
基于全细胞生物传感器的污染物生物有效性评估研究进展
2024-03-19 04:35谷彩文张小凯王震宇
福建农林大学学报(自然科学版) 2024年1期
谷彩文, 朱 一, 林 瀚, 张小凯, 王震宇
(江南大学环境与生态学院,江苏 无锡 214122)
近几十年来,自然资源的过度开发、生活和生产污染物的不合理处置导致各类环境污染问题频发。研究表明,全球已有超过14%的淡水径流被污染,污水总量超5.5亿t[1-2]。同时,大量的工业、农业、生活污水直接排入土壤,破坏土壤结构,影响微生物种群分布;作物积累的重金属等有害物质通过生物放大效应危害人体健康[3-4]。此外,调查显示,人口聚集地的大气污染水平远高于世界卫生组织所制订的空气质量标准[5]。因此,开展环境中污染物的检测和风险评估工作对人类健康发展和生态保护至关重要。
污染物浓度是研究污染物对生态系统和人体健康影响的重要指标,常用的检测方法包括电化学法、色谱法、原子光谱法、荧光法和比色法等[6-7]。这些化学方法适用于不同类型的污染物,可以有效地分析样品中污染物的总浓度。然而,污染物的总浓度并不能完全代表其对环境的毒害作用,无法有效反映污染物的生物毒性[8-10]。研究表明,生物有效性是判断污染物毒害大小的先决条件[11]。Wells et al[12]认为“没有生物有效性即没有生物毒性”。因此,在进行环境风险评估时,污染物的生物有效性是很重要的评估指标之一。
近年来,全细胞生物传感器在污染物生物有效性评估方面表现出了卓越的性能,在环境风险评估方面具有非凡的应用潜力[13]。全细胞生物传感器是基于基因工程的产物,能对环境中的目标化学物质做出感应,并产生可被检测的电化学或光学信号[14],通过信号的强弱判断环境中污染物的生物有效性。本文简要介绍全细胞生物传感器的分类、工作原理及其构建因素,重点阐述其在环境领域的应用,为推动该技术的商业化应用提供理论依据。
1 全细胞生物传感器的分类及工作原理
全细胞生物传感器通常以细菌为宿主菌株,利用一种或多种调控基因和报告基因产生一种可识别量化的信号[15]。图1为全细胞生物传感器检测环境中污染物生物有效性的工作机理。根据指示物被识别方式、信号转化及处理机制,全细胞生物传感器可以分为3大类:Ⅰ类、Ⅱ类和Ⅲ类[16]。Ⅰ类全细胞生物传感器为目标特异性全细胞生物传感器,参与特定遗传调控机制的启动子识别污染物后,通过报告基因的表达输出信号,从而检测特定污染物的生物有效性[17]。Ⅱ类全细胞生物传感器是应激特异性全细胞生物传感器,当感知到压力胁迫(如DNA损伤、蛋白质损伤、热休克)时,表现为信号输出[17]。该类全细胞生物传感器并不能区分是何种污染物造成的环境压力,因此其可被用于检测多重污染产生的环境风险。Ⅲ类全细胞生物传感器为非特异性全细胞生物传感器,通常由无毒环境中信号表达水平较高的细胞构建,当其暴露于存在有毒化学物质的环境中时,报告基因的表达被抑制,信号输出减弱[18-19]。这类全细胞生物传感器主要用于检测污染物的毒性。相比之下,Ⅰ类全细胞生物传感器的应用是最广泛的[20-23],下文将对其进行阐述。
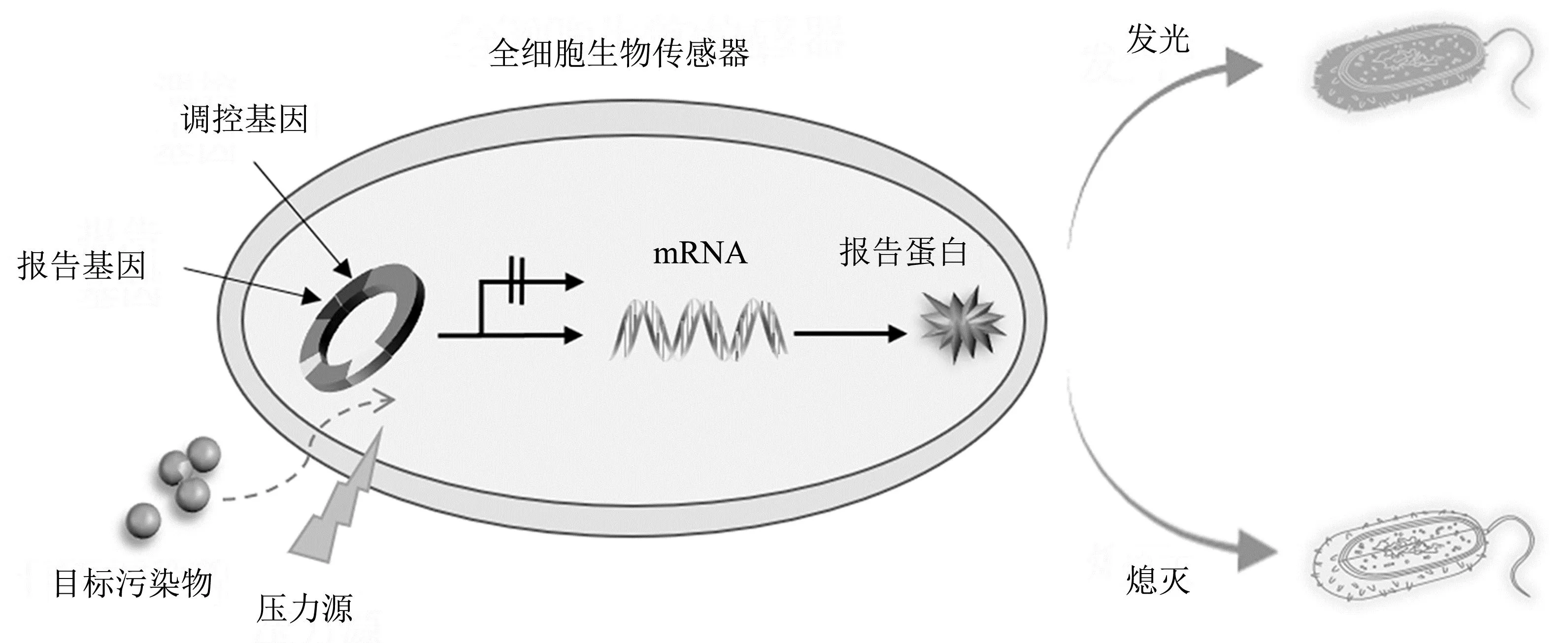
图1 全细胞生物传感器工作原理示意图Fig.1 Diagram of sensing mechanism of whole-cell bioreporters
2 全细胞生物传感器的构建
2.1 调控基因
