
Semaphorin 7A impairs barrier function in cultured human corneal epithelial cells in a manner dependent on nuclear factor-kappa B
2024-03-20 06:33:32ChengChengYangXiuXiaYangXiaoJingZhaoHengWangZiHanGuoKaiJinYangLiuBinHuiLi
Cheng-Cheng Yang, Xiu-Xia Yang, Xiao-Jing Zhao, Heng Wang, Zi-Han Guo, Kai Jin,Yang Liu, Bin-Hui Li
1Department of Ophthalmology, the Fifth Affiliated Hospital,Sun Yat-sen University, Zhuhai 519000, Guangdong Province,China
2Department of Ophthalmology, Hangzhou TCM Hospital Affiliated to Zhejiang Chinese Medical University, Hangzhou 310006, Zhejiang Province, China
3Eye Institute of Xiamen University, Fujian Provincial Key Laboratory of Ophthalmology and Visual Science, School of Medicine, Xiamen University, Xiamen 361005, Fujian Province, China
Abstract
● KEYWORDS: human corneal epithelial; barrier function;transepithelial electrical resistance; zonula occludens-1;occludin; nuclear factor-kappa B
INTRODUCTION
The corneal epithelium is a protective barrier between the outside world and the eyes[1-2].The epithelial barrier and its integrity are essential for corneal homeostasis and resistant to environmental contaminants[3-4].Disruptions in the epithelial barrier function and increased permeability have been associated with various eye disorders[1,4], including diabetic keratopathy[5-6]and dry eye disease[7].Human corneal epithelial cells (HCEs) exert their protective effect on eyes by through their barrier function, which is largely regulated by tight junctions (TJs)[8].TJs, consisting of signaling molecules such as zonula occludens-1 (ZO-1), claudins, and occludin, are located on the surface of the corneal epithelium and contribute to enhancing the cornea’s barrier function[9-10].Dysregulated expression of these genes often serves as diagnostic markers for impaired epithelial barrier function[10].
Semaphorins represent a diverse family of membrane-anchored or released proteins, divided into eight subclasses based on structural characteristics and sequence similarity[11].Although our understanding of the role of semaphorins in epithelial barrier function, particularly their direct impact on TJs,remains limited[12], certain semaphorins, such as 3A, 3B, 3C,and 3F, have been found to enhance epithelial barrier function,while others, like 4D and 3E, promote barrier disruption,particularly in cancer progression[12].Among the semaphorins,semaphorin 7A (Sema7A) stands out as the sole member of class Ⅶ due to its unique glycosylphosphatidylinositol (GPI)anchoring on the cell membrane[13].Initially discovered as neuronal guidance proteins[14], Sema7A also exhibits various immunoregulatory and tumor-related actions, including influencing cell infiltration, interaction[15], and angiogenesis[16].Previous research has primarily focused on the relationship between Sema7A and junctions in tumors.Sema7A has been shown to be necessary for transforming growth factor(TGF)-β-mediated epithelial-to-mesenchymal transition by downregulating E-cadherin in mammary epithelial cells[17].
Sema7A is constitutively expressed in the cornea and is known to stimulate neuronal regeneration, inflammatory cell influx[18],and corneal neovascularization[19].However, its impact on the barrier function of HCEs has not yet been established.Thus,we explored the involvement of Sema7A in the dysfunction of the corneal epithelial barrier and elucidate its underlying mechanisms.These results may enhance our knowledge of the pathophysiology of Sema7A in corneal epithelial barrier dysfunction.
MATERIALS AND METHODS
Ethical ApprovalAll experiments were approved by the Ethics Committee of the Fifth Affiliated Hospital of Sun Yatsen University, Guangdong, China (Project identification code k27-1).
MaterialsDulbecco’s modified Eagle medium-nutrient mixture F12 (DMEM-F12), gentamicin, trypsin-EDTA,phosphate-buffered saline (PBS), and fetal bovine serum (FBS)were obtained from Invitrogen-Gibco (Rockville, MD, USA).4-kilodalton (kDa) Dextran-fluorescein isothiocyanate (FITC),bovine recombinant insulin, bovine serum albumin (BSA),human recombinant epidermal growth factor, and cholera toxin were provided by Sigma-Aldrich (St.Louis, MO, USA).Corning (Corning, NY, USA) supplied 24-well transwell plates as well as 6-, 24-, or 96-well culture plates.ZO-1 mouse monoclonal antibody was obtained from Invitrogen (Carlsbad,CA, USA), while occludin rabbit monoclonal antibody was obtained from Cell Signaling Technology (Beverly, MA, USA).Rabbit monoclonal antibodies to phosphorylated (p-) and unphosphorylated forms of inhibitor of kappa B alpha (IκBα)were obtained from Abcam (Beverly, MA, USA).Antibody to glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was obtained from Protein Tech Group (Pearl Street, Rosemont,USA).Mouse or rabbit IgG-specific horseradish peroxidase conjugated goat antibodies were supplied by the Beyotime Institute of Biotechnology.Recombinant human Sema7A was provided by R&D Systems (Minneapolis, MN, USA).IκB kinase 2 (IKK2) inhibitor Ⅳ was supplied by Calbiochem(Merck, Darmstadt, Germany).Enhanced chemiluminescence was obtained from Amersham Pharmacia Biotech (Uppsala,Sweden).Alexa Fluor 488-labeled goat antibodies to mouse or rabbit IgG were from Molecular Probes (Eugene, OR).The 4’,6-diamidino-2-phenylindole (DAPI) solution was obtained from Abcam (Beverly, MA, USA).The human interleukin(IL)-1β enzyme-linked immunosorbent assay (ELISA) kit was offered by RayBio Corporation (California, CA, USA), and the IL-1 receptor (IL-1R) antagonist (Anakinra) was purchased from MedChemExpress (Wisconsin, WI, USA).
