
牛支原体贵州株黏附相关蛋白的初步鉴定
2024-05-07 12:39罗小芬谢晓东王永璇冉芳菲胡鹏飞朱二鹏程振涛
畜牧兽医学报 2024年4期
罗小芬,谢晓东,赵 超,胡 茜,王永璇,冉芳菲,胡鹏飞,文 明,2,朱二鹏,2*,程振涛,2*
(1.贵州大学动物科学学院,贵阳 550025, 2.贵州省动物疫病与兽医公共卫生重点实验室,贵阳 550025,3.盘州市动物疫病预防控制中心,盘州 553500)
支原体通常被认为是已知最小的自我复制生物,广泛分布在各种宿主中。1961年,美国首次分离出牛支原体(Mycoplasmabovis,Mb),可引起养殖场牛持续呼吸系统疾病、关节炎和腱鞘炎,以及犊牛乳腺炎、中耳炎和角膜结膜炎等[1-2]。尽管许多研究者都在努力探索其发病机制,但目前研究尚不完善。由于支原体无细胞壁,许多抗生素对其无效[3]。2008年,我国首次分离到该病原,随后我国多个省份陆续报道牛支原体感染牛肺炎病例[4]。近些年,本研究小组对贵州省规模养牛场(养殖小区)进行流行病学调查,发现疑似感染牛支原体肺炎,并成功分离得到两株牛支原体,分别命名Mycoplasmabovis-GZ-1、Mycoplasmabovis-GZ-2,表明贵州省存在牛支原体肺炎病例[5]。
许多研究表明支原体表面脂蛋白在感染和器官病变中起重要作用[6]。脂蛋白已被证明在细胞凋亡、ABC转运体操作、菌株毒力多样性和细胞黏附中发挥作用[7-12]。已有多项研究证明肺炎支原体的P1和P30蛋白[13],关节炎支原体的Maa1和Maa2蛋白[14],鸡毒原体(Mycoplasmagallisepticum, MG)的GapA和CrmA蛋白在细胞中有黏附作用[15]。也有研究在牛支原体中证明P26、α-烯醇化酶和Vsps家族的成员是黏附蛋白[16-17]。由于与宿主细胞的黏附是宿主定植和感染的先决条件,因此鉴定支原体中的黏附蛋白是了解其发病机制的重要前提。以上研究表明可以在牛支原体中鉴定黏附蛋白。然而关于Mycoplasmabovis-GZ-2是否存在其他黏附蛋白尚未报道。本研究旨在对Mycoplasmabovis-GZ-2的4个假定蛋白进行表达纯化、亚细胞定位和黏附性研究,为该蛋白的后续深入研究提供参考资料。
1 材料与方法
1.1 材料
1.1.1 实验动物 3月龄新西兰雌性大白兔10只购自贵阳经济技术开发区鹏程农业科技有限公司。
1.1.2 菌株与质粒Mycoplasmabovis-GZ-2[5]、重组pcold IM27、pcold IM32、pcold IM498、pcold IM663质粒、胚胎牛肺细胞(EBL)由贵州大学动物疫病研究所制备;大肠杆菌感受态细胞DE3(BL21)购自TaKaRa生物有限公司。
1.1.3 主要试剂 His标签蛋白纯化试剂盒、BCA蛋白定量试剂盒、羊抗鼠IgG-HRP、His标签单抗、聚丙烯酰胺凝胶试剂盒、5×SDS蛋白上样缓冲液均购于碧云天生物技术有限公司。Proteo-Extract Transmembrane Protein Extraction Kit购自上海贝博生物有限公司。激光共聚焦平皿购自Nest生物有限公司。异丙基-β-D-硫代半乳糖苷(IPTG)、Dil细胞膜染料,DAPI细胞核染料,胎牛血清,山羊抗兔FITC-IgG二抗,山羊抗兔HRP标记IgG抗体、弗氏佐剂购自Sigma生物有限公司。细胞培养皿、细胞刮购自Corning生物有限公司。Hanks平衡盐缓冲液购自Bio-Channel生物有限公司。2.5 g·L-1胰蛋白酶-EDTA消化液,双抗(10 000 U·mL-1青霉素,10 000 μg·mL-1链霉素),1 g·L-1Ⅰ型胶原酶购自Gibco生物有限公司。
1.2 重组蛋白的原核表达、鉴定及纯化