调控基因和报告基因是全细胞生物传感器遗传回路编码的重要组成部分,这两类基因在质粒上的融合是全细胞生物传感器构建的基础。全细胞生物传感器的性能主要取决于调控基因对待测物的响应效果。当感知到目标化合物或应激因子时,全细胞生物传感器中的转录调控因子会刺激调控基因做出响应,诱导报告基因的转录并改变输出信号[24]。
调控蛋白的主要工作机制涉及能够识别同族化学物质的效应物的结合反应[25]。调控蛋白与操纵子特异性结合阻止或启动调控基因的转录,当感知到环境中的特定污染物时,效应物作用于调控蛋白,变构的调控蛋白与操纵子的结合状态发生变化,从而调控报告基因的表达[26]。例如:砷酸盐全细胞生物传感器中的调控蛋白ArsR,在没有砷酸盐存在的情况下可与ars操纵子相结合,并阻止其自身和下游ars基因的表达;而当环境中存在砷酸盐时,它会进入细胞内与ArsR蛋白相互作用,导致ArsR蛋白与操纵子相分离,使得ars基因能够表达并调控报告基因使其产生荧光信号[27]。
在某些情况下,具有相似结构的效应物能引起同一种调控蛋白的响应,如对苯产生响应的XylR蛋白也能与甲苯、二甲苯发生反应[28],对萘产生响应的phnR蛋白也能与菲发生反应[29]。
学者们通过大量的研究不断探索全细胞生物传感器构建的分子机制,并将其与合成生物学相结合[30],极大地促进了该技术的发展。例如,在细胞内插入融合好的基因已被证明可以有效地提高生物传感器的特异性,这些基因不仅能够整合多个信号的输入,也能合理地调整细胞内受体蛋白的水平,进而大大改善了全细胞生物传感器的检测极限[31]。Turner et al[32]将融合好的hbpCp-luxAB基因插入大肠杆菌(Escherichiacoli)中所构成的全细胞生物传感器,能够对羟基化联苯保持较高的特异性检测水平,而cadCp-lucFF基因能够操控全细胞生物传感器,同时可对锌、铅、镉等多种重金属产生响应[33]。
2.2 报告基因
当全细胞生物传感器暴露于目标物质中时,报告基因在调控基因的诱导下表达并产生可测量的信号。报告基因在信号传导过程中起着重要作用,其中应用最广泛的有β-半乳糖苷酶(lacZ)、荧光素酶(lux)和绿色荧光蛋白(gfp)3种基因[34]。
研究发现,lacZ基因相对比较稳定,当X-gal(lacZ的显色底物)作为水解底物时,lacZ能催化生成便于光学检测的蓝色产物[35-36]。当以对氨基苯基-β-D-半乳糖苷(4-aminophenyl-beta-D-galactopyranoside)为底物时,通过电化学方法可以检测酶解产生的对氨基酚(4-aminophenol)的量,从而确定lacZ的表达情况[37]。研究人员通过将lacZ基因与DNA损伤应答基因(recA)启动子结合,开发出了一种可有效检测环境中农药(如毒死蜱、丙苯氧磷、氯氰菊酯)遗传毒性的全细胞生物传感器,其可用于环境中基因毒性物质的风险评估[38]。
具有特定发光效应的lux是另一种比较常见的用于构建全细胞生物传感器报告系统的基因。Engebrecht et al[39]从弧菌(Vibriofurnissii)中分离出完整的lux(luxCDABE)基因,在此基础上构建的重组大肠杆菌是在不添加醛的情况下可产生生物发光信号的全细胞生物传感器。lux基因可在多种生物体中存在,如萤火虫荧光素酶(firefly luciferase)基因(由luc编码)和海肾荧光素酶(sea pansy luciferase)基因(由ruc编码)[40]。基于lux基因构建的全细胞生物传感器可以用于检测多种污染物的生物有效性。例如:基于luxAB基因构建的全细胞生物传感器可以有效检测苯酚的生物有效性[41];基于luxCDABE基因构建的全细胞生物传感器可以检测水体中铅的生物有效性,其检测限低至1.2 μg·L-1[13]。
