
幽门螺杆菌代谢物拮抗宿主先天免疫的机制研究
2024-04-09 08:00:34陈智林焕雄杨惠钿
中国免疫学杂志 2024年3期
陈智 林焕雄 杨惠钿
(1.潮州市中心医院消化内科,潮州 521021;2.潮州市湘桥区中医院检验科,潮州 521000)
幽门螺杆菌(Helicobacter pylori,H.pylori)感染了世界上一半以上的人口[1]。H.pylori感染的流行率和H.pylori毒力因子的主要基因型在不同地理区域差异很大[2]。H.pylori可以在恶劣的胃环境中持续数十年,其可破坏胃黏膜并改变胃激素释放模式,从而影响胃生理。通过利用各种毒力因子,H.pylori靶向不同的细胞蛋白调节宿主先天免疫反应(包括炎症反应),并启动对胃黏膜的多次打击,导致慢性胃炎和消化性溃疡[3]。H.pylori感染的长期后果之一是胃恶性肿瘤,特别是胃癌和胃黏膜相关淋巴组织(MALT)淋巴瘤[4]。因此,H.pylori已被国际癌症研究机构认定为Ⅰ类致癌物[5]。尽管H.pylori感染与感染部位被抑制的先天免疫之间存在密切的因果关系,但该过程所涉及的确切机制尚不明确。可以通过改变其表面分子逃避先天免疫受体的识别[6]。H.pylori还可通过抑制下游信号通路阻断其他先天识别受体。另一方面,H.pylori能够通过调节T淋巴细胞功能逃避宿主适应性免疫[7]。微生物代谢物与宿主先天免疫系统之间动态相互作用,并在维持微环境稳态和抑制炎症方面发挥关键作用[8]。然而,H.pylori的代谢物是否参与宿主的先天免疫仍未清楚。基于此,本研究旨在探讨H.pylori代谢物拮抗宿主先天免疫的潜在机制。
1 材料与方法
1.1 材料 人胃黏膜细胞GES-1购自北京欣盛百泰科技有限公司(货号:SC135);L-谷氨酰胺购自广州小凡科技有限公司(货号:76523-100MG);DMEM培养基购自广州硕谱生物科技有限公司(货号:D917531-500ml);脂多糖(LPS)购自上海源叶生物科技有限公司(货号:S11060-100mg);H.pyloriJP26菌株受赠于中国科学院微生物所;新生小牛血清购自广州市欣福联生物科技有限公司(货号:16010-159);TRIzol试剂购自杭州新景生物试剂开发有限公司(货号:5301100);NF-κB荧光素酶报告载体购自YEASEN公司(货号:11501ES03);RNeasy试剂盒购自广州誉维生物科技仪器有限公司(货号:74124);TNF-α ELISA试剂盒、IL-6 ELISA试剂盒购自广州俊吉生物科技有限公司(货号:HM10001、BO60009);IL-8 ELISA试剂盒购自北京冬歌博业生物科技有限公司(货号:DG10305H-48T);2-D-吡喃葡萄糖(2-D-Glucopyranose,2DG)购自北京奇松生物科技有限公司(货号:BQS145219-1g);本研究所用的TLR1、TLR2、TLR3、TLR4、TLR5、TLR6、TLR7、TLR8、TLR9、TLR10敲除GES-1细胞系由上海朝瑞生物科技有限公司进行敲除。TLR1、TLR2和GAPDH抗体购自Abcam公司(货号:ab68158、ab209216、ab8245)。
1.2 方法
1.2.1 细胞培养与处理 胃黏膜细胞GES-1培养于含有10%FBS、2 mmol/L L-谷氨酰胺、100 U/ml青霉素和100 μg/ml链霉素的DMEM培养基。使用终浓度为100 ng/ml的LPS处理GES-1细胞。
1.2.2H.pylori的培养 在37 ℃的微需氧条件下,H.pyloriJP26菌株在布鲁氏菌肉汤液体培养基或琼脂平板上生长,辅以10%新生小牛血清。