Cell Culture and TreatmentSimian Virus 40 (SV40)-HCEs were immortalized by Araki-Sasakiet al[20]using an SV40-adenovirus recombinant vector lacking the origin of replication.They were provided by RIKEN Biosource Center (Tokyo,Japan).HCEs were grown in DMEM-F12 medium with 15% heat-inactivated FBS, 10 ng/mL recombinant human epidermal growth factor, 40 μg/mL gentamicin, 0.1 μg/mL cholera toxin, and 5 μg/mL bovine insulin, referred to as supplement hormonal epithelial medium (SHEM), at 37℃ and 5% CO2.Experiments were performed by plating HCEs at 1×105cells/well in 24-transwell plates or at 5×105cells/well, 5×104cells/well,or 3×103cells/well in 6-, 24-, or 96-well culture plates,respectively.HCEs were grown in SHEM for 2d and then in unsupplemented DMEM-F12 (DMEM-F12 with 40 μg/mL gentamicin) for 1d before being exposed to 0, 125, 250, or 500 ng/mL recombinant human Sema7A[21-22]or 0.1 μmol/L IKK2inhibitor Ⅳ[23]or 500 ng/mL IL-1R antagonist[24]for the indicated time in the latter medium.All experiments were repeated three times.
Cell Proliferation AssayFor the 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2Htetrazolium (MTS) assay, HCEs were plated on 96-well plates and treated with 0, 125, 250, or 500 ng/mL of Sema7A for 72h.Afterward, the cells were mixed with 20 μL of MTS assay solution and incubated for 4h.Cell proliferation was determined by measuring absorbance at 490 nm using a microplate reader.
Measurement of Transepithelial Electrical ResistanceTransepithelial electrical resistance (TEER) was measured using an EVOM voltameter (World Precision Instruments) to examine the integrity of the corneal epithelial barrier[4,25].After 48h of growth on a 0.4 μm filter in 24-transwell plates with SHEM as well as another 24h of growth in unsupplemented DMEM-F12, the HCEs were exposed to 0, 125, 250, or 500 ng/mL Sema7A for 72h.The TEER value (Ω●cm2) was obtained after multiplying the observed resistance by the surface area of the transwell filter.The TEER for each group was then adjusted by subtracting the background value for the blank transwell insert.The final TEER value was used for barrier function analysis.
Dextran-fluorescein Isothiocyanate Permeability AssayThe paracellular permeability of corneal epithelia was evaluated by the flow of 4 kDa Dextran-FITC, as previously reported[4,25-26].Briefly, after cells were exposed to 0, 125,250, or 500 ng/mL Sema7A for 72h, a 2-hour incubation with 200 μL Dextran-FITC (3 mg/mL) in the apical chamber at the temperature of 37℃ was performed.The supernatant was obtained from the bottom chamber.Next, the Envision microplate reader (PerkinElmer) with an excitation wavelength of 485 nm as well as an emission wavelength of 528 nm was used to determine the amount of Dextran-FITC.
Immunoblot AnalysisThe production of proteins, such as TJs and the nuclear factor-kappa B (NF-κB)-related components,in HCEs was evaluated utilizing immunoblot analysis.For the analysis of ZO-1 and occludin, HCEs were cultured in 6-well plates with SHEM for 2d, followed by an additional day in unsupplemented DMEM-F12.Subsequently, the cells were administered with 0, 125, 250, or 500 ng/mL Sema7A for 24,48, or 72h.For IκBα and p-IκBα analysis, HCEs were cultured in 6-well plates with SHEM containing only 0.5% FBS,followed by another day in unsupplemented DMEM-F12.Cells were then administered with 500 ng/mL Sema7A for 0, 5, 15, 30, 60, or 120min.To obtain cell extracts, cell lysis buffer radioimmunoprecipitation assay containing 1% protease inhibitors was used, following the provided instructions[27-28].Samples were subjected to centrifugation at 15 000 g for 10min at 4℃, and total protein concentrations were further measured and calculated.Approximately 20 μgof protein was separated from each sample using a 12.5% SDS-PAGEgel.The separated proteins were transferred onto polyvinylidene difluoride (PVDF) membranes (Merck Millipore).After blocking with 5% skim milk at room temperature (RT) for 1h, the membranes were incubated overnight at 4℃ with primary antibodies against ZO-1 (1:1000), occludin (1:1000),IκBα (1:1000), p-IκBα (1:1000), and GAPDH (1:4000).The membranes were then exposed to species-specific secondary antibodies (1:4000) for 1h.Protein bands were visualized using ECL reagents and recorded with the Tanon-5200 Multiimaging System (Tanon Science and Technology).The relativeexpression of the target proteins compared to GAPDH was analyzed using Image J.

