
产后女性脑镜像同伦功能连接改变的静息态fMRI研究
2024-04-12 06:37张凯华林嘉玉宿薇高樱莉李娜杜钰煊蔺星儒王晶赵诗佳杜小霞
磁共振成像 2024年3期
张凯华,林嘉玉,宿薇,高樱莉,李娜,杜钰煊,蔺星儒,王晶,赵诗佳,杜小霞
0 引言
女性生育后,会经历一系列母性行为、生理和心理上的变化[1]。影像学研究表明产后女性的大脑结构和功能也发生了改变。前人在研究产后女性应对婴儿刺激的脑反应中发现,产后女性的杏仁核、内外侧前额叶皮质、下丘脑和前扣带皮层、眼窝前额皮质[2]等脑区活动发生改变,即和情绪调节、奖励与动机、同理心和执行力等功能相关脑区发生了变化[3]。我们课题组前期对产后女性应对婴儿啼哭的脑激活中发现,与运动、言语、处理听觉刺激和照顾行为相关的脑区(如辅助运动区、额下区、颞上区、中脑和纹状体)的活动显著增强[4];静息态功能磁共振成像(resting-state functional magnetic resonance imaging,rs-fMRI)研究也揭示,产后女性随时间推移大脑自发神经活动发生动态变化,与未育女性间存在显著差异[5]。HOEKZEMA 等[6]和我们前期的研究[7]均提示,产后女性的脑结构表现出动态可塑性,包括灰质体积和皮质厚度的降低。这些研究结果一致表明,健康女性在产后期间为应对外在因素、身体(激素等)、或心理(母亲角色转换等)变化,其大脑发生了结构和功能的适应性变化,即产后女性的脑机制存在动态可塑性[8]。
双侧脑半球之间的同位连接性是最重要的功能基础结构特征之一,反映了双侧脑半球之间的信息交流与整合,对于协调两半球间的活动有重要意义。体素镜像同伦连接(voxel-mirrored homotopic connectivity, VMHC),是一种新的rs-fMRI分析指标,描述了大脑半球每个体素与其在另一侧半球镜像体素之间的静息态功能连接。VMHC 可用于评估双侧大脑半球之间的功能连接程度,反映大脑功能过程的协同性[9]。目前,VMHC 已广泛应用于阿尔茨海默病[10]、孤独症[11]、抑郁症[12]等神经精神系统疾病的研究,这些研究揭示VMHC 对于探测脑半球间功能协调的改变具有敏感性。
已有研究表明与产后健康女性相比,产后抑郁女性双侧背内侧前额叶皮层、背侧前扣带皮层和眼窝前额叶皮层的VMHC 值显著降低[13-14]。但目前并未有研究直接比较健康产后女性与未育女性之间的双侧大脑半球之间的功能连接程度,也尚未揭示健康产后女性脑半球间功能协调的改变。并且以往对产后女性的研究主要分为两个方向,一是探索产后女性大脑对婴儿刺激(声音、面孔等)的反应[2-4],二是探究产后女性大脑结构的变化,如灰质体积和皮层厚度等变化[6-7,15]。从功能连接角度探究产后女性神经机制的自发变化研究较少,仅有两篇采用静息态功能连接(resting-state functional connectivity, rs-FC)方法揭示产后女性边缘系统[16]和认知功能[17]相关脑区的连接改变,尚未有研究采用VMHC 方法评估产后女性大脑功能过程的协同性。因此,本研究拟采用VMHC方法分析产后女性双侧大脑半球的同伦连接功能改变,通过与未育女性对比,观察产后女性双侧大脑半球间同源神经元的同伦功能连接,为探讨产后女性神经机制的自发变化填补空白。
1 材料与方法
1.1 一般资料
本研究为横断面调查研究,在华东师范大学上海市磁共振重点实验室进行。本研究于2015 年7 月至2017 年7 月期间通过广告和网络在上海招募了47 名产后1年内的健康女性(婴儿年龄:8.16±3.82个月)和26名未育女性,剔除4名在扫描过程中头动过大的产妇和4名二胎产妇,最后有39名健康产后女性和26名未育女性的数据纳入分析。本研究经华东师范大学伦理委员会批准(伦理编号:201508001),所有受试者在参与研究前均签署了知情同意书。本试验中所有程序均遵循《赫尔辛基宣言》的指导方针和条例。
健康产后女性纳入标准:(1)健康产妇,年龄25~35 岁,右利手;(2)怀孕周期37~40 周;(3)无产后抑郁症,爱丁堡产后抑郁量表评分<3;(4)视力或矫正视力正常。未育女性纳入标准:(1)未生育过婴儿,年龄25~35岁,右利手;(2)视力或矫正视力正常。所有被试(包括健康产后女性和未育女性)排除标准:(1)严重躯体疾病、脑创伤、药物和酒精依赖史;(2)任何可能导致大脑功能紊乱的疾病,如失眠障碍、神经衰弱等;(3)精神病或神经系统疾病史;(4)磁共振扫描禁忌证。