1.2.3 RNA测序 TRIzol试剂裂解仅LPS刺激的GES-1细胞、LPS与H.pylori培养上清共同处理的GES-1细胞及未处理的GES-1细胞。使用RNeasy试剂盒提取总RNA。RNA的质量用2100 Bioanalyzer检测,定量用NanoDrop 2000检测。选择RNA完整性数(RIN)≥8的RNA样品制备文库。RNA-Seq文库使用TruSeq®RNA Library Prep Kit v2进行。在HiSeq 2500(Illumina)中对文库进行双端测序(2×150 bp)。在数据分析之前,根据3个标准去除污染的原始reads:①去除带有adapter的reads:包含超过5个adapter污染碱基的reads被认为是adapter污染的reads,将被过滤掉;②排除频率>5%的未识别碱基的reads;③排除低质量的reads:低质量碱基数(Phred Quality 值<19)占总碱基数15%以上的reads被视为低质量reads。剩余的reads被认为是“干净的reads”用于生物信息学分析。使用Fisher精确检验进行通路富集分析,然后使用基于KEGG通路的Benjamini & Hochberg检验,使用每个肿瘤的差异表达基因。FDR<0.05的通路被认为是显著富集的通路。
1.2.4 NF-κB活性的检测 通过由NF-κB荧光素酶报告载体驱动的荧光素酶产生来测量H.pylori代谢物或16 h的H.pylori培养上清对NF-κB通路活性的影响。将15 μl GeneJuice在250 μl DMEM中在25 ℃下平衡 5 min,加入2 μg报告质粒并再孵育15 min,然后将混合物添加到约5×106个GES-1细胞中。立即将细胞接种于96孔板中,并使其黏附长达24 h。将H.pylori代谢物或不同浓度的2DG或LPS添加到新鲜培养基的细胞中。4~16 h后,根据制造商的说明使用商业试剂测量荧光素酶活性,并使用Wallac Victor2光度计从重复的孔中评估发光。
1.2.5 TNF-α、IL-6和IL-8分泌水平的检测 ELISA检测不同条件处理后GES-1细胞培养上清中TNF-α、IL-6和IL-8的分泌水平。
1.2.6 免疫共沉淀和免疫印迹 将2DG处理或不处理的GES-1细胞在Hanks缓冲液中充分洗涤,并用1 ml预冷的裂解缓冲液[20 mmol/L Tris( pH=8)、137 mmol/L NaCl、1%Triton X-100、2 mmol/L EDTA、10%甘油]裂解蛋白酶抑制剂PMSF(1 nmol/L)、亮肽素和抑肽酶(10 μg/ml)。裂解物以10 000 g离心5 min,收集上清液并与20 μl TLR1或TLR2抗体在4 ℃下孵育1 h。将沉淀在缺乏蛋白酶抑制剂的裂解缓冲液中彻底洗涤,并通过SDS-PAGE在10%聚丙烯酰胺凝胶和蛋白质印迹上进行分析。使用抗TLR1、TLR2或GAPDH抗体孵育过夜后,以HRP偶联的羊抗兔多克隆抗体孵育1 h,进行化学发光分析。