Table 1 Sequences of primers
Quantitative Reverse Transcription-Polymerase Chain Reaction AnalysisTo examine Sema7A’s effects on the gene expression of TJs, HCEs were cultured in 6-well plates with SHEM for 2d, followed by an additional day in unsupplemented DMEM-F12.The cells were administered with 0, 125, 250, or 500 ng/mL Sema7A for 24, 48, or 72h.To assess Sema7A’s effects on the gene expression of IL-1β, HCEs were cultured in 6-well plates with SHEM for 2d, followed by another day in unsupplemented DMEM-F12.Cells were then treated with 500 ng/mL Sema7A for 24 or 48h.Total RNA was extracted from HCEs using the RNAprep pure Cell/Bacteria Kit (Tiangen).Subsequently, cDNA synthesis was conducted utilizing the Primescript RT Master Kit (Takara Bio Inc.) with 500 ng of RNA.Quantitative reverse transcription-polymerase chain reaction analysis (RT-PCR) detection was carried out utilizing the Bio-Rad (Hercules, CA) CFX96 optical reaction system.The relative fold changes in target gene expression were determined using the 2-ΔΔCT method.The primer sequences used in this study are presented in Table 1[1].
Immunofluorescent StainingFor ZO-1 and occludin staining,HCEs were cultured on coverslips in 24-well culture plates with SHEM for 2d, followed by an additional day in unsupplemented DMEM-F12.Subsequently, the cells were exposed to 500 ng/mL Sema7A for 72h with or without 0.1 μmol/L IKK2inhibitor Ⅳ.After fixing the HCEs with 100% methanol at RT for a duration of 20min, the cells were rinsed three times for 5min each with PBS with 0.1% Tween 20(PBST).Following that, the HCEs were blocked with 1% BSA in PBS (BSA-PBS) at RT for 1h, and then incubated overnight at 4℃ with antibodies against ZO-1 (1:100 in 1% BSA-PBS)and occludin (1:100 in 1% BSA-PBS).The next morning,the coverslips were washed three times with PBST, incubated at RT for 1h with Alexa Fluor 488-conjugated secondary antibodies (1:500 in 1% BSA-PBS), washed again three times with PBST, and stained with DAPI staining solution for 10min to visualize the nuclei.The results were observed and recorded using a fluorescent microscope (Oberkochen, Germany).
Measurement of IL-1β SecretionAfter exposure to Sema7A (500 ng/mL) for different durations, supernatants were harvested and subjected to centrifugation at 120×g for a duration of 5min, with the resulting supernatant stored at the temperature of -80℃[28].The released IL-1β in the supernatant was quantified using ELISA kits.
Statistical AnalysisData analysis was carried out using GraphPad Prism program version 9.0 (GraphPad Software,Inc.).Data were presented with mean±standard deviation(SD).Comparisons between variables were assessed utilizing Student’st-test.One-way analysis of variance (ANOVA) as well as Dunnett’s multiple comparisons were employed to determine intergroup variability.Pvalues of less than 0.05 indicated to be statistically significant.
RESULTS
Impairment of Barrier Function by Sema7A in Cultured Human Corneal Epithelial CellsTo evaluate the effect of Sema7A on the barrier function of HCEs, the TEER and flow of Dextran-FITC assay were carried out.The results revealed a statistically significant decrease in TEER among HCEs exposed to Sema7A (125-500 ng/mL) at 24, 48, and 72h compared to the control group at the corresponding time points in ANOVA analysis (P<0.05).Notably, the group treated with 500 ng/mL exhibited significantly lower TEER than the groups treated with 0, 125, and 250 ng/mL at 48h and 72h in ANOVA analysis (P<0.05).However, no significant difference was observed between the groups treated with 125 and 250 ng/mL at 24, 48, and 72h (P>0.05; Figure 1A).The impact of Sema7A was evident at 24h and showed the maximum decrease at 72h.A time-dependent trend in Sema7A’s inhibitory effect on TEER was observed, with a significantly stronger inhibition at 48h compared to 24h (P<0.05), while no significant change was observed between 48h and 72h (P>0.05).Additionally, the increased permeability of Dextran-FITC further confirmed the decreased barrier function induced by Sema7A (Figure 1B).Both experiments collectively demonstrate that Sema7A negatively affects the barrier function of HCEs.Moreover, the MTS assay indicated that Sema7A had no cytotoxic effect on HCEs.
