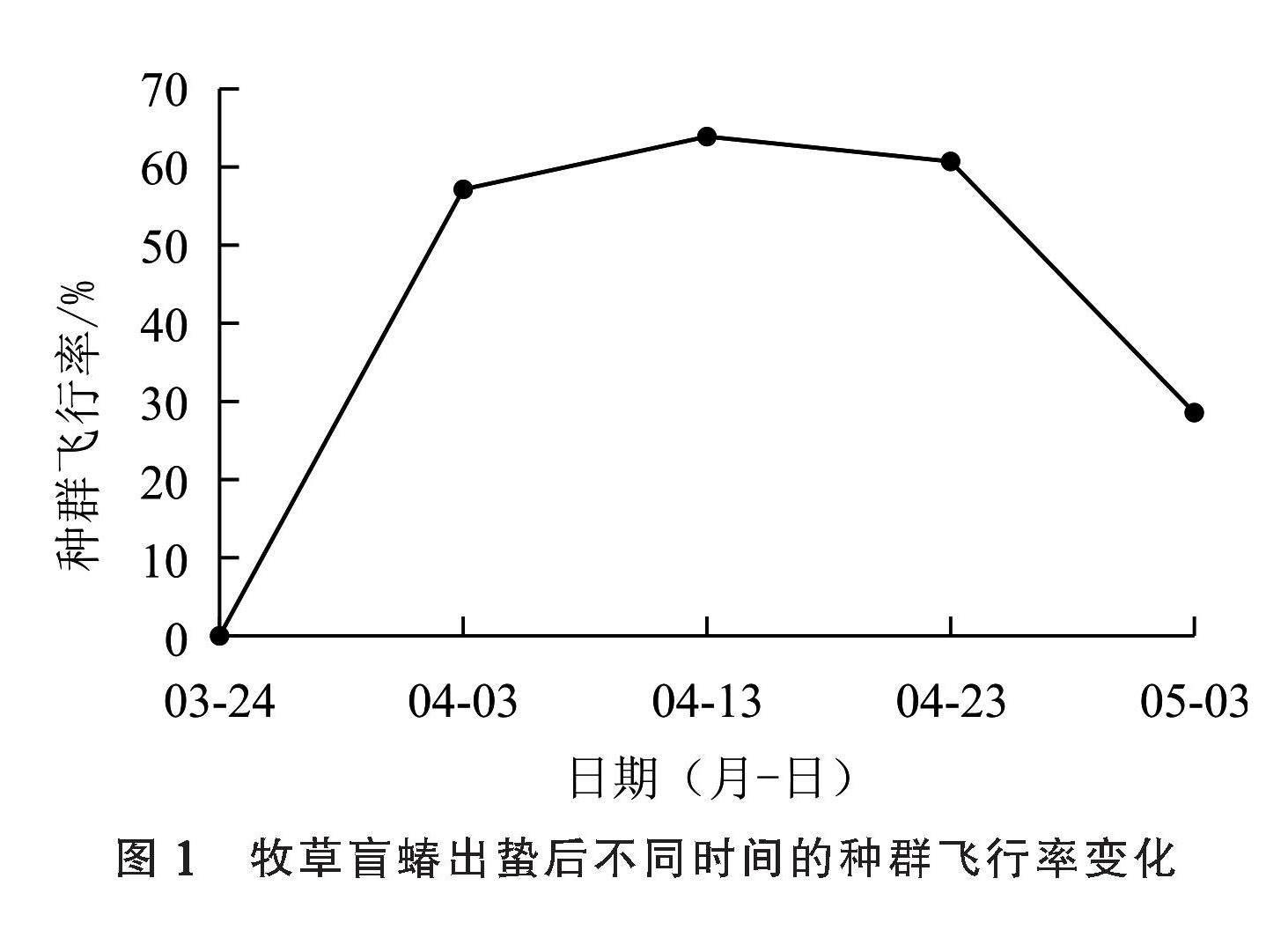
2.1 牧草盲蝽出蛰后不同时间的种群飞行率
根据种群动态调查结果(图1),在3月24日第1次采集的牧草盲蝽种群飞行率为0%,此时的牧草盲蝽没有飞行能力;在之后4个日期的比较中可以看出,随着出蛰后时间的推移,种群飞行率呈先上升后下降的趋势,在4月13日种群飞行率最高达到63.89%,随后开始下降,在5月3日为28.57%。
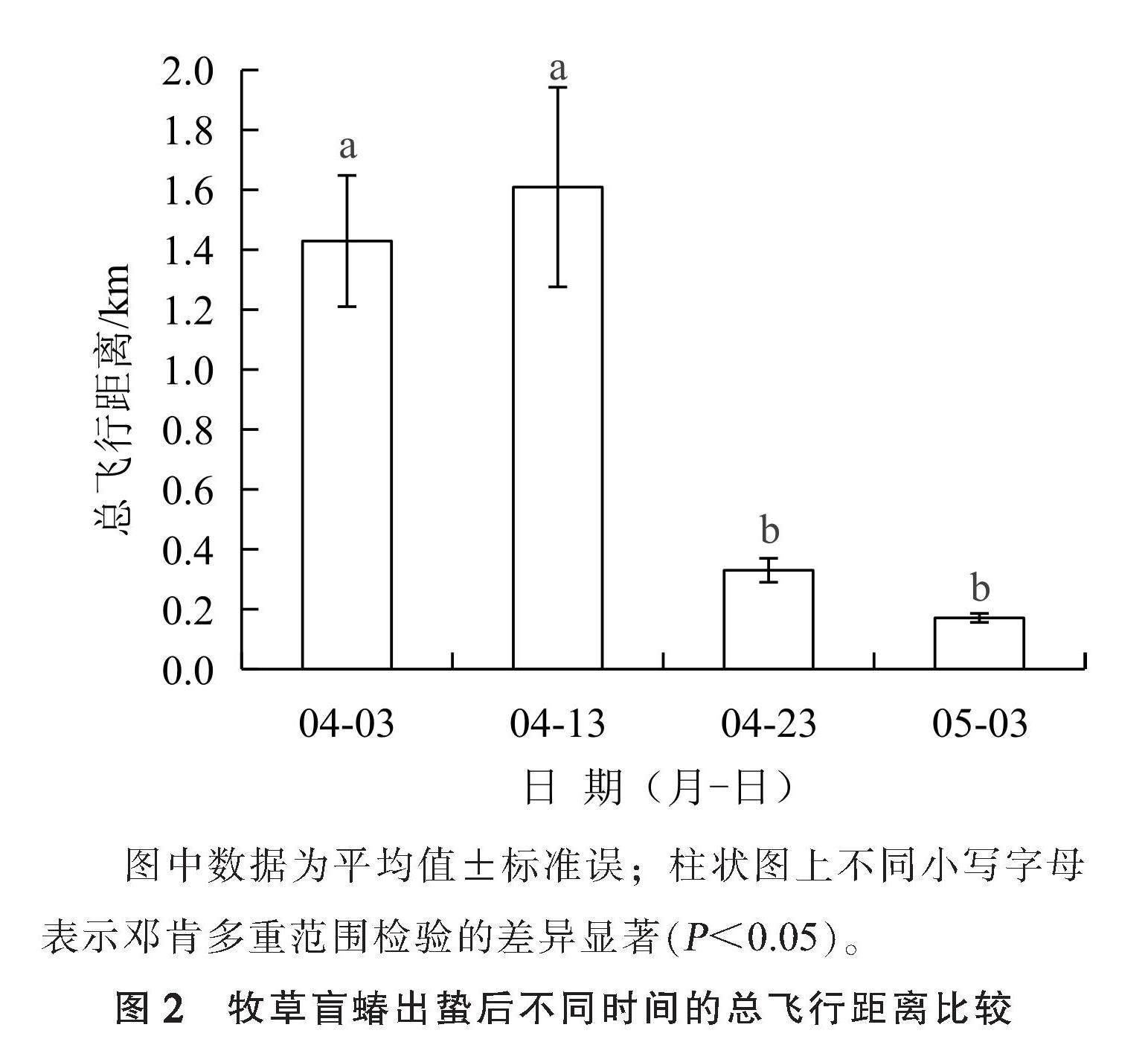
2.2 牧草盲蝽出蛰后不同时间的总飞行距离
从牧草盲蝽出蛰后不同时间的总飞行距离(图2)可以看出,总飞行距离随着出蛰后时间的推移呈现先上升后下降的趋势。牧草盲蝽的总飞行距离在4月3日为(1.429±0.219) km,在4月13日达到最大,为(1.609±0.333) km,这2个日期之间差异不显著;随后开始呈下降趋势,4月23日为(0.330±0.040) km,5月3日最小,为(0.171±0.015) km,这2个日期之间差异也不显著,但与4月3日和4月13日调查的总飞行距离有显著差异。
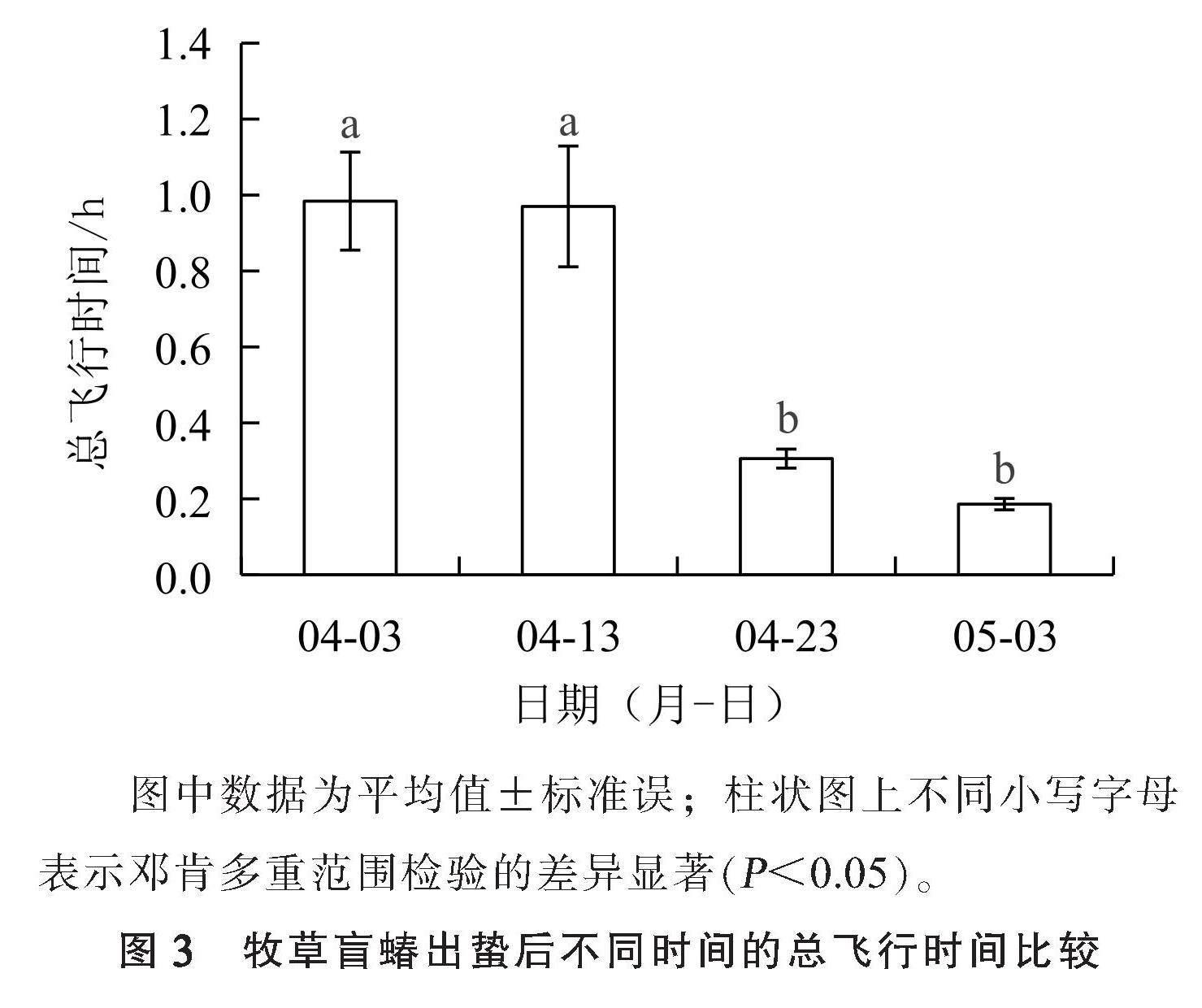
2.3 牧草盲蝽出蛰后不同时间的总飞行时间
