
Patterns and drivers of plant sexual systems in the dry-hot valley region of southwestern China
2024-04-12 02:03:58RongQiXuYongqinGoDeliPengHngSunBoSong
植物多样性 2024年2期
Rong M ,Qi Xu ,Yongqin Go ,Deli Peng ,Hng Sun ,** ,Bo Song ,***
a State Key Laboratory of Plant Diversity and Specialty Crops/Yunnan Key Laboratory for Integrative Conservation of Plant Species with Extremely Small Populations,Kunming Institute of Botany,Chinese Academy of Sciences,Kunming 650201,China
b College of Life Sciences,Shaanxi Normal University,Xi'an 710119,China
c Yunnan Forestry Technological College,Kunming 650224,China
d School of Life Science/Key Laboratory of Yunnan for Biomass Energy and Biotechnology of Environment,Yunnan Normal University,Kunming 650500,Yunnan,China
Keywords: Floral traits Fruit type Growth form Pollination system Reproductive phenology
ABSTRACT Sexual systems play important roles in angiosperm evolution and exhibit substantial variations among different floras.Thus,studying their evolution in a whole flora is crucial for understanding the formation and maintenance of plant biodiversity and predicting its responses to environmental change.In this study,we determined the patterns of plant sexual systems and their associations with geographic elements and various life-history traits in dry-hot valley region of southwestern China,an extremely vulnerable ecosystem.Of the 3166 angiosperm species recorded in this area,74.5%were hermaphroditic,13.5%were monoecious and 12%were dioecious,showing a high incidence of diclinous species.Diclinous species were strongly associated with tropical elements,whereas hermaphroditic species were strongly associated with temperate and cosmopolitan elements.We also found that hermaphroditism was strongly associated with showy floral displays,specialist entomophily,dry fruits and herbaceous plants.Dioecy was strongly associated with inconspicuous,pale-colored flowers,generalist entomophily,fleshy fruits,and woody plants,whereas monoecy was strongly associated with inconspicuous,pale-colored flowers,anemophily,dry fruits,and herbaceous plants.In addition,hermaphroditic species with generalist entomophily tended to flower in the dry season,whereas diclinous species with specialist entomophily tended to flower in the rainy season.However,independent of sexual systems,plants that produce dry fruits tended to flower in the rainy season and set fruits in the dry season,but the opposite pattern was found for fleshy fruit-producing plants.Our results suggest that in the dry-hot valleys,plant sexual systems are associated with geographic elements as well as various life-history traits that are sensitive to environmental change.
1.Introduction
Angiosperms exhibit an amazing diversity of reproductive traits to ensure reproductive success (Lloyd and Barrett,1996),by adapting to various selective agents,including pollinators,biotic non-pollinators and abiotic factors (Strauss and Whittall,2006;Song et al.,2013).Of all these reproductive traits,sexual systems have long been considered to be one of the most important traits in influencing the genetic,evolutionary dynamics and diversification of angiosperms and thus have been a central topic in evolutionary biology for hundreds of years (Barrett,2010;Schoen et al.,2019).
Although angiosperms are mainly hermaphroditic (Renner and Ricklefs,1995),substantial variation in the frequency of sexual system (hermaphroditic,monoecious and dioecious) has been reported among different geographic regions or floras (Fox,1985;Wang et al.,2020,2021).Furthermore,several studies have suggested that this variation in plant sexual system is driven by not only external climatic factors (e.g.,temperature and precipitation)but also the intrinsic plant life-history traits(e.g.,growth form,fruit type,and reproductive phenology) (Gross,2005;Moeller et al.,2017;Wang et al.,2021;Ramírez and Brice~no,2022).
Previous studies have shown that geographical variation in the composition of sexual systems of local floras may be caused by variation in local temperature and precipitation (Hultine et al.,2016;Wang et al.,2021).However,the effect of these environmental factors on the sexual systems of plants has not been uniform.For example,high temperature has been shown to aggravate male-biased sex ratios due to sexual dimorphism in resource acquisition and allocation (Tognetti,2012;Etterson and Mazer,2016;Hultine et al.,2016).Consequently,dioecious species in regions with high temperature are generally at risk(Etterson and Mazer,2016;Wang et al.,2021).These findings indicate that dioecy should be less common in regions with high temperatures,whereas hermaphroditism should be more common in warm and arid regions.This hypothesis has been supported by findings that dioecy in local floras increases with latitude (Godin,2017;Kevan and Godglick,2017;Wang et al.,2021) and that many dioecious species are thought to be more resistant to cold stress than other sexual systems (Lithgow and Kirkwood,1996).Some studies,however,have not supported the hypothesis that plant sexual systems are simply related to temperature.For example,Peng et al.(2014) found that the frequency of dioecious species was particularly low in cold alpine subnival belt of the Hengduan Mountains,while the frequency of hermaphroditic species was particularly high.This result is likely because compared with diclinous species,hermaphroditic species can provide reproductive assurance by selfing.In addition,several studies have shown that dioecy is more common in humid and tropical climate regions (Ruiz-Zapata and Arroyo,1978;Bullock,1985;Sakai et al.,1995;Gross,2005;Chen and Li,2008b;Vary et al.,2011).Importantly,the positive correlation between dioecy and tropical environments may be attributed to growth form.Specifically,reports have shown that dioecy is strongly associated with woody plants,which are predominate in tropical climates(Bawa,1980;Givnish,1980),whilst hermaphroditism is associated with herbaceous plants,which are predominant in temperate climates (Vamosi et al.,2003;Moeller et al.,2017).Therefore,when considering how sexual system composition varies geographically,we should not only consider climate factors,but also plant life-history traits(e.g.,Wang et al.,2020).