Reduction of Protein Concentrations of Tight Junctions by Sema7A in Human Corneal Epithelial CellsTo elucidate the effect of Sema7A on barrier function in HCEs,we measured the protein levels of ZO-1 and occludin.We observed a dose-dependent reduction in the expression of ZO-1 and occludin proteins in HCEs (Figure 2A).The protein levels of ZO-1 showed a statistically significant decrease only at 500 ng/mL (Figure 2B) in ANOVA analysis (P<0.05),while occludin protein levels significantly decreased at both 250 ng/mL and 500 ng/mL (P<0.05; Figure 2C).Moreover,both ZO-1 and occludin protein levels decreased over time(Figure 2D) in ANOVA analysis (P<0.05), but the inhibitory effect of 500 ng/mL Sema7A on ZO-1 and occludin proteins was only statistically significant at 72h (P<0.05;Figure 2E, 2F).

Figure 1 Effects of Sema7A on the barrier function of HCEs A:Concentration- and time-dependent effects of Sema7A on the barrier function of HCEs.HCEs were cultured in 24-well transwell plates for 2d, followed by incubation in unsupplemented DMEM-F12 for an additional day.Then, HCEs were administrated with various concentrations (0, 125, 250, 500 ng/mL) of Sema7A for 24, 48,or 72h, and TEER was measured to assess the barrier function.aP<0.05 compared to the control at the same time point.The data is presented as mean±SD; n=3.B: To evaluate the paracellular permeability of HCEs, the fluorescent intensity of 4 kDa Dextran-FITC in the basal chambers was examined after 72-hour treatment with different concentrations (0, 125, 250, 500 ng/mL) of Sema7A.aP<0.05 compared to the control.The data is presented as mean±SD;n=3.Sema7A: Semaphorin 7A; HCEs: Human corneal epithelial cells;DMEM-F12: Dulbecco’s modified eagle medium/nutrient mixture F-12; TEER: Transepithelial electrical resistance; FITC: Fluorescein isothiocyanate; SD: Standard deviation.
Reduction of Tight Junctions mRNA Levels by Sema7A in Human Corneal Epithelial CellsUsing RT-PCR, we observed a dramatical decrease in the mRNA fold changes of ZO-1 and occludin among HCEs following Sema7A treatment in a time- and dose-dependent manner (Figure 3).Consistent with the protein level analysis, a statistically significant decrease in the mRNA levels of ZO-1 (Figure 3A) and occludin (Figure 3B) was only observed at 500 ng/mL (P<0.05), and these effects became apparent after 24h (P<0.05; Figure 3C, 3D) in ANOVA analysis.