将保存的重组pcold IM27、pcold IM32、pcold IM498、pcold IM663质粒菌与空载体pCold Ⅰ 种子菌液复苏(37 ℃ 170 r·min-1)。按照20%的比例转接至新的200 mL LB液体培养基(含Amp+),OD600 nm达到0.4~0.6时,加入1 mmol·L-1的IPTG,37 ℃诱导表达目的蛋白3 h。全部菌液4 ℃ 8 500 r·min-1离心5 min。PBS洗3次,超声破碎20 min。4 ℃ 8 500 r·min-1离心30 min,分别取20 μL上清和沉淀进行SDS-PAGE分析。将上清与镍柱亲和纯化,收集5次洗脱液,进行SDS-PAGE分析。具体纯化操作步骤按His标签蛋白纯化试剂盒说明书进行。取20 μL上清进行Western blot鉴定,简要步骤:100 mA恒流湿转转膜3 h,将4个目的蛋白转至PVDF膜上,5%脱脂奶粉4 ℃封闭过夜,Anti-His tag鼠源单克隆抗体(1∶1 000)作为一抗,37 ℃ 孵育1 h,TBST洗3次,加入HRP标记羊抗鼠IgG (1∶1 000)酶标二抗,37 ℃孵育1 h,TBST洗涤3次。DAB底物显色溶液显色,红外扫描仪观察结果。
1.3 重组蛋白多克隆抗体的制备及效价的测定
将纯化后的M27、M32、M498、M663蛋白分别与弗氏完全佐剂等体积混合均匀充分乳化,按500 μg·只-1通过颈、背部多点皮下接种试验兔进行首次免疫(收集免疫前血清作为阴性对照),随后的两次免疫接种,用等量的弗氏不完全佐剂混合蛋白,每隔2周进行一次;第三次免疫后7 d,再补免一次,3 d后通过耳缘静脉采血测定抗体效价,当血清效价达到预期值,则通过心脏采血,制备血清,于-80 ℃冻存。将纯化后的4个重组蛋白分别用碳酸盐包被液稀释后, 以1 μg·mL-1的包被量包被酶标板,利用间接酶联免疫吸附试验 (ELISA) 分别检测4个抗体的效价。
1.4 牛支原体各组分蛋白的制备
取300 mL对数生长后期的牛支原体新鲜培养物,12 000 r·min-1离心30 min收集菌体,灭菌PBS洗涤3次,加入0.5 mL PBS进行超声破碎至清亮,得到的产物即为牛支原体总蛋白。按照Proteo-Extract Transmembrane Protein Extraction Kit说明书提取牛支原体的细胞膜和细胞质蛋白。简要步骤如下:按照上述方法收集对数生长后期的牛支原体,沉淀中加入200 μL预冷试剂A和2 μL蛋白酶抑制剂混匀,再加入2 μL稳定剂,高速涡旋振荡15 s,冰上10~15 min,再次高速涡旋振荡5 s,4 ℃ 8 200 r·min-1离心5 min。快速将上清吸入另一预冷的干净离心管,37 ℃水浴5~10 min,37 ℃ 1 200 r·min-1离心5 min,此时溶液分为两层,上层即为胞质蛋白,150~200 μL预冷试剂B稀释下层相,混匀,冰浴2 min,即为膜蛋白。
1.5 目的蛋白的亚细胞定位
分别取40 μL的牛支原体总蛋白、胞膜蛋白、胞浆蛋白以及4个表达蛋白,与5×蛋白上样缓冲液混合,100 ℃煮沸5 min,冷却至室温,8 000 r·min-1离心2 min,取20 μL上样,进行SDS-PAGE电泳和Western blot鉴定,操作步骤见方法“1.2”(一抗为兔抗M27、M32、M498、M663血清抗体)。
1.6 黏附与黏附抑制试验
1.6.1 激光共聚焦扫描显微镜分析 利用激光共聚焦显微镜分析M27、M32、M498、M663蛋白及牛支原体对EBL细胞的黏附影响,和目的蛋白多抗黏附抑制影响。具体步骤如下。
黏附试验:当激光共聚焦平皿中EBL细胞密度长至60%时,4%多聚甲醛固定,分别加入5 μL生长至对数期的牛支原体(1×109CFU·mL-1)和1 mL含20 μg目的蛋白的PBS,37 ℃,5% CO2培养箱中分别孵育4和2 h,经洗涤除去非黏附目的蛋白和牛支原体,未加蛋白组作为空白对照;黏附抑制试验:将目的蛋白和牛支原体分别与相应各多抗混合,37 ℃,5% CO2培养箱中孵育30 min,设立4个蛋白多抗混合组。取200 μL孵育混合物加到生长密度60%的EBL细胞中,37 ℃,5% CO2培养箱中孵育4 h。然后在含有细胞的平皿上覆盖兔抗牛支原体的阳性血清。用抗兔IgG-FITC检测与细胞结合的抗体。DiI标记细胞膜;DAPI标记细胞核。使用激光扫描共聚焦显微镜对免疫荧光进行评估。