In recent decades,studies on the correlation between plant sexual system and life-history traits have emerged in various regions at the community level (e.g.,Chen and Li,2008a,b;Vary et al.,2011;Peng et al.,2014;Godin,2017;Ramírez and Brice~no,2022).Many authors have found that hermaphroditism is strongly associated with showy floral displays,specialist entomophily,perennial forbs,and dry fruits;whereas dioecy is strongly associated with non-specialized pollination vectors,small and pale-colored flowers,woody plants,and fleshy fruits(reviewed in Renner and Ricklefs,1995;Sakai et al.,1995;Matallana et al.,2005;Peng et al.,2014;Vamosi and Queenborough,2010;Godin,2017;Ohya et al.,2017).However,studies on the correlations between sexual system and plant lifehistory traits have mainly concentrated on hot and humid tropical climate regions(e.g.,Gross,2005;Matallana et al.,2005;Chen and Li,2008b;but see Godin,2017;Xia et al.,2020),and some floras with extreme high temperature and drought conditions,remain to be studied.
Our goal in this study is to determine the patterns of sexual systems and their correlates in the dry-hot valley (hereafter DHV)region of southwestern China.The DHV region is distributed mainly in Yunnan and Sichuan Province in the deep,hot,and dry valleys of several big rivers.It has a mountainous topography with the mountain ridges generally running in a north-south direction,decreasing in elevation southward.The DHV region is the lowest vertical zone in the mountain ecosystems,and is characterized by tropical (hot) dry climates due to the “foehn effect” (Zhu et al.,2020).This region has distinct wet and dry seasons (He et al.,2000).The flora in the DHV is a well-documented subset of the whole flora,comprising herbaceous plants (grasses) and dotted with sparse trees or shrubs(Jin,1986).Despite its harsh biotic and abiotic environmental conditions,the DHV hosts remarkable high plant diversity;in total,3219 seed plant species and varieties from 1038 genera in 163 families have been recorded from the DHV(Jin,2002;Zhu et al.,2020).In addition,several important conservation areas have been established in this area due to its high plant diversity and a particularly high proportion of endemic plant species(Li et al.,2011;Tang et al.,2020;Zhu and Du,2022).Several studies have indicated that the flora of the DHV is marginally of tropical and has been influenced by temperate elements (Zhu,2014;Zhu et al.,2020).Here,we hypothesized that in the DHV region of southwestern China: (1) diclinous species are more common in tropical elements and hermaphroditic species are more common in temperate elements;and (2) diclinous species may be correlated with some advantageous reproductive traits that allow diclinous plants to adapt to fewer pollinators and distinct dry and rainy season.We further hypothesized that sexual systems and various reproductive traits show seasonal variations,i.e.,some reproductive traits are adapted to the dry season and other reproductive traits are adapted to the rainy season.
To test the above hypotheses,we used the latest and most comprehensive plant checklist to identify sexual systems of species in the DHV region and then checked for associations between sexual systems and geographical elements and various morphological and ecological traits (Jin,2002;Zhu et al.,2020).Specifically,we had four main objectives: (1) to determine the frequency of hermaphroditic,monoecious and dioecious sexual systems;(2) to compare the frequency of sexual systems in different geographical elements (e.g.,cosmopolitan,tropical and temperate elements);(3) to determine the correlations of sexual systems with other morphological and ecological traits;(4) to explore the seasonal patterns in the occurrence of sexual systems and their correlates.
2.Materials and methods
2.1.Study area
The DHV region of southwestern China,which was formed by several rivers,is a well-known natural landscape.Our study focused on the flora of four major river valleys (i.e.,Jinshajiang River,Lancangjiang River,Nujiang River and Yuanjiang River).The river valleys range between 23°-28°10′N and 98°50′-103°50′E(Zhu et al.,2020),spanning the river valley region of western,northwestern and southern Yunnan,and western Sichuan (Fig.1).The DHV region is characterized by high temperature (21-24°C),low annual rainfall (600-800 mm),and high evaporation(2750-3850 mm) (Zhang,1992).Because of the special geographical position,complex geological history and unique ecological environments,the DHV region hosts abundant biodiversity (Zhu,2014;Zhu et al.,2020),but the ecosystem is extremely vulnerable.
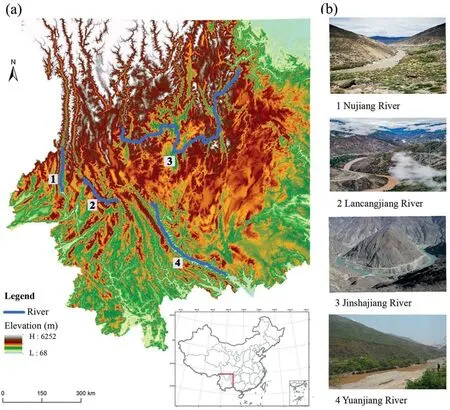
Fig.1.The geographical range of our study.Location of the study area in China and main rivers(a);Landscape of Nujiang River,Lancangjiang River,Jinshajiang River and Yuanjiang River (b).