从牧草盲蝽出蛰后不同时间的总飞行时间(图3)可以看出,总飞行时间随着日期的推移呈下降趋势。4月3日,其总飞行时间最长,为(0.984±0.129) h,4月13日总飞行时间与之接近,为(0.970±0.159) h,两者差异不显著;4月23日和5月3日总飞行时间明显下降,分别为(0.306±0.025) h和(0.186±0.015) h,两者差异不显著,但与4月3日和4月13日调查的总飞行时间有显著差异。
2.4 牧草盲蝽出蛰后不同时间的飞行速率
从牧草盲蝽出蛰后不同时间的飞行速率(图4)可以看出,不同时间最大飞行速率呈现先上升后下降的趋势。其中:4月13日最大飞行速率最高,为(3.128±0.312) km·h-1,与其他时间的最大飞行速率存在显著差异;其次是4月3日的最大飞行速率,为(2.496±0.223) km·h-1,与4月23日最大飞行速率(1.615±0.110) km·h-1、5月3日的最大飞行速率(1.750±0.110) km·h-1存在显著差异;而4月23日的最大飞行速率最低,与5月3日的差异不显著。
牧草盲蝽在出蛰后不同时间的平均飞行速率随着时间的推移呈先上升后下降的趋势(图4)。其中:4月13日的平均飞行速率最高,为(1.687±0.190) km·h-1,与4月3日的平均飞行速率(1.423±0.098) km·h-1差异不显著;4月23日平均飞行速率下降为