1.2.7 非靶向代谢质谱 收集培养4 h(对照组)和16 h(实验组)的H.pylori培养上清。将含有2 μg/ml L-2-氯苯丙氨酸作为内标的1 ml提取溶液(乙腈∶甲醇∶水=2∶2∶1)加入H.pylori上清中。涡旋振荡30 s后,将样品在40 Hz频率下均质化4 min,然后在冰水浴中超声处理10 min。均质化和超声处理循环重复3次。将样品于-40 ℃下孵育1 h,并在4 ℃下以10 000 r/min离心15 min。将总共800 μl上清液转移到新的试管中,并于37 ℃的真空浓缩器中干燥,然后冰上超声处理10 min,将干燥的样品在200 μl 50%乙腈中复溶,4 ℃下13 000 r/min离心15 min,并将75 μl上清液转移到新的玻璃小瓶中进行LC/MS分析。MS原始数据文件随后由ProteoWizard转换为mzXML格式,并由R包“xcms”处理。该过程包括峰解卷积、对齐和积分。Minfrac和截止分别设置为0.5和0.3。内部MS2数据库用于代谢物鉴定。缺失值被最小值的一半填充。将数据进行log2转换,然后进行总离子电流归一化。
1.2.8 分子对接 所有分子对接研究均使用AutoDock 4.2.6软件包进行。从RCSB蛋白质数据库(https://www.rcsb.org/)中检索到TLR2的晶体结构(ID:6NIG)。该结构是通过添加氢原子、去除水和添加Gasteiger电荷制备的。2DG的三维结构取自chemspider(https://www.chemspider.com/),ID为71358,通过添加氢原子和Gasteiger电荷制备。使用Propka 3.1在pH=7下测定受体蛋白和VP的质子化状态。然后将配体以默认参数对接到TLR2的结合位点,并导出20个对接结构以进行进一步的视觉分析。使用AutoDock 4.2计算每个结构的对接分数。结合自由能使用分子力学/泊松-玻尔兹曼表面积(MM-PBSA)方法计算。使用PyMOL 2.4.0(https://pymol.org/2/)构建2DG与TLR2对接结果的最佳结合结构。
1.3 统计学分析 所有统计分析均使用R软件包进行。两组间比较采用双尾t检验。多组间比较采用方差分析,差异有统计学意义后,其两组间比较采用双尾t检验,P值经Bonferroni法校正。P<0.05认为差异有统计学意义。
2 结果
2.1H.pylori代谢物抑制GES-1细胞的NF-κB通路 相较于仅LPS刺激的GES-1细胞,LPS与H.pylori培养上清共同处理后,多种基因的表达水平受到调控,并趋于未处理的GES-1细胞水平,见图1A。通路富集分析发现,上述基因主要存在于NF-κB通路,主要包括CCL2、CCL20、CCL4、CCL5、CCND1、CCNL1、CCRL2、CD44、CD69、CD80、CD83、CEBPD、CFLAR、CLCF1、CSF1、CSF2、CXCL1、CXCL10、CXCL11、CXCL2、CXCL3、CXCL6、CXCR7、CYR61、DDX58、DENND5A、FJX1、IFIH1、IFIT2、IFNGR2、IL8、IL15RA、IL18、IL1A、IL1B、IL23A、IL6、IRF1、IRS2、JAG1、JUN、KLF4、KLF6、KLF9、KYNU、LAMB3、MAP3K8、MARCKS、MCL1、NFAT5、OLR1、REL、RELA、RELB、RHOB、RIPK2、SMAD3、SNN、SOCS3、SOD2、TANK、TAP1、TNC、TNF、TNFAIP2、TNFAIP3、TNFAIP6、TNFAIP8、TNFRSF9、TNFSF9、TNIP1、TNIP2、TRAF1,存在于NF-κB通路的下游,见图1B。LPS与H.pylori培养上清共同处理后,NF-κB通路的活性受到抑制(P<0.05,图1C)。使用3KD超滤管过滤H.pylori培养上清,并采用过滤的滤液(代谢物部分)和截留的溶液(蛋白部分)处理LPS刺激的GES-1细胞,发现3KD超滤管过滤的滤液(代谢物部分)能够抑制NF-κB通路活性(P<0.05,图1D)。此外,H.pylori代谢物能够抑制NF-κB的磷酸化,见图1E。同时,H.pylori代谢物能够抑制NF-κB通路效应因子TNF-α、IL-6和IL-8的分泌(P<0.05,图1F~H)。