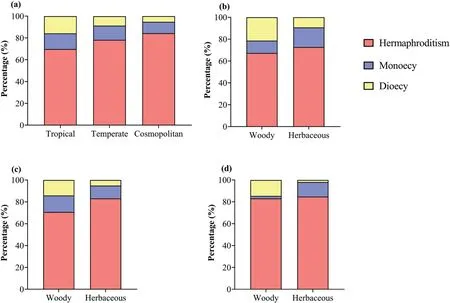
Fig.2.Distribution of hermaphroditism,monoecy and dioecy in different geographical elements (a) and the distribution of sexual systems in different growth forms in each geographical elements (b-d): tropical elements (b),temperate elements (c),and cosmopolitan (d).
2.2.Data collection
The plant species used for this study were derived from the checklist of angiosperms from the four valleys in southwestern China(Jin,2002;Zhu et al.,2020).In total,3166 species belonging to 1059 genera and 162 families were included in our final dataset.We collected information on plant geographical elements and various life-history traits from theFlora of China(Wu et al.,1994),China Virtual Botanical Herbarium(http://www.cvh.ac.cn),specimens(herbarium KUN),published literature,field observation,and communication with experts.Plant geographical elements and various life-history traits considered in this study are listed in Table 1.All species were classified into three categories according to their sexual system following Wang et al.(2020): hermaphroditic,dioecious or monoecious.Dioecious plant species include androdioecious,gynodioecious,and polygamodioecious species,while monoecious plant species include monoecious,andromonecious,and gynomouoecious species.We divided geographical elements into three categories according to Wu (1991) and Wu et al.(2006):cosmopolitan,tropical or temperate elements.We classified growth form as herbaceous or woody.Followed the methods of Gross(2005),Chen and Li (2008b) and Peng et al.(2014),pollination syndrome,floral clustering,flower color,flower size,floral symmetry and fruit type for each species were classified.Flowering time and fruiting time for each species were classified as rainy season (from May to October) or dry season (from October to May).In addition,given that clonal reproduction has been found to be an important reproductive assurance for plants in adverse environments (Gao et al.,2020),we also investigated whether a species is capable of producing new and potentially independent ramets through clonal reproduction based on the morphological description inFlora of Chinafollowing Zhang et al.(2018)and Gao et al.(2020)
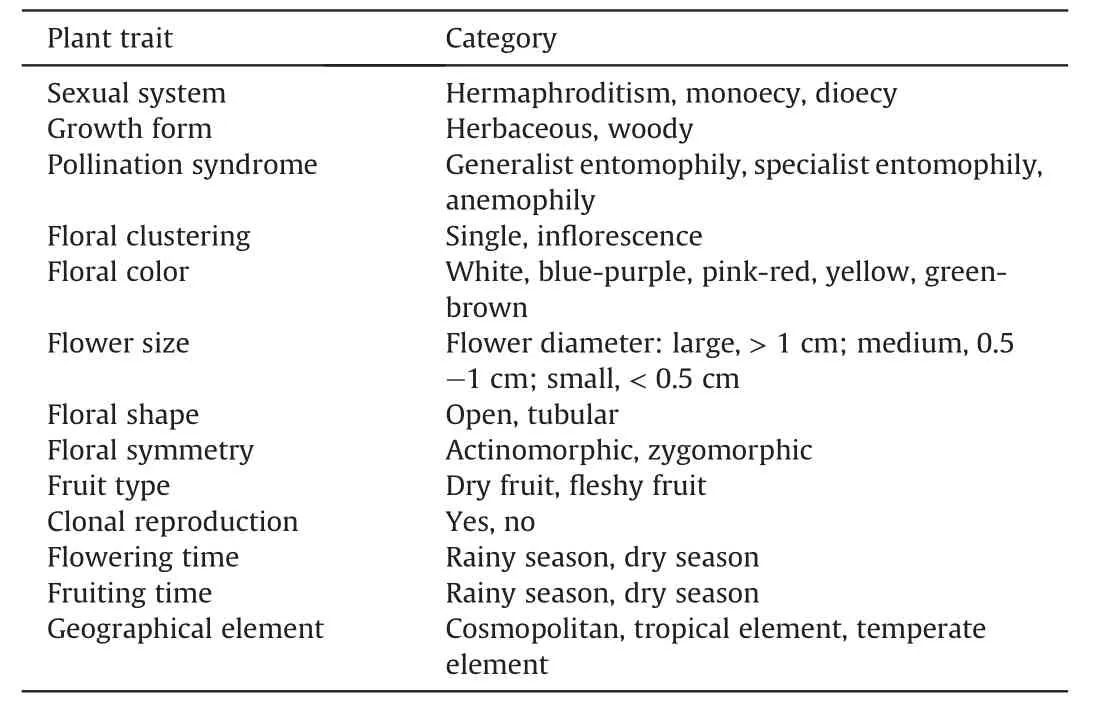
Table 1 Plant traits and their categories in this study.
2.3.Data analysis
Chi-square test (χ2) was used to identify correlations between sexual system and geographical element type,growth form,fruit type,pollination syndrome,clonal reproduction and various floral traits.We also analyzed associations between flowering/fruiting time(dry and rainy season)and reproductive traits of plants usingchi-square test.In the chi-square test,the observed frequency was the actual one obtained from investigations and the expected frequency was a theoretically predicted by statistical software.Our null hypothesis was that no difference would exist between groups.The ratio of the observed value to the expected value was compared with 1.A value of the ratio >1 indicates that the specific sexual system (or other plant trait) is overrepresented for the trait group under the assumption that the sexual systems and geographical elements and morphological or ecological traits are statistically independent.All statistical analyses were performed in SPSS(version 26,SPSS,Chicago,IL,USA).Graphs were plotted by GraphPad Prism (v.8.0.2).