(1.034±0.051) km·h-1;5月3日的平均飞行速率最低,为(0.907±0.063) km·h-1,与4月23日的平均飞行速率差异不显著,但两者与4月3日、4月13日的平均飞行速率存在显著差异。
3 结论与讨论
本研究结果表明,牧草盲蝽越冬成虫出蛰后时间与飞行能力关系密切:3月24日牧草盲蝽种群飞行率为0%,不具备飞行能力;随着时间的推移,牧草盲蝽的飞行能力从4月3日起呈现先增强后减弱的趋势;4月13日飞行能力最强,种群飞行率为63.89%,平均飞行速率为(1.687±0.190) km·h-1,最大飞行速率为(3.128±0.312) km·h-1,总飞行距离为(1.609±0.333) km,总飞行时间为(0.970±0.159) h,但除最大飞行速率外,均与4月3日无显著差异。前人研究表明,昆虫在自然条件下为了抵御冬季的寒冷进化出了适应低温的特征[6],为确保能在不利的条件下存活,在春季温度适宜时才开始活动[7]。其中,牧草盲蝽越冬代成虫在春季温度超过9 ℃时开始出蛰[8]。由此推测,3月下旬到5月上旬当地田间温度逐渐升高,可能是牧草盲蝽飞行能力随出蛰后时间推移而增强的主要原因,在麦长管蚜Sitobion avenae[9]、草地螟阿格姬蜂Agrypon flexorius Thunberg[10]、桔小实蝇Bactrocera dorsalis[11]、绿盲蝽Apolygus lucorum等盲蝽[12]的研究中发现,它们的飞行能力会随着温度升高而增强,在最适温度达到最强。