1.6.2 流式细胞术分析 待EBL细胞密度在细胞培养皿中长至60%时,加入400 μL生长至对数期的牛支原体(1×109CFU·mL-1),37 ℃,5% CO2培养箱中孵育过夜,消化细胞转移至1.5 mL离心管中,1 000 r·min-1,离心5 min,弃上清,加入一抗、二抗进行孵育(一抗为兔抗牛支原体阳性血清,二抗为山羊抗兔FITC-IgG抗体,阴性对照组以兔阴性血清为一抗),经洗涤加入200 μL PBS重悬EBL细胞,加入等量4%多聚甲醛,过300目滤网后用流式细胞仪分析结果;空白对照组不加入牛支原体。黏附抑制试验中,牛支原体分别先与200 μL兔抗目的蛋白抗体37 ℃孵育30 min,再与EBL细胞黏附,其后具体操作同上。
2 结 果
2.1 重组蛋白的表达及纯化
重组pcold IM27、pcold IM32、pcold IM498、pcold IM663质粒表达菌(DE3),经ITPG诱导,进行SDS-PAGE分析。图1结果显示:重组M27、M32、M498、M663蛋白成功表达,大部分存在上清,为可溶性蛋白表达。以His单克隆抗体和兔抗牛支原体阳性血清为一抗对4个重组蛋白进行Western blot鉴定,图2结果显示:存在4个重组蛋白的目的条带,蛋白分子量大小为27、16、19和26 ku。在非变性条件下,用His标签蛋白纯化试剂盒进行纯化,图3结果显示:利用Image J软件对SDS-PAGE凝胶电泳条带进行强度分析,成功纯化得到4个重组蛋白。
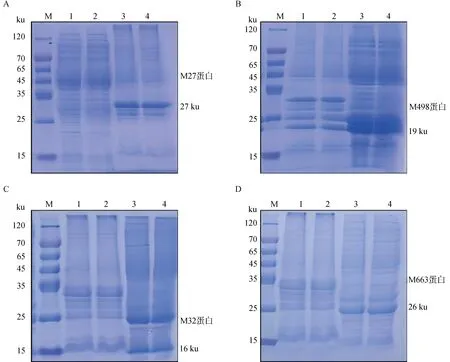
A. M27;B. M498;C. M32;D. M663;M. 蛋白质相对分子质量标准;1、2. 超声裂解沉淀;3、4. 超声裂解上清A. M27; B. M498; C. M32; D. M663; M. Protein marker; 1, 2. Lysate precipitation from ultrasonic lysis; 3, 4. Lysate supernatant from ultrasonic lysis图1 M27、M32、M498、M663重组蛋白SDS-PAGE分析Fig.1 SDS-PAGE analysis of M27, M32, M498, M663 recombinant proteins

A~D. 以His单克隆抗体为一抗的Western blot;E~H. 以 兔抗牛支原体阳性血清为一抗的Western blot;A、E. M27;B、F. M498;C、G. M32;D、H. M663;1. pcold I空载体对照;2. 表达蛋白;M. 蛋白质相对分子质量标准A-D. Western blot detection using His monoclonal antibody as the primary antibody; E-H. Western blot detection using rabbit anti Mycoplasma bovis positive serum as the primary antibody; A, E. M27; B, F. M498; C, G. M32; D, H. M663; 1. PCold I empty carrier control; 2. Expressed proteins; M. Protein marker图2 M27、M32、M498、M663表达蛋白的Western blot分析Fig.2 Western blot analysis of M27, M32, M498, and M663 expressed proteins

A. M27;B. M498;C. M32;D. M663;M. 蛋白质相对分子质量标准;1. 蛋白穿流液;2、3. 蛋白洗涤液;4~9. 洗脱蛋白A. M27; B. M498; C. M32; D. M663; M. Protein marker; 1. Protein transmembrane fluid; 2, 3. Protein washing solution; 4-9. Elution protein图3 纯化后M27、M32、M498、M663蛋白的SDS-PAGE分析Fig.3 SDS-PAGE analysis of purified M27, M32, M498, and M663 proteins