Promotion of Phosphorylation of NF-κB Inhibitory Protein IκBα by Sema7A in Human Corneal Epithelial CellsTo further clarify the mechanism underlying the antibarrier function of Sema7A, HCEs were stimulated with 500 ng/mL Sema7A for various durations (ranging from 0 to 120min) to evaluate IκBα activity through phosphorylation.Results of Western blot analysis (Figure 4A) and quantitative analysis (Figure 4B) showed that the levels of p-IκBα/IκBα significantly increased upon exposure to Sema7A, reaching a peak at 5min (P<0.05).
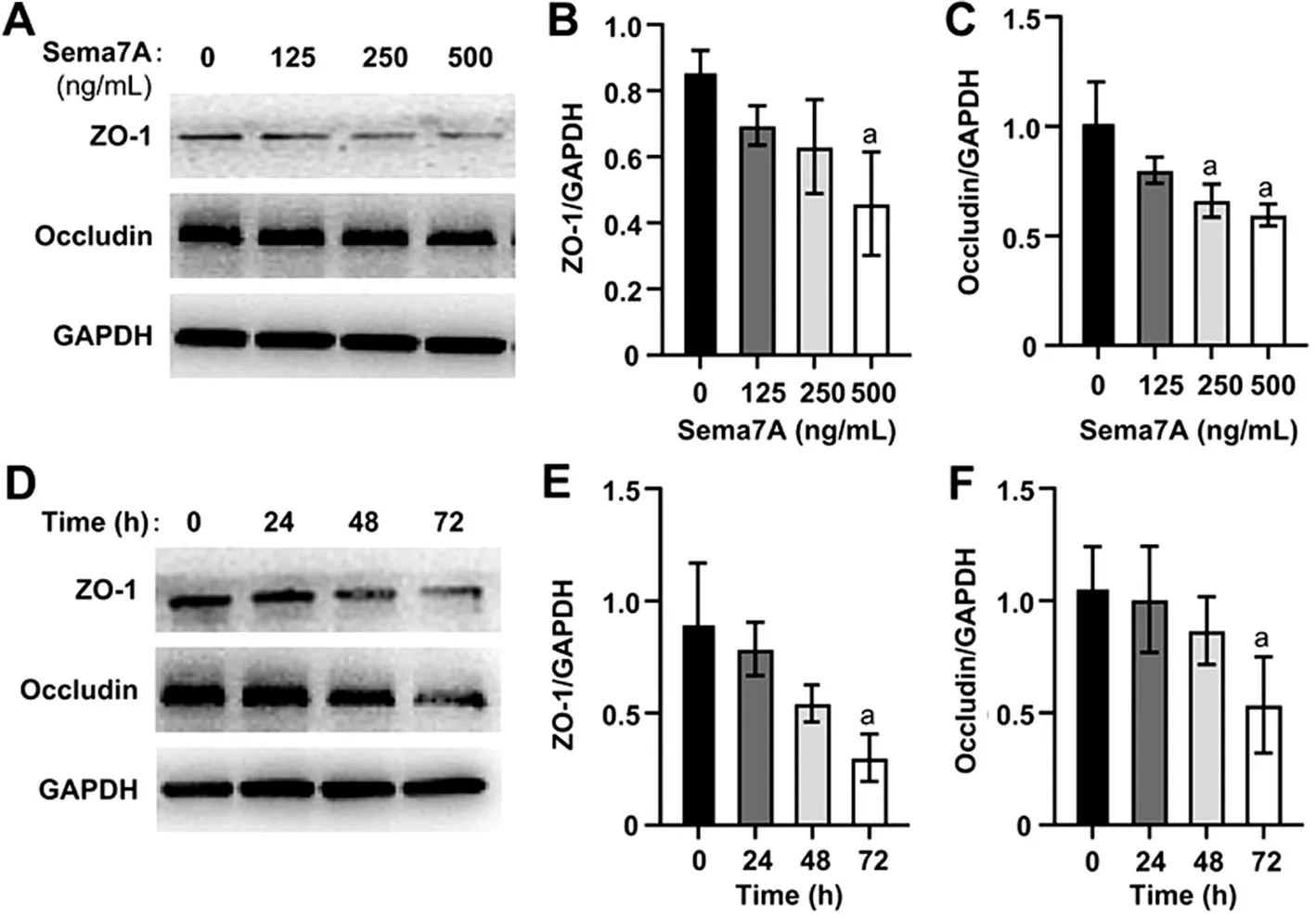
Figure 2 Concentration-dependent and time-dependent inhibition effects of Sema7A on the protein levels of the TJs in HCEs A-C: HCEs were cultured in SHEM for 2d, followed by a day in unsupplemented DMEM-F12.Then, HCEs were treated with different concentrations (0,125, 250, 500 ng/mL) of Sema7A for 72h.Representative immunoblots (A) and quantitative analysis (B, C) were used to quantify the protein levels of ZO-1 and occludin in HCEs.aP<0.05 compared to the control.The data is presented as mean±SD; n=3.D-F: HCEs were treated with 500 ng/mL Sema7A for 24, 48, or 72h after being cultured in SHEM for 2d and then unsupplemented DMEM-F12 for another day.Representative immunoblots (D) and quantitative analysis (E, F) were performed to assess the protein levels of ZO-1 and occludin in HCEs.aP<0.05 compared to the control.The data is presented as mean±SD; n=3.Sema7A: Semaphorin 7A; TJ: Tight junction; HCEs: Human corneal epithelial cells; SHEM: Supplement hormonal epithelial medium; DMEM-F12: Dulbecco’s modified eagle medium/nutrient mixture F-12; ZO-1:Zonula occludens-1; SD: Standard deviation.

Figure 3 Effects of Sema7A on the mRNA levels of the TJs in HCEs HCEs were cultured in SHEM for 2d, followed by a day in unsupplemented DMEM-F12.Subsequently, HCEs were treated with various concentrations (0, 125, 250, 500 ng/mL) of Sema7A for 72h (A, B) or with 500 ng/mL Sema7A for 24, 48, or 72h (C, D).RT-PCR was used to quantify the mRNA expression levels of ZO-1 and occludin.GAPDH was used as a reference gene.aP<0.05 compared to the control.The data is presented as mean±SD; n=3.Sema7A: Semaphorin 7A; TJ: Tight junction; HCEs: Human corneal epithelial cells; SHEM: Supplement hormonal epithelial medium; DMEM-F12: Dulbecco’s modified eagle medium/nutrient mixture F-12; RT-PCR: Reverse transcription quantitative polymerase chain reaction; ZO-1: Zonula occludens-1; GAPDH: Glyceraldehyde 3-phosphate dehydrogenase; SD: Standard deviation.
Inhibition of NF-κB Signaling Pathway Alleviated Sema7A-induced Negative Influences on Barrier FunctionSubsequently, pre-incubation with 0.1 μmol/L IKK2inhibitorⅣ for 2h successfully reversed the decreased TEER induced by 500 ng/mL Sema7A at 72h utilizing Student’st-test(P<0.05; Figure 5A).This finding was further supported by a decrease in Dextran-FITC permeability (P<0.05; Figure 5B).