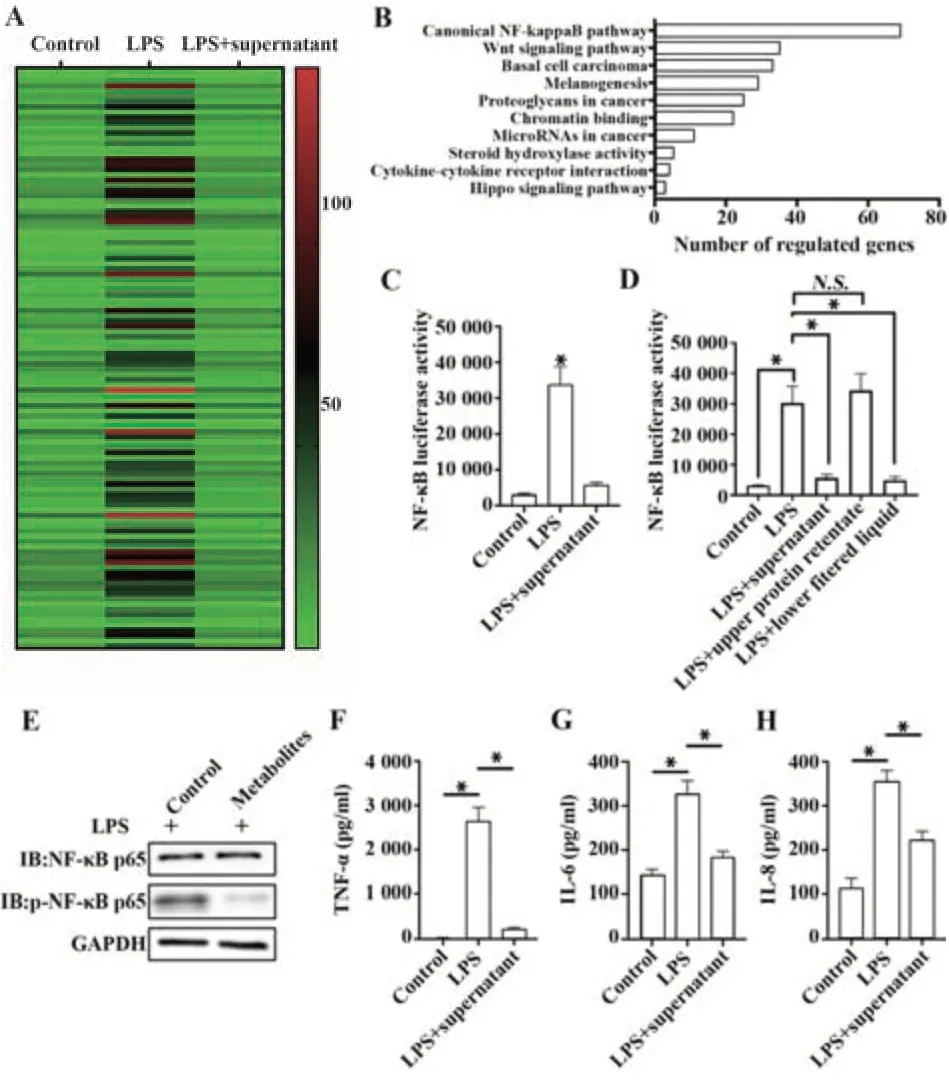
图1 幽门螺杆菌代谢物抑制GES-1细胞的NF-κB通路Fig.1 Helicobacter pylori metabolites inhibits NF-κB pathway of GES-1 cells
2.2 2 DG抑制GES-1细胞的NF-κB通路 将H.pylori代谢物进行非靶向代谢质谱,相比于对照组(培养4 h的H.pylori代谢物),培养16 h的H.pylori代谢物中有32种满足以下条件:①表达上调;②具有统计学意义,见图2A。在LPS处理的同时,将上述代谢物以5 μmol/L处理GES-1细胞,发现2DG处理后,GES-1细胞中NF-κB通路活性受到抑制(图2B)。在LPS处理的同时,使用1、5、25 μmol/L 2DG处理后,GES-1细胞中NF-κB通路活性均受到抑制(图2C)。在LPS处理的同时,1、5、25 μmol/L 2DG处理后,GES-1细胞中NF-κB通路效应因子TNF-α、IL-6和IL-8的分泌受到抑制(P<0.05,图2D~F)。在LPS处理的同时,1、5、25 μmol/L 2DG均可抑制NF-κB的磷酸化,见图2G。

图2 2DG抑制GES-1细胞的NF-κB通路Fig.2 2DG inhibits NF-κB pathway of GES-1 cells
2.3 2 DG拮抗宿主TLR2分子抑制NF-κB通路识别细菌刺激的先天免疫系统主要是TLR受体家族,且TLR受体家族是NF-κB通路的上游。因此,本研究构建了TLR1、TLR2、TLR3、TLR4、TLR5、TLR6、TLR7、TLR8、TLR9、TLR10敲除的胃黏膜细胞GES-1细胞系。使用LPS处理后,相对于对照组(Parent组),TLR1、TLR2、TLR4、TLR5、TLR6、TLR9、TLR10敲除的GES-1细胞中NF-κB活性显著降低(P<0.05);而TLR3、TLR7、TLR8敲除的GES-1细胞中NF-κB活性无显著变化。在LPS处理的同时使用2DG或H.pylori培养上清处理后,TLR3、TLR4、TLR5、TLR6、TLR7、TLR8、TLR9、TLR10敲除的GES-1细胞中NF-κB活性显著降低(P<0.05);而TLR1和TLR2敲除的GES-1细胞中NF-κB活性无显著变化(图3A)。使用2DG处理后,GES-1细胞中TLR1和TLR2的相互作用均减弱(图3B、C)。通过分子对接发现,2DG能够与TLR2氨基酸残疾R321、K347、F349结合,结合能为-12 kcal/mol(图3D)。构建TLR2野生型和突变型质粒(R321K、K347R、F349A),并分别转染TLR2敲除的GES-1细胞,在LPS处理的同时使用2DG或H.pylori培养上清处理后并不能减少转染TLR2突变型GES-1细胞的NF-κB活性(图3F)。
3 讨论