2.2 重组蛋白多克隆抗体ELISA效价测定
经酶联免疫吸附试验(ELISA)测定兔抗M27、M32、M498、M663蛋白血清抗体。结果显示:兔抗M27、M32、M663蛋白高免血清抗体效价高达1∶204 800倍,兔抗M498蛋白高免血清抗体效价达1∶25 600(免疫组/对照组>2.1判为阳性)。
2.3 重组蛋白的亚细胞定位
利用Western blot分析M27、M32、M498、M663蛋白在牛支原体中的亚细胞定位,兔抗M27、M32、M498、M663高免血清为一抗,山羊抗兔HRP标记IgG抗体为二抗,结果如图4所示,结果显示:M27、M32、M498均在牛支原体总蛋白、胞膜蛋白与胞质蛋白中均出现特异印迹条带,说明M27、M32、M498蛋白同时存在牛支原体的胞膜组分与细胞质组分中。然而,M663在牛支原体提取的细胞膜部分蛋白、细胞质部分蛋白和牛支原体总蛋白中没有检测到目的条带。

A. M27;B. M498;C. M32;D. M663;1. 表达蛋白;2. 牛支原体总蛋白;3. 胞膜蛋白;4. 胞质蛋白A. M27; B. M498; C. M32; D. M663; 1. Expressed proteins; 2. Total protein of Mycoplasma bovis; 3. Membrane proteins; 4. Cytoplasmic proteins图4 4个重组蛋白的亚细胞定位Fig.4 Subcellular localization of the four recombinant proteins
2.4 激光共聚焦扫描显微镜分析
利用激光共聚焦扫描显微镜分析M27、M32、M498、M663蛋白(图5~7)及牛支原体对EBL细胞的黏附试验和兔抗M27、M32、M498、M663多克隆抗体抑制相应蛋白对EBL细胞的黏附抑制试验,以及单独和联合作用抑制牛支原体对EBL细胞的黏附抑制试验(M27、M32、M498、M663蛋白多抗、牛支原体阳性血清为一抗,山羊抗兔FITC-IgG为二抗),DAPI染色后细胞核呈蓝色 (EBL细胞),Dil染色后细胞膜呈红色,绿色则为FITC-IgG荧光标记二抗显色效果及Image J软件分析绿色荧光强度。图5和图8A结果显示:牛支原体、M27、M32、M498、M663均能黏附在EBL细胞,图6和图8B结果显示:兔抗M27、M32、M498、M663血清抗体能够明显的抑制蛋白M27、M32、M498、M663对EBL细胞的黏附:图7和图8C结果显示:在牛支原体黏附抑制试验中,通过与牛支原体正常组对照可得,兔抗M27、M32、M498、M663血清抗体能够抑制牛支原体黏附EBL细胞,4个蛋白兔抗多克隆抗体联合对抑制牛支原体黏附EBL细胞的作用比较为明显,但是牛支原体不是只有这4种黏附蛋白,还有其他黏附蛋白存在。
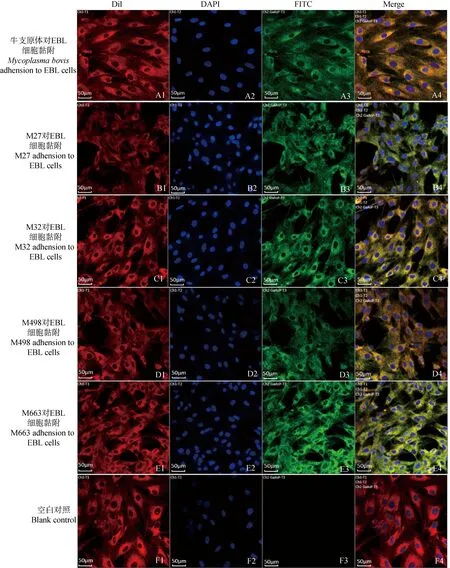
图5 激光共聚焦观察牛支原体、M27、M32、M498、M663对EBL细胞黏附Fig.5 Laser confocal observation of Mycoplasma bovis, M27, M32, M498, M663 adhesion to EBL cells

图6 激光共聚焦观察目的蛋白抗血清对M27、M32、M498、M663黏附EBL细胞抑制Fig.6 Laser confocal observation of inhibition of M27, M32, M498, M663 adhesion to EBL cells by target protein antisera

图7 激光共聚焦观察目的蛋白抗血清对牛支原体黏附EBL细胞抑制Fig.7 Laser confocal observation of inhibition of Mycoplasma bovis adhesion to EBL cells by target protein antisera