Figure 4 Effects of Sema7A on the phosphorylation of IκBα stimulated by Sema7A in HCEs HCEs were cultured in SHEM containing 0.5% FBS for 1d, followed by an additional day in unsupplemented DMEM-F12.Then, HCEs were treated with 500 ng/mL Sema7A for different time intervals ranging from 0 to 120min.A:Immunoblots were performed to assess the protein levels of p-IκBα,IκBα, as well as GAPDH.Equal loading was confirmed by GAPDH.B: IκBα and p-IκBα proteins underwent densitometric analysis.Subsequently, the ratio of p-IκBα to IκBα in HCEs at various time points was calculated and compared using ANOVA analysis.aP<0.05 compared to the control; bP<0.05 compared to cells incubated with 500 ng/mL Sema7A for 5min.The data is presented as mean±SD;n=3.Sema7A: Semaphorin 7A; HCEs: Human corneal epithelial cells;FBS: Fetal bovine serum; SHEM: Supplement hormonal epithelial medium; DMEM-F12: Dulbecco’s modified eagle medium/nutrient mixture F-12; p-: Phosphorylated; IκBα: Inhibitor of kappa B alpha;GAPDH: Glyceraldehyde 3-phosphate dehydrogenase; SD: Standard deviation.
Inhibition of NF-κB Signaling Pathway Alleviated Sema7A-induced Negative Influences on the Degradation of Tight JunctionsIn addition, Western blot analysis (Figure 6A) and quantitative analysis (Figure 6B, 6C) revealed that pretreatment with IKK2inhibitor Ⅳ significantly restored the decreased protein levels of ZO-1 and occludin caused by Sema7A utilizing Student’st-test (P<0.05).Immunostaining assay (Figure 6D) revealed that in contrast with the control group, HCEs treated with 500 ng/mL Sema7A exhibited obviously less intense and discontinuous localization of ZO-1 and occludin.However, these effects were blocked by pretreatment with 0.1 μmol/L IKK2inhibitor Ⅳ.
Sema7A-induced IL-1β Secretion Promotion and Barrier Function Impairment ReversalviaIL-1R InhibitionFinally, RT-PCR (Figure 7A) and ELISA (Figure 7B) revealed that treatment with 500 ng/mL Sema7A significantly increased the mRNA and protein levels of IL-1β in HCEs at 48h, as indicated by ANOVA analysis (P<0.05).Pre-incubation of HCEs with 500 ng/mL IL-1R antagonist for 2h successfully reversed the decreased TEER induced by 500 ng/mL Sema7A at 48h (P<0.05; Figure 7C).This finding was further supported by a decrease in Dextran-FITC permeability by Student’st-test(P<0.05; Figure 7D).
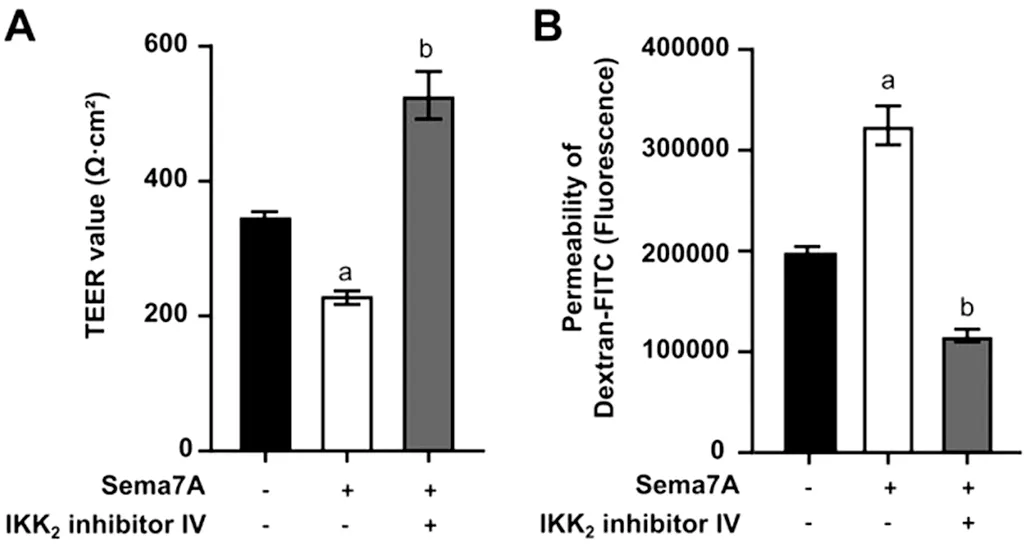
Figure 5 Effects of Sema7A on the tight junctions of HCEs after treatment with IKK2 inhibitor IV HCEs were pre-incubated with or without 0.1 μmol/L IKK2 inhibitor IV for 2h, and then incubated for 72h with/without 500 ng/mL Sema7A.TEER (A) and permeability of 4 kDa Dextran-FITC (B) were measured to evaluate the barrier function of HCEs.aP<0.05 compared to the control; bP<0.05 compared to cells incubated with 500 ng/mL Sema7A alone.The data is presented as mean±SD; n=3.Sema7A: Semaphorin 7A; HCEs: Human corneal epithelial cells; IKK2: IκB kinase 2 inhibitor; TEER: Transepithelial electrical resistance; FITC: Fluorescein isothiocyanate; SD: Standard deviation.