3.Results
3.1.Sexual systems and geographical elements
Of the 3166 species recorded in the DHV region,74.5% (2359)were hermaphroditic,13.5%(426)were monoecious,and 12%(381)were dioecious.Of the 1059 genera recorded in our dataset,74.5%(789)were hermaphroditic,13.4%(142)were monoecious,and 9.1%(96)were dioecious.The remaining 3%(32)had two or three types of sexual systems.Of the 162 families,59.3% (96) were exclusively hermaphroditic,6.8%(11)were exclusively monoecious and 8%(13)were exclusively dioecious.The remaining 25.9% (42) had at least two kinds of sexual systems.
Among the 1059 genera,58.2% (616) genera were tropical,35.6% (377) were temperate,and 6.2% (66) were cosmopolitan.Among the 3166 species,51.3% (1623) were classified as tropical,37.8% (1198) were temperate,and 10.9% (345)were cosmopolitan.Sexual systems were strongly related to geographical elements (Figs.2 and 3).The proportion of diclinous species were higher in tropical elements than in temperate elements or cosmopolitan elements;in addition,hermaphroditic species were more frequent in cosmopolitan and temperate elements than in tropical elements (Figs.2 and 3).When each dry-hot valley was considered separately,linear regression analyses showed that the proportion of hermaphroditic species was negatively correlated with the proportion of tropical elements and positively correlated with temperate elements;in contrast,the proportion of diclinous species were positively correlate with tropical elements and negatively correlated with temperate elements (Fig.S1).
Similar to other savanna plant communities,in the DHV region hermaphroditic species were dominant,with a relatively higher proportion of dioecious species (Table 2).When only tropical elements were considered,the patterns of sexual systems in the DHV were similar to those of tropical plant communities;similarly,when only temperate elements were considered,the patterns of sexual systems in the DHV were similar to those of temperature plant communities (Table 2).
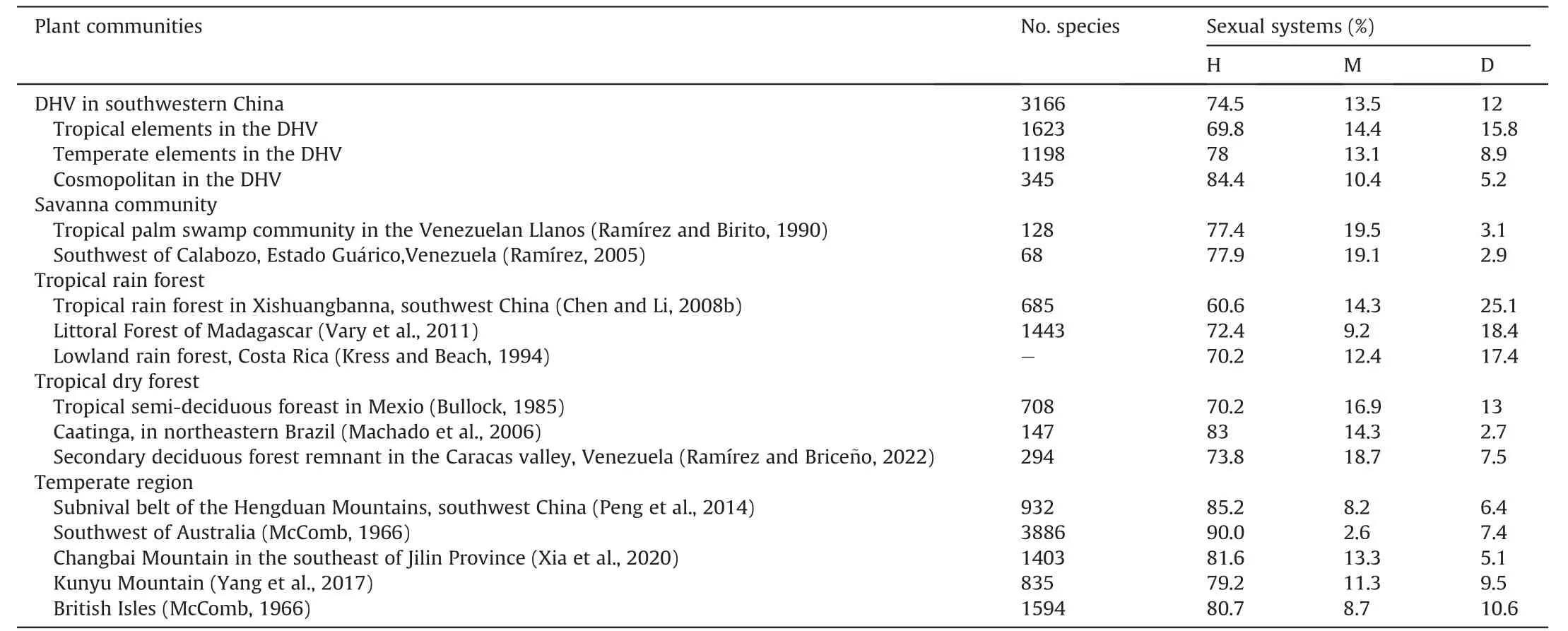
Table 2 Comparison of frequency of sexual systems of the dry-hot valley(DHV) region of southwestern China to that of other floras.