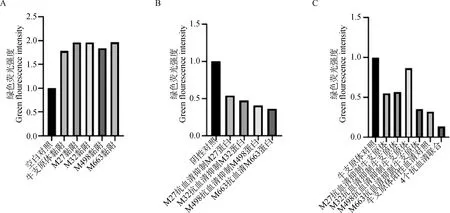
A. 牛支原体、M27、M32、M498、M663对EBL细胞黏附的绿色荧光强度;B. M27、M32、M498、M663蛋白抗血清抑制M27、M32、M498、M663黏附EBL细胞的绿色荧光强度;C. M27、M32、M498、M663蛋白抗血清抑制牛支原体黏附EBL细胞的绿色荧光强度A. The green fluorescence intensity of Mycoplasma bovis, M27, M32, M498, and M663 on EBL cell adhesion; B. The anti-serum of M27, M32, M498, and M663 proteins inhibits the green fluorescence intensity of M27, M32, M498, and M663 adhering to EBL cells; C. M27, M32, M498, M663 protein antiserum inhibits the green fluorescence intensity of Mycoplasma bovis adhesion to EBL cells图8 免疫荧光-绿色荧光强度分析Fig.8 Immunofluorescence-green fluorescence intensity analysis
2.5 流式细胞术分析
利用流式细胞仪检测牛支原体对EBL细胞的黏附试验以及兔抗M27、M32、M498、M663多克隆抗体单独和联合抑制牛支原体对EBL细胞的黏附抑制试验。图9结果显示,与空白对照、阴性对照相比,牛支原体对EBL细胞的黏附率为59.6%,抗M27、M32、M498、M663蛋白血清抑制率分别为59.6%、59.5%、59.6%、33.3%,4个抗蛋白血清联合作用牛支原体黏附EBL细胞时,抑制率为53.1%,牛支原体阳性血清的抑制率为51.8%,表明4个蛋白抗血清均能抑制牛支原体对EBL细胞的黏附,但是牛支原体不是只有这4种黏附蛋白,还有其他黏附蛋白存在。

(图9续 Continued)A. 牛支原体对EBL细胞的黏附;B. 抗M27血清对牛支原体黏附EBL细胞的抑制;C. 抗M32血清对牛支原体黏附EBL细胞的抑制;D. 抗M498血清对牛支原体黏附EBL细胞的抑制;E. 抗M663血清对牛支原体黏附EBL细胞的抑制;F. 抗M27、M32、M498、M663血清联合作用对牛支原体黏附EBL细胞的抑制;G. 牛支原体阳性血清对牛支原体黏附EBL细胞的抑制;H. 空白对照;I. 阴性对照A. Adhesion of Mycoplasma bovis to EBL cells; B. Inhibition of Mycoplasma bovis adhesion to EBL cells by anti M27 serum; C. Inhibition of Mycoplasma bovis adhesion to EBL cells by anti M32 serum; D. Inhibition of Mycoplasma bovis adhesion to EBL cells by anti M498 serum; E. Inhibition of Mycoplasma bovis adhesion to EBL cells by anti M663 serum; F. The combined effect of anti M27, M32, M498, and M663 serum on the adhesion of Mycoplasma bovis to EBL cells; G. Inhibition of Mycoplasma bovis positive serum on Mycoplasma bovis adhesion to EBL cells; H. Blank control; I. Negative control图9 牛支原体对EBL细胞的黏附与抗M27、M32、M498、M663血清单独以及联合作用对该黏附的抑制Fig.9 Adhesion of Mycoplasma bovis to EBL cells and inhibition of this adhesion by anti-M27, M32, M498, and M663 sera alone and in combination