DISCUSSION
In this study, we have demonstrated that Sema7A impairs the barrier function of HCEsin vitro.This impairment was assessed by measuring TEER and Dextran-FITC permeability.We observed a time- and dose-dependent downregulation of ZO-1 and occluding at both the protein and mRNA levels,suggesting their involvement in the disruption of the barrier function.Our findings also indicated that the anti-barrier function of Sema7A might be mediated through the activation of the NF-κB signaling pathway.Additionally, we found that Sema7A promoted the expression of IL-1β, while inhibiting IL-1R partially reversed the detrimental effects of Sema7A on barrier function.

Figure 6 Effects of Sema7A on the expression of ZO-1 and occludin in HCEs HCEs were pre-incubated with or without 0.1 μmol/L IKK2 inhibitor IV for 2h and then incubated for an additional 72-hour with/without 500 ng/mL Sema7A.Representative immunoblots (A) and quantitative analysis (B, C) were performed to assess the protein levels of ZO-1 and occludin in HCEs after 72-hour different treatments.aP<0.05 compared to the control; bP<0.05 compared to cells incubated with 500 ng/mL Sema7A alone.The data is presented as mean±SD; n=3.Additionally, HCEs were fixed and immunofluorescence analyses (D) were performed using antibodies against ZO-1 (green)/occludin (green).DAPI staining was performed for nuclei (blue).Scale bar: 20 μm.Sema7A: Semaphorin 7A; ZO-1: Zonula occludens-1; HCEs: Human corneal epithelial cells; IKK2:IκB kinase 2 inhibitor; SD: Standard deviation; DAPI: 4’,6-Diamidino-2-phenylindole.
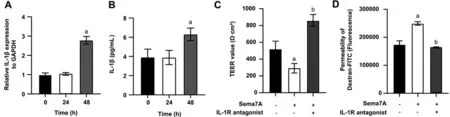
Figure 7 Sema7A-induced IL-1β secretion and barrier function impairment reversal via IL-1R inhibition in HCEs RT-PCR (A) and ELISA (B)demonstrated that 500 ng/mL of Sema7A significantly increased the mRNA and protein levels of IL-1β in HCEs at 48-hour.HCEs were preincubated with/without 500 ng/mL of an IL-1R antagonist for 2h, followed by incubation with or without 500 ng/mL of Sema7A.TEER (C)and permeability of the Dextran-FITC (D) were detected to evaluate the barrier function of HCEs after 48h of different treatments.aP<0.05 compared to the control; bP<0.05 compared to cells treated with 500 ng/mL of Sema7A alone.The data is presented as mean±SD; n=3.Sema7A:Semaphorin 7A; IL: Interleukin; IL-1R: Interleukin-1 receptor; HCEs: Human corneal epithelial cells; ELISA: Enzyme-linked immunosorbent assay; RT-PCR: Reverse transcription quantitative polymerase chain reaction; TEER: Transepithelial electrical resistance; FITC: Fluorescein isothiocyanate; SD: Standard deviation.
TJs, located beneath the apical surface, are responsible for maintaining the integrity of the paracellular route and preventing uncontrolled passage of macromolecules, cells,electrolytes and fluid[29].The TJ complex is composed of integral transmembrane proteins such as junctional adhesion molecules, occluding, and claudins.Furthermore, TJs includes membrane-associated proteins ZO-1, ZO-2, and ZO-3,which interact with cytoplasmic transmembrane proteins and bind them to the actin cytoskeleton[30].The proteins of claudin-1, occluding as well as ZO-1 are detectedin vivowithin the superficial/sub-superficial HCEs and related to the improvement of the barrier function[9,29].The cornea is densely innervated to maintain the ocular surface’s integrity.Nerve growth factors play a pivotal role in orchestrating the regeneration of corneal nerves and modulating inflammatory responses within the cornea.Recent studies have demonstrated the significant role of nerve growth factors in corneal epithelial wound healing progress, such as Netrin-1.Netrin-1 has been shown to reduce apoptosis,enhance proliferation and migration of HCEsin vitroandin vivo[31].Recombinant human nerve growth factor has also been found to promote corneal epithelial wound healing and nerve regeneration[32].Sema7A, as a neuronal guidance protein, is constitutively expressed in the cornea.In neurotrophic keratitis models of mice, Sema7A expression was increased in corneal epithelial cells by day 8.Furthermore, anti-Sema7A resulted in decreased corneal sensitivity and impaired reinnervation[33].These findings suggest that Sema7A may play a role in corneal disease.However, its exact impact on the corneal barrier function remains unclear.Our study discovered that Sema7A decreased the barrier function of HCEs, as evidenced by reduced TEER and increased Dextran-FITC permeability.Concurrently, we observed decreased transcription and translation of ZO-1 and occluding.This finding was consistent with previous studies on the barrier function of Sema7A in various diseases.For example, mice lacking Sema7A exhibited reduced blood-brain barrier permeability during West Nile virus infection[34].Additionally, Sema7A significantly promoted monolayer permeability during acute airway inflammation[35].These observations suggest that inflammation responses mediated by Sema7A contribute to the decline of barrier function in the brain and lung.Therefore, it is reasonable to propose that inflammation responses mediated by Sema7A may likewise lead to a decrease in barrier function in HCEs.