在本研究中,H.pylori代谢物2DG能够抑制NF-κB通路活性,抑制NF-κB磷酸化,抑制NF-κB通路效应因子TNF-α、IL-6和IL-8的分泌。有研究发现,H.pylori利用Ⅳ型分泌系统(T4SS)将CagA注入宿主胃上皮细胞,并通过激活NF-κB通路诱导炎症反应[9]。CagA是一种蛋白,应在3KD超滤管过滤的截留的溶液中(蛋白部分)。但本研究的蛋白部分不能影响NF-κB通路的活性,而代谢物2DG抑制了NF-κB通路。不同H.pylori菌株之间存在异质性。在许多非洲国家发现H.pylori感染率高,但H.pylori相关疾病发病率低[10]。因此,本研究所使用的H.pyloriJP26可能并不表达CagA蛋白。研究发现,在接种H.pylori后第1天,小鼠NF-κB活性和炎症反应显著上升[11]。然而,在接种H.pylori后第30天或第135天,胃中NF-κB活性水平下降。因此,H.pylori急性感染和长期感染时,NF-κB活性水平有所差异。本研究所用的是体外培养的H.pylori,其抑制NF-κB活性的代谢物2DG是否在H.pylori长期感染中抑制NF-κB活性值得进一步探讨。
在本研究中,使用LPS处理后,相对于对照组(Parent组),TLR1、TLR2、TLR4、TLR5、TLR6、TLR9、TLR10敲除的GES-1细胞中NF-κB活性显著降低;而TLR3、TLR7、TLR8敲除的胃黏膜细胞GES-1细胞中NF-κB活性无显著变化。Toll样受体(TLR)家族蛋白通过识别多种微生物分子(包括脂蛋白、脂肽、脂多糖、鞭毛蛋白和核酸)中的保守模式在先天免疫中发挥关键作用[12]。研究表明,TLR1、TLR2、TLR4、TLR5、TLR6、TLR9、TLR10能够识别细菌的模式识别受体,而TLR3、TLR7、TLR是识别病毒的基因组[13]。因此,TLR3、TLR7、TLR8敲除后,细菌独有物质LPS刺激下NF-κB的活性水平无显著差异。
TLR2与TLR1结合对于识别细菌脂蛋白和脂肽至关重要[14]。这些脂蛋白通过由脂质链修饰的保守N末端锚定在细胞膜上,并在巨噬细胞中诱导强烈的促炎信号[15]。TLR2缺陷小鼠对脂蛋白无反应,更容易患金黄色葡萄球菌引起的败血症、肺炎链球菌和单核细胞增生李斯特菌引起的脑膜炎,以及结核分枝杆菌感染[16]。因此,TLR2在细菌诱导的炎症反应中具有不可或缺的作用。本研究发现2DG能够与TLR2氨基酸残疾R321、K347、F349结合。研究发现,TLR1和TLR2胞外域的异二聚化是介导NF-κB通路活化所必需的[17]。TLR1和TLR2氨基酸形成疏水、氢键和离子相互作用,稳定蛋白质异二聚体。其中,TLR2的R321会与TLR1的E321和E366形成离子键,TLR2的K347会与TLR1的T361和T363形成氢键,TLR2的F349会与TLR1的Y320和V339形成疏水键。因此,2DG打断了这些键的形成,减少了TLR1和TLR2的相互作用,使TLR1-TLR2的异源二聚体不稳定,抑制了NF-κB通路活化。
综上所述,H.pylori代谢物2DG能够与TLR2相互作用,减少TLR2与TLR1异源二聚体的形成,抑制先天免疫NF-κB通路活性。
猜你喜欢
中国农业科技导报(2023年10期)2023-11-02 07:48:50
现代临床医学(2022年4期)2022-09-29 07:36:10
蜜蜂杂志(2022年5期)2022-07-20 09:54:16
海峡科技与产业(2021年1期)2021-05-22 01:26:20
服饰导报·鞋世界(2021年4期)2021-05-17 14:01:41
中国药理学通报(2019年5期)2019-01-11 18:03:39
知识就是力量(2016年8期)2016-11-02 01:00:11
分析测试学报(2015年7期)2016-01-13 06:19:16
质谱学报(2015年5期)2015-03-01 03:18:37
食品科学(2013年15期)2013-03-11 18:25:48