3.2.Sexual systems,growth form and clonal reproduction
Herbaceous species represented 53.8% (1703) and woody species represented 46.2% (1463) of the flora.Sexual systems were significantly related to growth forms,especially for diclinous sexual system (Fig.4).We found that monoecy was more represented in herbaceous plants and less represented in woody plants,and dioecy was more represented in woody plants and less represented in herbaceous plants (Fig.4).These results were maintained in analyses using the dataset including only tropical elements(Fig.3b).
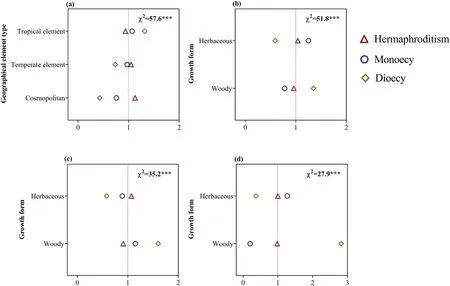
Fig.3.Ratio of observed and expected abundance of different sexual systems in different geographical element types (a) and in different growth forms in tropical elements (b),temperate elements (c),and cosmopolitan (d) in the dry-hot valley region of southwestern China.The value of the ratio >1 indicates that the specific sexual system is overrepresented for the trait group under the assumption that sexual system and geographical element or growth form are statistically independent.χ2 indicates the reliability of the differences;***P <0.001;0-2 (or 3) are the ratio of observed and expected abundances of species.
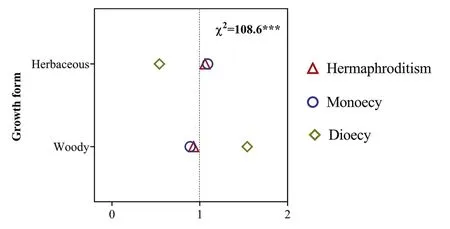
Fig.4.Ratio of observed and expected abundance of different sexual systems in different growth forms in the dry-hot valley region of southwestern China.The value of the ratio >1 indicates that the specific sexual system is overrepresented for the trait group under the assumption that the sexual system and growth form are statistically independent.χ2 indicates the reliability of the differences;***P <0.001;0-2 are the ratio of observed and expected abundances of species.
Of the 3166 species,24.2% (765) were clonal,and 75.8% (2401)were non-clonal.Sexual systems significantly correlated with clonal reproduction (Fig.5a): within species with clonal reproduction,the diclinous species account for the higher proportion than expected.We also found that clonal reproduction significantly correlated with growth forms.Clonal reproduction was more represented in herbaceous plants,whereas non-clonal reproduction was more represented in woody plants (Fig.5b).
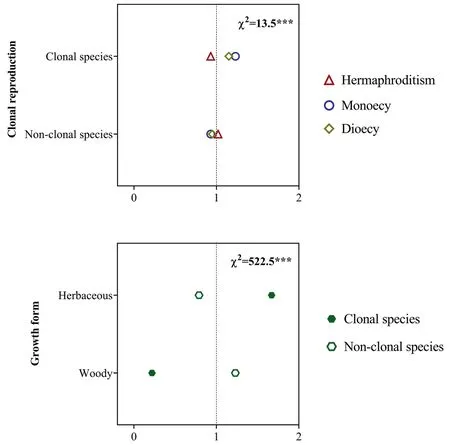
Fig.5.Ratio of observed and expected abundance of different sexual systems in plants with clonal reproduction and without clonal reproduction (a) and of clonal reproduction types in different growth forms (b) in the dry-hot valley region of southwestern China.The value of the ratio >1 indicates that the specific sexual system or clonality are overrepresented for the trait group under the assumption that sexual system and clonal reproduction,and clonal reproduction and growth form are statistically independent.χ2 indicates the reliability of the differences;***P <0.001;0-2 are the ratio of observed and expected abundances of species.
3.3.Sexual systems,flower morphology and flowering time
In the DHV region,91.4% (2893) of plant species produced inflorescences;the remaining 8.6% (273) produced solitary flower.The highest proportion of species had yellow flowers (28.5%;903 species),followed by white flowers (26.7%;845 species),greenbrown flowers (17.6%;557 species) and blue-purple flowers(15.4%;487 species).The fewest species had pink-red flowers(11.8%;374 species).In total,42.2%(1337)of plant species had large flowers,21.6% (683) medium flowers and 36.2% (1146) small flowers.A total of 76.8% (2433) of plant species produced actinomorphic flowers;the remaining 23.2% (733) produced zygomorphic flowers.The highest proportion (53.4%) of plant species were pollinated through generalist entomophily,followed by specialist entomophily (31.7%) and anemophily (14.9%).Sexual systems showed significant associations with floral morphology and pollination syndrome (Fig.6);moreover,these relationships varied between the dry and rainy seasons.Specifically,plants with generalist floral morphology (e.g.,small and inconspicuous color)often blossomed in dry season (Fig.7),and the relationship was stronger in hermaphroditic plants (Fig.8a).Plants with specialist floral morphology (e.g.,large and showy color) usually blossomed in rainy season (Fig.7),and the relationship was stronger in diclinous plants (Fig.8b).