3 讨 论
牛支原体是引起饲养场奶牛和犊牛慢性肺炎的重要病原体[18]。由于抗生素治疗的结果不太理想,这种原核生物给全世界的养牛业造成了巨大的经济损失[19]。牛支原体被发现至今已有60余年,但其致病和免疫机制研究目前尚不完善[20]。牛支原体无细胞壁,直接与宿主细胞相互作用,获取营养物质依赖于许多特征不明的膜相关蛋白。由于对易感宿主细胞的黏附是许多细菌定植和感染的先决条件,因此目前的研究主要集中在支原体的膜蛋白黏附成分上。迄今为止,已有多种牛支原体的相关黏附蛋白获得鉴定,如P27[21]、MBOV_0503[22]、mbfN[23]等黏附有关的膜蛋白。这些发现对加强牛支原体对宿主细胞的黏附能力及阐明牛支原体的毒力因素和致病机制有一定的影响。
有研究通过免疫蛋白组学方法鉴定出相关膜蛋白具有很好免疫原性,探索蛋白在牛支原体细胞中的分布及其是否具有免疫原性,对其克隆表达是基础[24-26]。也有研究利用Western blot成功鉴定鸡毒支原体rP25、rP33膜蛋白,并证实其具有黏附性[27];还有研究也利用Western blot成功鉴定牛支原体NOX2蛋白为牛支原体膜蛋白组成成分[28]。综上所述,原核表达及纯化,操作技术简单且易获得大量蛋白、Western blot进行膜定位、激光共聚焦和流式细胞术进行黏附性分析,鉴定技术传统且操作简单,在一定程度上增加了数据结果的科学性和展示的直观性。因此,本研究通过对牛支原体贵州株全基因组中的假定蛋白进行原核表达及纯化、亚细胞定位和黏附性研究,发现M27、M32和M498蛋白免疫原性良好,其特异性血清抗体能与牛支原体膜蛋白成分发生特异性反应,均为牛支原体黏附相关蛋白。但M663蛋白血清抗体虽未与牛支原体膜蛋白发生特异性反应,M663蛋白及其血清抗体仍能够黏附EBL细胞和抑制牛支原体对EBL细胞的黏附。本研究经过反复试验,结果也是如此,目前可能原因是M663蛋白在牛支原体表达量较低,不足以达到Western blot 检测所需含量,可通过该蛋白的过表达来进一步验证其蛋白属性,但具体原因尚且不知,需进一步试验验证。此外,流式细胞仪观察结果和激光共聚焦观察结果均显示M498、M663蛋白均能黏附EBL细胞,对牛支原体有一定的抑制作用。其中M663蛋白膜定位的Western blot 检测呈阴性,但仍具有抑制支原体的黏附性,可能是因为Western blot与免疫荧光试验和流式细胞术中的抗原-抗体反应特性和灵敏度有差异。在牛支原体膜蛋白黏附研究中,单一蛋白可能并不发挥黏附作用或不会影响牛支原体的黏附,而是通过多种蛋白协同作用才发挥黏附作用和影响牛支原体黏附功能;M663蛋白也可能作为一种协同蛋白参与牛支原体黏附[29]。本文对M27、M32、M498、M663蛋白的功能初步验证,为后期研究该蛋白的深入研究奠定一定的基础及防控牛支原体贵州株提供了科学依据。
4 结 论
本研究表明,M27、M32 、M498是牛支原体的黏附相关蛋白。虽然M663在Western blot鉴定中未检测到,但具有黏附性,且其抗血清能够抑制牛支原体对EBL细胞的黏附,这一发现值得深入研究,并可能为了解牛支原体黏附宿主细胞的作用机制提供科学依据。
猜你喜欢
兽医导刊(2019年1期)2019-02-21
兽医导刊(2019年1期)2019-02-21
妈妈宝宝(2017年3期)2017-02-21
哈尔滨医药(2016年3期)2016-12-01
兽医导刊(2016年12期)2016-05-17
兽医导刊(2016年12期)2016-05-17
肝博士(2015年2期)2015-02-27
癌变·畸变·突变(2015年4期)2015-02-27
现代检验医学杂志(2015年4期)2015-02-06
现代检验医学杂志(2015年1期)2015-02-06