Previous studies have shown that Sema7A is activated during inflammation and has a potential in promoting inflammation,facilitating the dissemination of deleterious alterations essential for pulmonary inflammation in lung injury[36].The involvement of Sema7A in corneal inflammation has been investigated in previous research.Namavariet al[18]found that Sema7A significantly increased inflammatory cell influx into the cornea.Ghanemet al[19]observed an increase in corneal neovascularization and the presence of F4/80, a macrophage marker, in Sema7A-injected corneas, indicating that Sema7A might mediate an inflammatory response and recruit infiltrating cells such as macrophages.In our study, we observed that Sema7A increased IL-1β secretion in HCEs, which could potentially attract inflammatory cells by regulating the production of other inflammatory cytokines.Additionally,Sema7A has been shown to mediate inflammation in endothelial cells through the activation of integrin β1/FAK/MEK1/2/NF-κB signaling pathway[37-38].NF-κB activation is linked to the generation of various cytokines and growth factors and has been implicated in diminishing epithelial barrier function in other tissues[36].Based on these findings and previous research, we hypothesized that Sema7A may affect the barrier function by playing a role in NF-κB activation.
We also found that Sema7A induced the phosphorylation of the NF-κB-inhibitory protein IκBα.To further investigate the involvement of NF-κB signaling pathway within Sema7Amediated disruption of the barrier function in cultured HCEs,we inhibited this pathway using an IKK2inhibitor Ⅳ.IKK2inhibitor Ⅳ effectively suppressed NF-κB activation by degrading NF-κB inhibitor proteins in animal models and in culture conditions[39].We found that IKK2inhibitor Ⅳ blocked the negative effects of Sema7A on TEER and Dextran-FITC flux.Furthermore, the recovery of barrier function was linked to an elevation in the protein levels of ZO-1 and occluding.Immunostaining assay demonstrated that IKK2inhibitor Ⅳprevented the loss of the immunoreactivity of ZO-1/occluding detected at the cellular boundaries induced by Sema7A at 72h.These results provide evidence that disruption of corneal epithelial barrier function mediated by Sema7A in cultured HCEs is associated with the activation of the NF-κB signaling pathway.
As a major pro-inflammatory cytokine, IL-1β plays a crucial role in regulating inflammation and wound healing in the cornea[40].NF-κB positively regulates the expression of IL-1β[41],and IL-1β has been implicated in the disruption of ZO-1 and occludin at the junctions between adjacent HCEs, leading to impairment of corneal epithelial function[42].In our study,we observed that Sema7A promotes the secretion of IL-1β at both the protein and mRNA levels.This effect became more pronounced after 48h, although it was not significant at 24h.Importantly, the use of an IL-1R antagonist effectively suppressed the Sema7A-induced decrease in TEER as well as the increase in permeability.Based on these findings, we hypothesize that IL-1β likely plays a crucial role in the antibarrier function of Sema7A in HCEs.However, since the inhibitory effect of Sema7A on barrier function occurs at 24h,it is reasonable to assume that factors other than IL-1β may also be related to the early regulation of HCEs barrier function by Sema7A.
In conclusion, our study demonstrated that Sema7A might affect corneal barrier function by decreasing the expressions of ZO-1 as well as occludin at the translational/transcriptional levels, activating the NF-κB inhibitory protein IκBα, and enhancing the expression of IL-1βin vitro.The disruption of barrier function caused by Sema7A could be reversed by the IKK2inhibitor Ⅳ and IL-1R antagonists.These findings suggest Sema7A to be a potential therapeutic target for regulating corneal epithelial barrier function.
ACKNOWLEDGEMENTS
Foundations:Supported by the National Natural Science Foundation of China (No.81770889); Zhuhai Science and Technology Program (No.ZH22036201210134PWC).
Conflicts of Interest: Yang CC,None;Yang XX,None;Zhao XJ,None;Wang H,None;Guo ZH,None;Jin K,None;Liu Y,None;Li BH,None.
 International Journal of Ophthalmology2024年3期
International Journal of Ophthalmology2024年3期
- International Journal of Ophthalmology的其它文章
- Late infection after peri-orbital autologous micro-fat graft:a case presentation and literature review
- Stromal lenticule addition keratoplasty with corneal crosslinking for corneal ectasia secondary to FS-LASlK:a case series
- Clinical features and possible pathogenesis of multiple evanescent white dot syndrome with different retinal diseases and events: a narrative review
- Utility of real-time 3D visualization system in the early stage of phacoemulsification training
- Efficacy of scleral buckling for the treatment of rhegmatogenous retinal detachment using a novel foldable capsular buckle
- Effect of navigation endoscopy combined with threedimensional printing technology in the treatment of orbital blowout fractures