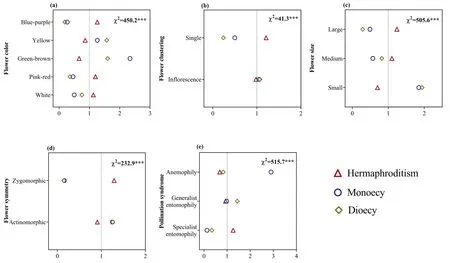
Fig.6.Ratio of observed and expected abundance of different sexual systems in different floral morphologies (a-d) and pollination syndromes (e) in the dry-hot valley region of southwestern China.The value of the ratio >1 indicates that the specific sexual system is overrepresented for the trait group under the assumption that sexual system and the morphology trait and pollination syndrome are statistically independent.χ2 indicates the reliability of the differences;***P <0.001;0-2 (3 or 4) are the ratio of observed and expected abundances of species.
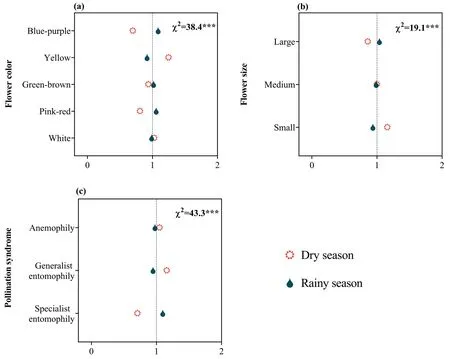
Fig.7.Ratio of observed and expected abundance of different flowering time in different flower colors(a),flower sizes(b)and pollination syndromes(c)in the dry-hot valley region of southwestern China.The value of the ratio >1 indicates that the specific flowering time is overrepresented for the trait group under the assumption that flowering time and morphology trait or pollination syndrome are statistically independent.χ2 indicates the reliability of the differences;***P <0.001;0-2 are the ratio of observed and expected abundances of species.
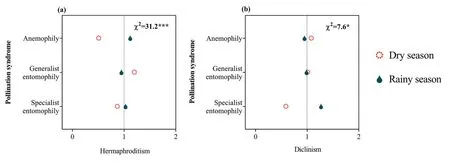
Fig.8.Ratio of observed and expected abundance of different flowering time in different pollination syndromes for hermaphroditism (a) and diclinism (b) in the dry-hot valley region of southwestern China.The value of the ratio>1 indicates that the specific flowering time is overrepresented for the trait group under the assumption that flowering time and pollination syndrome are statistically independent.χ2 indicates the reliability of the differences;***P <0.001;*P <0.05;0-2 are the ratio of observed and expected abundances of species.
3.4.Sexual systems,fruit type and fruiting time
Of all species,74.1%(2347) produced dry fruits and 25.9%(819)produced fleshy fruits.We found that sexual system and fruit type were significantly correlated (Fig.9a).In the DHV region,plants that produce dry fruits were predominantly hermaphroditic or monoecious,whereas those that bear fleshy fruits tended to be dioecious.We also found that fruit type significantly correlates with growth form(Fig.9b):woody plants produced a high proportion of fleshy fruits,whereas herbaceous plants produced more dry fruits.However,independent of sexual systems,dry-fruited plants tended to flower during the rainy season and fruit during the dry season,whereas fleshy-fruited plants tended to flower during the dry season and fruit during the rainy season (Fig.10).
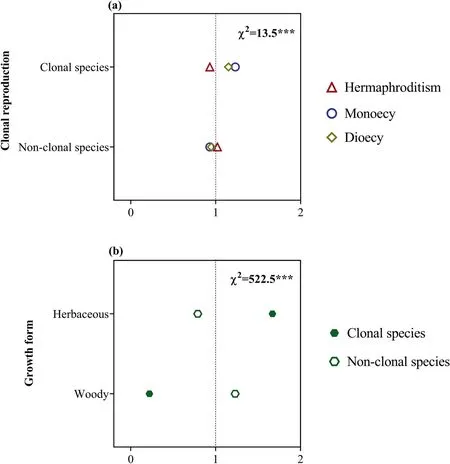
Fig.9.Ratio of observed and expected abundance of different sexual systems in different fruit types(a)and of different fruit types in different growth forms(b)in the dry-hot valley region of southwestern China.The value of the ratio >1 indicates that the specific sexual system or fruit type are overrepresented for the trait group under the assumption that sexual system and fruit type,and fruit type and growth form are statistically independent.χ2 indicates the reliability of the differences;***P<0.001;0-2 are the ratio of observed and expected abundances of species.
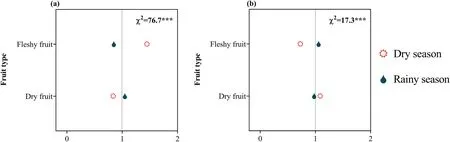
Fig.10.Ratio of observed and expected abundance of different flowering time (a) and fruiting time (b) in different fruit types in the dry-hot valley region of southwestern China.The value of the ratio>1 indicates that the specific flowering time or fruiting time is overrepresented for the trait group under the assumption that the flowering time and fruiting time and the fruit type are statistically independent.χ2 indicates the reliability of the differences;***P<0.001;0-2 are the ratio of observed and expected abundances of species.