采用由詹氏修订的中文版人际反应指针量表(Interpersonal Reactivity Index-C, IRI-C)[18]评估母亲的母性共情[19]。该量表测量多维共情项目,包括个人痛苦(personal distress, PD)、共情性关心(empthy concern, EC)、观点采择(perspective taking, PT)和想象力(fantasy, FS)。FS 和PT 分量表可以测量共情的认知要素,而PD 和EC 分量表能够衡量共情的情感内容[18]。量表共包含22个条目,采用Likert五点评分系统,受试者从0 分(不恰当)到4 分(非常恰当)中选择。由于修订删除了6 个条目,中文版IRI-C 量表共包含22 个条目,已被证明具有良好的效度和信度[20]。本研究中,产后女性组Cronbach'sα系数为0.779,未育女性组Cronbach'sα系数为0.755。
1.2 仪器与方法
本研究在华东师范大学上海市磁共振重点实验室扫描,采用Siemens Prisma 3.0 T Trio Tim MR 仪,配备12 通道头部线圈。扫描时,所有受试者都被要求仰卧、放松,闭上眼睛保持清醒,尽量不要进行任何思维活动,并使用定制的泡沫垫来减少头部运动。扫描序列包括两部分。(1)高分辨3D-T1 加权结构像,采用梯度回波脉冲序列收集。采集参数为:TR 2 530 ms,TE 2.34 ms,翻转角7°,矩阵256×256,视野256 mm×256 mm,切片数192,层厚1.0 mm,序列总时长363 s;(2)rs-fMRI 的图像采用基于梯度回波平面成像的全脑平面回波成像(echo planar imaging,EPI)序列收集,共210 个全脑体积。采集参数为:TR 2 000 ms,TE 30 ms,翻转角90°,切片数33,视场220 mm×220 mm,矩阵64×64,层厚3.5 mm,层间距25%,序列总时长426 s。
1.3 图像预处理
所有fMRI 原始数据均采用基于Matlab R2020.a平台的DPARSF 5.3(https://rfmri.org/DPARSF)进行预处理。预处理过程如下:将DICOM 数据转换为NIFTI 图像,去除前10 个受试者数据不稳定的时间点;之后进行时间校正和头动校正,排除头部平动方向超过2 mm 或者转动超过2°的数据;再将校正后的图像标准化到蒙特利尔神经研究所(Montreal Neurological Institute, MNI)的标准脑空间,将空间标准化后的图像重新采样到3 mm×3 mm×3 mm体素大小;之后选择6 mm 半高全宽高斯平滑核进行空间平滑;随后对数据进行去线性漂移,并进行带通滤波(0.001~0.010 Hz)去噪;最后对头动、全脑白质及脑脊液信号等协变量进行线性回归。
1.4 VMHC分析
使用DPARSF 5.3 工具包对预处理结果进行VMHC 分析。将所有受试者的T1 图像标准化至MNI 空间取平均值,得到T1 图像标准化平均值。然后,通过标准化T1 图像的平均值及其左右镜像的平均值生成特定群体的对称大脑模板。再将预处理后的静息态功能图像标准化到生成的特定群体对称脑模板,计算大脑一侧半球内每个体素和另一侧半球对应体素时间序列之间的Pearson 相关系数(VMHC值),随后对该相关系数进行FisherZ-转换以提高正态性,生成各受试者的VMHC图,把年龄作为协变量回归进行组间比较。
1.5 统计学分析
使用SPSS 26.0 里的独立样本t检验分析产后女性和未育女性在年龄、受教育年限和IRI-C 评分的差异,采用Benjamini-Hochberg 方法,以FDR 进行多次比较校正,P<0.05认为差异具有统计学意义。将年龄和受教育年限作为协变量,使用SPM12软件对产后女性和未育女性的VMHC值进行双样本t检验,结果报告以体素水平P<0.001 未校正且团块水平P<0.05 的FDR校正为标准。在纳入数据的受试者中,有一例产后女性的行为学评分缺失,因而被排除在行为学分析之外。使用xjview 将组间VMHC 值差异脑区定义为感兴趣区域(region of interest, ROI),提取每个受试者ROI 内的平均VMHC 值。利用SPSS 26.0,对VMHC值和IRI-C评分进行Pearson偏相关分析,其中年龄和受教育年限作为协变量纳入。考虑到探索性分析和相对较小的样本,未进行多次比较校正。