越冬后的昆虫成虫会经历交配、产卵和日龄增长[13],这些昆虫的生长发育进程及繁殖对昆虫飞行能力都有显著影响[14]:从羽化开始随着日龄增加飞行能力逐渐增强,但随着接近生命的末期飞行能力减弱[12,15-16];交配会使雌性成虫飞行能力增强,从而使雌虫能在更大范围内寻找寄主产卵[17],而产卵会使飞行能力减弱[18]。牧草盲蝽成虫出蛰后的飞行能力总体上随时间的延长而减弱,是否与成虫产卵、日龄有关,还需要进一步研究。
本研究在室内适宜的温度、湿度、光照条件下对田间自然采集的牧草盲蝽进行飞行能力的测试,可以真实反映出蛰后不同时间牧草盲蝽的飞行能力,但不能确定其影响因素。因此,今后可针对温度、湿度、光照强度、日龄等因素进行研究,明确影响其飞行能力的主要因素。此外,本研究是在室内环境下测得数据,不能完全反映牧草盲蝽在自然条件下的飞行能力,因此应采取室内外多种方法进一步研究牧草盲蝽扩散与迁飞的规律,为牧草盲蝽越冬后的扩散区域预测和防控提供参考。
参考文献:
[1] 王伟,张仁福,刘海洋,等. 新疆喀什地区牧草盲蝽为害棉花防治指标研究[J]. 应用昆虫学报,2016,53(5):1146-
1152.