4.Discussion
4.1.High incidence of diclinous plants in the DHV
Similar to the mountain top area (i.e.,the alpine subnival ecosystem),DHV regions have been shown to have low pollintor richness(Yi et al.,2021),which may limit the pollination success of plants in these areas (Blionis and Vokou,2001;Totland,2001;Liu et al.,2006;Wang et al.,2019).Previous studies have also shown that when pollinators are absent,hermaphroditic plants have an advantage over diclinous plants (e.g.,Xu et al.,2023).Accordingly,we expected that in the DHV region of southwestern China the frequency of diclinous species would be particularly low and the frequency of hermaphroditic species would be particularly high(Peng et al.,2012,2014).However,we found that the proportion of dioecious species was 5.6%higher in the DHV than in the subnival belt of the Hengduan Mountains.In fact,the proportion of diclinous(dioecy and monoecy) species was much higher in DHV region(25.5%)than in temperate regions(Table 2).One explanation for the high incidence of diclinous species within the flora of DHV region may be the high proportion of tropical elements.The DHV is dominated by subtropical and tropical monsoon climate and is located in a transitional region of north subtropical and plateau climate.With the rapid geological uplift of the eastern Qinghai-Tibet Plateau,the nearly north-south direction ridges and valleys may facilitate species migration between warm temperate regions in the north and subtropical and tropical regions in the south.Therefore,the flora in the DHV region of the southwestern China included many tropical and temperate elements at both generic and species level.At the species level,51.3%of all species of the flora were tropical,37.8% were temperate,and only 10.9% were cosmopolitan.Previous studies have shown that dioecy is more commonly represented in tropical than in temperate regions(Bawa,1980;Vamosi et al.,2003;Vary et al.,2011).In our study,analyses based on the whole flora of the dry-hot valleys and on the dry-hot valley of each river showed that dicliny was strongly associated with tropical elements,whereas hermaphroditism was strongly associated with temperate elements (Figs.3a and S1).Furthermore,the frequency of monoecious species was similar to that of dioecious species in tropical elements (Table 2).
An additional factor that may contribute to the high incidence of diclinous species in DHV region is plant growth form.Our analyses showed that dioecy was strongly associated with woody plants,whereas hermaphroditism was strongly associated with herbaceous plants (Fig.4).These findings are consistent with previous studies(Renner and Ricklefs,1995;Vamosi et al.,2003;Vamosi and Queenborough,2010;Peng et al.,2014;Ohya et al.,2017) and expand our understanding of the evolution of sexual systems to include a role for growth form.These findings also help explain the unexpectedly high frequency of diclinous plant species in the DHV region.Specifically,in this region woody plants composed a high proportion of the flora(46.2%).In contrast,the proportion of woody plants was much lower in Siberia (6.7%) and subnival belt of the Hengduan Mountains (8.4%) (Peng et al.,2014;Godin,2017).In addition,7.9%(249 species)of the herbaceous plant species in DHV region were grasses (e.g.,EragrostisandCymbopogon(Poaceae)),which are strongly associated with monoecy.
4.2.Flower morphology and its association with sexual systems
High temperature or low air humidity have been found to have the potential to accelerate the respiration and transpiration of flower organs(Primack,1985).In addition,large flowers,in general,need more carbon hydrates and water to meet respiration and transpiration rates (Teixido et al.,2016).Thus,smaller flowers are favorable in hot and dry environments(Galen,2005;Teixido et al.,2016;Song et al.,2022).However,in the flora of the DHV,the highest proportion of flowers were large (>1.0 cm wide).One possible reason for such a pattern is that most plants(74.7%)in the DHV flower in the rainy season.Plants that flower in the rainy season generally maximize the utilization rate of water to produce large flowers,as large floral displays can attract more pollinators(Arroyo et al.,2006).
Floral morphology is also known to evolve in response to selection by pollinators (reviewed by Van der Niet and Johnson,2012).Plants with generalized floral morphology usually have small,open or actinomorphic flowers,allowing them to be generally visited,and pollinated,by diverse groups of pollinators(Richards,1997).In contrast,flowers with specialized morphology(e.g.,zygomorphic flowers) are often visited by few pollinators.Thus,generalized pollination systems have been hypothesized to evolve in flora where pollinators are rare or unpredictable (Waser et al.,1996).In addition to low pollinator diversity and richness,generalist pollinators or inefficient pollinators are more common than specialists,especially in dry season(Liu et al.,2015).Thus,we found that in the DHV,generalized pollination systems were more common,especially in species that flower in the dry season(Figs.7 and 8).In addition,our results showed that plants with small and inconspicuous perianths(e.g.,yellow)generally flowered in the dry season(Fig.7).This finding is consistent with a previous study that showedTerminalia franchetii(Combretaceae) bloomed in dry season,and had generalized flowers pollinated by both wind and insects(Liu et al.,2015).We also found that plants with specialized floral morphology were more likely to flower in the rainy season(Fig.7),suggesting that specialized pollinators may be more frequent in this season.
We found that sexual systems were significantly correlated with floral morphological traits,suggesting that these associations are the result of strong selection for pollination(Peng et al.,2014;Wang et al.,2017).For example,in our study area,hermaphroditism was strongly associated with specialist pollination syndrome (Fig.6e).One possible explanation for this pattern is that the hermaphroditism can to some extent compensate for the pollinator deficiency that accompanies specialized pollination systems in unpredictable pollination environments,while specialized pollination systems carry a lower risk of geitonogamous pollination when pollinator availability is reliable (Johnson and Steiner,2000;Barrett,2002).In contrast,dioecious and monoecious species depend completely on vectors for pollination,so diclinous species are disadvantaged in harsh environments with fewer pollinators(Peng et al.,2012).However,diclinous species have evolved some reproductive strategies to succeed in the harsh pollination environments of the DHV.First,we found that monoecy was strongly associated with anemophily (Fig.6e);importantly,these species(e.g.,EragrostisandCymbopogon;Poaceae)do not need pollinating insects but instead rely on wind for pollination.Second,some monoecious plants can attract pollinators by special signal;for example,the floral scents ofAmorphophallus albus(Araceae)played an important role in attracting pollinators (rove beetles) (Tang et al.,2020).In addition,we found that diclinous species was strongly associated with clonal reproduction (Fig.5a).Clonal reproduction can partly compensate for failure of sexual reproduction by producing new individuals and keep them alive for a long time (Gao et al.,2020).