2 结果
2.1 社会人口学及心理测评结果分析
产后女性和未育女性与在年龄上差异有统计学意义(P<0.001),在受教育年限上差异无统计学意义(P=0.281)。产后女性的EC 评分高于未育女性(P=0.001),其余评分包括PT、FS、PD 和IRI-C 差异均无统计学意义(P均>0.05),见表1。

表1 产后女性和未育女性的社会人口学资料及心理测评量表比较Tab.1 Sociodemographic data and psychometric scales of postpartum womenand nulliparous women
2.2 产后女性和未育女性VMHC值比较
与未育女性相比,产后女性在双侧梭状回、海马旁回、额中回、额内侧回、中央前回、壳核和豆状核等脑区的VMHC 值显著降低(FDR 校正,体素水平P<0.001,团块水平P<0.05,双尾)(表2,图1)。将阈限放宽,发现产后女性在双侧楔前叶、顶上小叶和顶下小叶的VMHC 值显著增加(FDR 校正,体素水平P<0.01,团块水平P<0.05,双尾)。
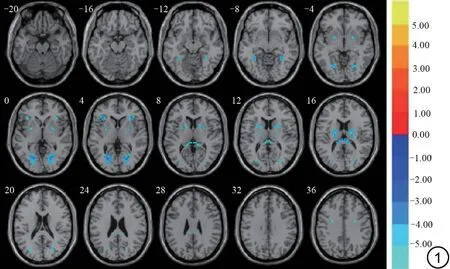
图1 VMHC 值在产后女性和未育女性之间的差异图。冷色调显示产后女性较未育女性VMHC值降低的脑区。VMHC:体素镜像同伦连接。Fig.1 The difference diagram of VMHC value between postpartum women and nulliparous women.Cold colors show the brain regions where the VMHC value of postpartum women is lower than that of nulliparous women.VMHC: voxel-mirrored homotopic connectivity.
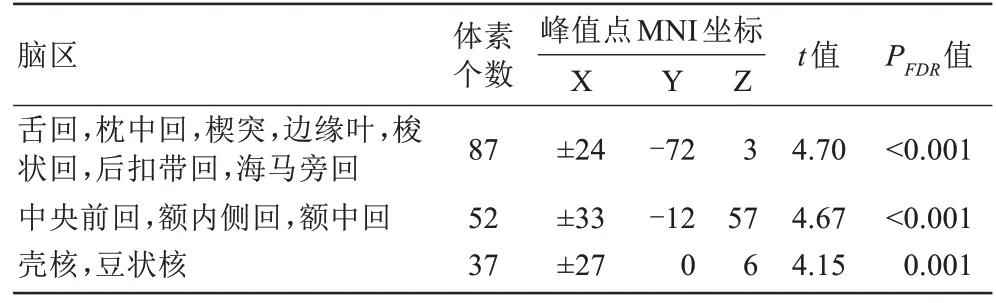
表2 产后女性较未育女性VMHC值显著降低的脑区Tab.2 The brain regions with decreased VMHC value in postpartum womencompared with nulliparous women
2.3 组间差异脑区VMHC值与IRI-C量表偏相关分析
以年龄和受教育年限为协变量,产后女性梭状回、后扣带回和海马旁回的VMHC 值与FS 评分(r=-0.345,P=0.039)和PD 评分(r=-0.351,P=0.036)呈负相关,见图2。

图2 以年龄和受教育年限为协变量的偏相关图。2A:产后女性FS 评分与梭状回、后扣带回和海马旁回VMHC 值的偏相关图;2B:产后女性PD 评分与梭状回、后扣带回和海马旁回VMHC 值的偏相关图。VMHC:体素镜像同伦连接;FS:想象力;PD:个人痛苦。Fig.2 Partial correlation diagram with age and education as covariates.2A: Partial correlation graph between FS score and VMHC value of postpartum women fusiform gyrus, posterior cingulate gyrus and parahippocampal gyrus; 2B: Partial correlation between PD score and VMHC value of postpartum women fusiform gyrus, posterior cingulate gyrus and parahippocampal gyrus.VMHC: voxel-mirrored homotopic connectivity; FS: fantasy; PD: personal distress.