[2] 夏欣,郑艺翔,姚成层,等. 5种牧草盲蝽寄主植物挥发物鉴别、选择反应及油菜诱集带在棉田的诱集效果[J]. 棉花学报,2023,35(2):128-137.
[3] 曹娜,冷凌云,刘端春,等. 牧草盲蝽的寄主种类及取食选择性研究[J]. 中国棉花,2017,44(3):27-29,38.
[4] 张仁福,王伟,刘海洋,等. 棉区牧草盲蝽寄主种类及其寄主取食季节性演替规律[J]. 新疆农业科学,2022,59(3):707-715.
[5] Stewart S D,Gaylor M J. Effects of age,sex,and reproductive status on flight by the tarnished plant bug (Heteroptera:Miridae)[J]. Environmental Entomology,1994,23(1):80-
84.
[6] 陈豪,梁革梅,邹朗云,等. 昆虫抗寒性的研究进展[J]. 植物保护,2010,36(2):18-24.
[7] Hessien M M,Rashad M M,Zaky R R,et al. Controlling the synthesis conditions for silica nanosphere from semi-
burned rice straw[J]. Materials Science and Engineering: B,2009,162(1):14-21.
[8] 杨明超,杨涛. 牧草盲蝽在南疆的发生危害及其防治[J]. 植物保护,2001(5):31-32.
[9] 程登发,田喆,李红梅,等. 温度和湿度对麦长管蚜飞行能力的影响[J]. 昆虫学报,2002(1):80-85.
[10] 苏春芳,刘爱萍,高书晶,等. 温度和湿度对草地螟阿格姬蜂飞行能力的影响[J]. 中国生物防治学报,2014,30(5):612-617.
[11] 袁瑞玲,杨珊,冯丹,等. 温度、湿度、光照对桔小实蝇飞行能力的影响[J]. 环境昆虫学报,2016,38(5):903-911.
[12] 陆宴辉. 盲蝽蟓生态适应性研究[D]. 北京:中国农业科学院,2008.
[13] 赵金平. 牧草盲蝽滞育后生物学特性和种群动态研究[D]. 阿拉尔:塔里木大学,2021.
[14] 刘莎,吕召云,高欢欢,等. 昆虫飞行能力研究进展[J]. 环境昆虫学报,2018,40(5):995-1002.
[15] 程云霞. 草地螟Loxostege sticticalis迁飞与生殖行为的调控及互作关系[D]. 北京:中国农业科学院,2012.
[16] 侯国辉,崔笑雄,朱悦,等. 金纹细蛾飞行能力测定[J]. 昆虫学报,2022,65(5):612-620.
[17] Hashiyama A,Nomura M,Kurihara J,et al. Laboratory evaluation of the flight ability of female Autographa nigrisigna(Lepidoptera:Noctuidae),measured by actograph and flight mill[J]. Journal of Economic Entomology,2013,106(2):690-694.
[18] 潘攀,罗礼智,江幸福,等. 稻纵卷叶螟飞行行为特征[J]. 应用昆虫学报,2013,50(3):583-591.
(责任编辑:杨子山 责任校对:秦凡)
●
猜你喜欢
今日农业(2021年21期)2021-11-26 05:07:00
现代畜牧科技(2021年10期)2021-11-19 08:42:26
当代水产(2021年4期)2021-07-20 08:10:58
现代园艺(2017年11期)2017-06-28 11:32:51
现代园艺(2017年11期)2017-06-28 11:32:28
农家科技中旬版(2016年12期)2016-04-16 03:41:33
农家科技中旬版(2016年12期)2016-04-16 03:41:32
农家科技中旬版(2016年12期)2016-04-16 03:41:20
中国农业文摘-农业工程(2016年5期)2016-04-12 05:38:13
现代农业(2016年4期)2016-02-28 18:42:06