4.3.Fruit type and its association with sexual systems
Abundant precipitation is essential for the growth and development of fleshy fruits because water is the most abundant component of most fleshy fruits (Coombe 1976;Chen et al.,2017).Accordingly,fleshy fruits are more common in humid areas,especially in tropical rainforests (Chen et al.,2004,2017;Vary et al.,2011;Zhao et al.,2018).In our study area,savanna-like vegetation patches occur mainly at dry habitats of the deep valleys,and water availability is low due to the“foehn effect”(Yunnan Meteorological Bureau,1983).Therefore,it is not surprising that fleshy-fruited species accounted for a very low proportion (25.9%) of plants in the DHV region.In addition to reducing water consumption,dry fruits have a dispersal advantage over fleshy fruits.For fleshy fruits,seed dispersal usually depends on birds and mammals (Bawa,1980;Vamosi et al.,2003;Vary et al.,2011;Ohya et al.,2017).However,seed dispersers (e.g.,birds and mammals) are expected to be particularly scarce in the DHV as the presence of mammals and birds is positively correlated with precipitation (Grayson,1998;Liang et al.,2020).In contrast,seeds in dry fruits(e.g.,capsula and achene)can be easily dispersed by ballistics or by wind.
Our results showed that dry fruits were significantly associated with hermaphroditic and monoecious plants,while fleshy fruits were significantly associated with dioecious plants(Fig.9a),which supports earlier findings from floras with tropical and subtropical climates(Bawa,1980;Givnish,1980;Vamosi et al.,2003;Vary et al.,2011).Moreover,we found that fruit type was also related to plant growth form,with woody species being more likely to produce fleshy fruits and herbaceous species being more likely to produce dry fruits(Fig.9b).Therefore,in the DHV a high proportion of fleshy fruits was found in dioecious species with woody growth form.This finding can be explained in two ways.First,large and long-lived woody species can accumulate more nutrients and water used to produce high-cost fleshy fruits,and woody species producing fleshy fruits may better advertise to frugivores than herbaceous species (Wang et al.,2021).Second,dioecy might be more easily established in species producing fleshy fruits dispersed by birds or mammals because female fitness could be enhanced disproportionately by the production of attractive fruits.Fleshy fruits are generally related to long-distance dispersal and most fleshy-fruited species have the potential to migrate from the neighboring subtropical and temperate regions(Zhu et al.,2020),which can partly explain the high proportion of dioecious plants in the DHV.
5.Conclusion
Our study provides the first flora-level investigation of sexual system patterns in the DHV region of southwestern China.Despite the extremely harsh pollination environments in the DHV region,we found a high incidence of diclinous (dioecy and monoecy)species.We also found that sexual systems were strongly associated with geographic elements and various life-history traits.Our findings shed light on previous hypotheses about the associations between sexual system and various life-history traits,reflecting reproductive strategies used by the plants growing in the DHV region.Our results also suggest the necessity to consider the geographic origin of the flora when investigating the evolution and ecology of sexual systems.This study improves our understanding of plant reproductive adaptation in the DHV region.
Author contributions
B.S.,H.S.and D.L.P.conceived and designed the study.R.M.,Q.X.and Y.Q.G.collected the data.R.M.analyzed the data.M.R.,D.L.P.and B.S.wrote the manuscript.All authors read,commented and approved the final version of the manuscript.
Data availability
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgements
We would like to thank Prof.Da-Rong Yang for providing photos.This work was supported by the Key Projects of the Joint Fund of the National Natural Science Foundation of China(U23A20149),the Strategic Priority Research Program of the Chinese Academy of Sciences (XDA20050203),the Second Tibetan Plateau Scientific Expedition and Research program (2019QZKK0502),the National Natural Science Foundation of China (32371702,32071669 and 31770249),the West Light Foundation of the Chinese Academy of Sciences Interdisciplinary Innovation Team Program (xbzg-zdsys-202319),and the Ten-thousand Talents Program of Yunnan Province (YNWR-QNBJ-2018-208).
Appendix A.Supplementary data
Supplementary data to this article can be found online at https://doi.org/10.1016/j.pld.2023.07.010.

- 植物多样性的其它文章
- pyIFPNI: A package for querying and downloading plant fossil data from the IFPNI
- Intraspecific floral colour variation in three Pedicularis species
- Photosynthetic response dynamics in the invasive species Tithonia diversifolia and two co-occurring native shrub species under fluctuating light conditions
- Reproductive height determines the loss of clonal grasses with nitrogen enrichment in a temperate grassland
- Sex-specific facilitation and reproduction of the gynodioecious cushion plant Arenaria polytrichoides on the Himalaya-Hengduan mountains,SW China
- Different mechanisms underlie similar species-area relationships in two tropical archipelagoes