3 讨论
本研究通过静息态VMHC 分析方法,发现产后女性大脑在与认知、情感、运动和言语、处理听觉刺激和护理等相关脑区,大脑半球间功能连接性发生了显著变化。揭示了产后女性抚养婴儿大脑发生适应性变化,为探讨产后女性神经机制提供了科学依据和客观参考。
3.1 产后女性情感-认知-奖赏网络VMHC变化
本研究发现,与未育女性相比,产后女性海马旁回、梭状回、额上回、额内侧回、中央前回、壳核和豆状核等脑区VMHC 值显著降低。研究表明,海马旁回[21]、梭状回[22]和额叶[23]均与情感认知功能相关。STEINBACH 等[24]发现,大脑两侧半球在某些功能上具有偏侧化,异侧脑区的功能活动存在差异性。右脑在对情绪信息的无意识加工中起到主导作用[25],并且右利手被试表现出明显的情绪偏侧化[26]。本研究发现产后女性在静息态下有关情感功能的部分脑区功能连接性降低,可能与产后女性发生大脑功能的偏侧化有关,产后女性大脑右半球对情感的自发反应比左半球更敏感,所以两侧脑区功能连接不同。同时产后女性这种自发性情感调节的变化,说明女性在生育完后为应对婴儿刺激,大脑发生了功能重组,脑半球间协同活动发生变化。这些改变有助于其更好地感知婴儿的情感,以便满足婴儿的成长需求。此外,梭状回也是面孔识别的核心脑区[27]。许多神经影像研究表明,识别单词和面孔涉及到梭状回中高度专业化的区域,分别为视觉词形区和梭状回面部区,二者具有侧向化的不对称性,视觉词形区通常偏左脑,而梭状回面部区偏右脑[28]。本研究发现产后女性在静息状态梭状回功能连接性降低,可能表明产后女性更容易进行自发地面孔识别,如更迅速、准确地识别婴儿的情绪面孔,以提高对婴儿的情绪的反应性[29]和敏感性[30-31]。壳核和豆状核作为基底神经节的重要结构,参与情感和奖赏等高级认知功能[32]。研究发现,婴儿图式会广泛激活与照顾、支持、情感和奖赏相关等脑区[2,4,33-34],尤其是独特的婴儿面孔会让奖励处理相关的神经区域激活增加[35]。MARIE 等[36]指出,不同奖励类型在半球优势和空间范围上有明显差异,食物奖励处理倾向于右半球,而金钱奖励倾向于双侧基底神经节包括其最前部伏隔核。因此,本研究发现产后女性在静息态下壳核和豆状核功能连接性降低,提示产后女性可能存在将婴儿视为有效奖励,并以支持和养育的方式对其作出反应的倾向[37]。
另外,我们放宽阈限后发现,与未育女性相比,产后女性楔前叶和顶上小叶VMHC 值显著增加。AGATHOS 等[38]发现,认知重构广泛诱发了核心默认网络脑区(default mode network, DMN)和额顶叶控制脑区的激活。楔前叶作为DMN的核心脑区[39],主要负责信息的收集与注意的分配[40],在自我和社会认知重组过程中起关键作用[38]。而一项脑卒中的研究[41]也发现,顶上小叶通过双侧额顶叶网络和半球间顶叶网络对认知进行了调节。因此,产后女性楔前叶和顶上小叶脑区功能连接增强,可能揭示其更愿意重新分配认知资源,将认知资源从内部集中注意力中分离出来,以便重新关注外部环境,更好地抚养婴儿。
3.2 产后女性和未育女性共情能力差异
行为学评分结果显示,产后女性比未育女性的EC 评分更高,表明产后女性的共情能力可能更高。前人研究发现婴儿情绪刺激会激活母亲大脑与共情加工[7]和心理理论[30]相关的脑区,这可能表明母亲更容易感知、协调和响应婴儿的情绪。较高的共情能力可以帮助母亲有效地感知和准确地理解婴儿的感受和愿望,有助于母亲对婴儿投入更多的情感和爱护,从而帮助母亲抚养婴儿[31]。
3.3 产后女性认知脑区VMHC变化与共情认知的关系
偏相关结果显示,产后女性梭状回、后扣带回和海马旁回的VMHC 值与FS 评分和PD 评分呈显著负相关,这表明产后女性梭状回、后扣带回和海马旁回的功能连接与想象力和痛苦感知有关。REX 等[42]发现,想象力与广泛的脑容量网络有关,包括双侧海马、舌回和额叶增加,以及伏隔核和默认模式网络(如楔前叶、后扣带回、横颞叶)的减少。而一项关于场景构建障碍的研究[43]也指出,后扣带皮层和海马旁回的损坏会损害想象虚拟场景的能力。本研究发现产后女性梭状回、后扣带回和海马旁回脑区的功能连接与想象力存在负相关,进一步证实了这些观点。此外,研究表明梭状回[22]和海马旁回[21]与情感认知功能有关。本研究发现产后女性梭状回和海马旁回与痛苦感知有关,可能因为痛苦是母亲感受婴儿的重要情感之一,痛苦感知能够及时提醒母亲关注婴儿的不安状况并做出调整,以便更好地养育婴儿。
3.4 局限性
本研究的不足与展望:首先,样本量相对较少,并且对照组年龄偏小,尽管我们在分析过程中已将年龄作为协变量加入统计分析,但是年龄这一因素可能仍对结果存在一定影响,未来我们将继续扩大样本量来平衡年龄。其次,本研究为横向对比,为了更好揭示产后女性神经机制的动态可塑性,我们后续进行了为期两年的纵向追踪,探讨产后女性神经机制随抚养婴儿行为的长期变化,该结果已发表[7]。
4 结论
综上所述,本研究使用VMHC 方法探讨初次生育完后的女性大脑静息态半球间功能连接。产后女性大脑在与认知、情感、奖赏、运动和言语、处理听觉刺激和护理等相关脑区,大脑半球间功能连接性发生了显著变化,可能是在产后期间为抚养婴儿发生的适应性变化。这一发现为产后女性脑区功能连接研究提供了新的思路,同时为产后女性认知、情感与奖赏相关脑区异常提供理论依据。
作者利益冲突声明:全体作者均声明无利益冲突。
作者贡献声明:杜小霞、张凯华构思和设计本研究的方案,实施研究并采集数据,对稿件的重要知识性内容进行了修改并参与了数据分析过程,其中张凯华获得了国家自然科学基金和山东省自然科学基金项目的资助;林嘉玉、宿薇起草了原始稿件,分析和解释本研究的数据;高璎莉、杜钰煊、李娜、蔺星儒、王晶、赵诗佳分析和解释本研究的数据,并对文中重要内容进行了修改。全体作者都参与了数据分析过程,并一致同意发表最后的修改稿,同意对本研究的所有方面负责,确保本研究的准确性和诚信。
猜你喜欢
临床精神医学杂志(2024年1期)2024-02-29
数学年刊A辑(中文版)(2023年4期)2024-01-04
分析仪器(2022年6期)2022-12-27
计算机集成制造系统(2022年11期)2022-12-05
家庭医学(2022年3期)2022-04-07
科学大观园(2022年4期)2022-02-22
计算机集成制造系统(2020年4期)2020-05-08
中国惯性技术学报(2019年1期)2019-05-21
中国化肥信息(2016年33期)2016-05-17
振动、测试与诊断(2014年6期)2014-03-01