gfp基因最早是从水母(Aequoreavictoria)中分离得到的,具有体积小(238个氨基酸)、能够在多种生物体内成功表达、无需任何外源底物或腺嘌呤核苷三磷酸(adenosine triphosphate)等特点,是目前最常用的报告基因[42]。然而,基于gfp的全细胞生物传感器面临着检测背景信号强、对污染物的响应较慢[24]等问题。为了解决这些问题,采用具有优越稳定性且能够发出更强荧光信号的增强型绿色荧光蛋白基因(egfp)是一个很好的选择。Kang et al[43]将zntAp与egfp融合,在此基础上构建的全细胞生物传感器可以对镉、汞、锌做出响应。Kim et al[44]基于egfp构建的全细胞生物传感器通过改变启动子区域的氨基酸序列,改变对重金属离子的选择性,从而能够检测环境中存在的多种金属离子。研究表明,以egfp作为报告基因的全细胞生物传感器已逐渐成为检测污染物生物有效性的重要工具[45-46]。
除了上述常用的报告基因外,还有一些基因也可被用作构建全细胞生物传感器。如:Stenzler et al[47]基于黄素荧光基因(PpFbFP)构建的全细胞生物传感器可在厌氧条件下检测环境中汞的生物有效性;以参与类胡萝卜素合成的crtA基因作为报告基因的全细胞生物传感器可以对环境中较低浓度的亚砷酸盐进行检测[48]。
2.3 宿主菌株
在构建全细胞生物传感器时,除了确定调控基因和报告基因外,宿主菌株的选择也尤为重要[11]。全细胞生物传感器通常以细菌作为宿主菌株,在不同污染物的刺激下,每个菌株的敏感性和响应度不同。因此,在实际检测时,针对不同的目标污染物,需要选择对其有特定抗性机制的生物作为宿主菌株[49]。
大肠杆菌已被广泛用作全细胞生物传感器的宿主菌株,用于检测典型环境污染物(如重金属和有机化合物)的遗传毒性[50]。除大肠杆菌外,其他革兰氏阴性细菌,如真氧产碱杆菌(Alcaligeneseutrophus)[51]和耐金属贪铜菌(Cupriavidusmetallidurans)[52],以及革兰氏阳性细菌,如枯草芽孢杆菌(Bacillussubtilis)和金黄色葡萄球菌(Staphylococcusaureus)[53-54]也被广泛用于全细胞生物传感器的构建。Magrisso et al[52]以耐金属贪铜菌为宿主菌株构建的全细胞生物传感器可用于检测土壤中多种重金属的生物有效性;Kim et al[33]利用枯草芽孢杆菌构建的全细胞生物传感器可对水环境中的铅、锌和锡等重金属做出响应。此外,真菌、酵母菌和藻类[55-56]也可作为全细胞生物传感器的宿主菌株。
3 全细胞生物传感器的应用
3.1 重金属检测
重金属由于具有较高的毒性、不可被降解和环境持久性等特点,对生态系统造成了巨大的威胁[57]。近年来,使用全细胞生物传感器评估重金属的生物有效性已经成为研究热点[58-59]。表1总结了全细胞生物传感器在重金属生物有效性评估方面的应用。研究表明,基于不同宿主菌株构建的全细胞生物传感器的灵敏度不同。例如:基于蓝细菌(Cyanobacteria)构建的全细胞生物传感器对砷的检测限为11.24 mg·L-1[60];而基于大肠杆菌DH5α构建的全细胞生物传感器对砷的检测限低至微克每升的级别[61]。因此,可以根据环境中重金属污染水平的差异选择不同的全细胞生物传感器进行生物有效性检测。
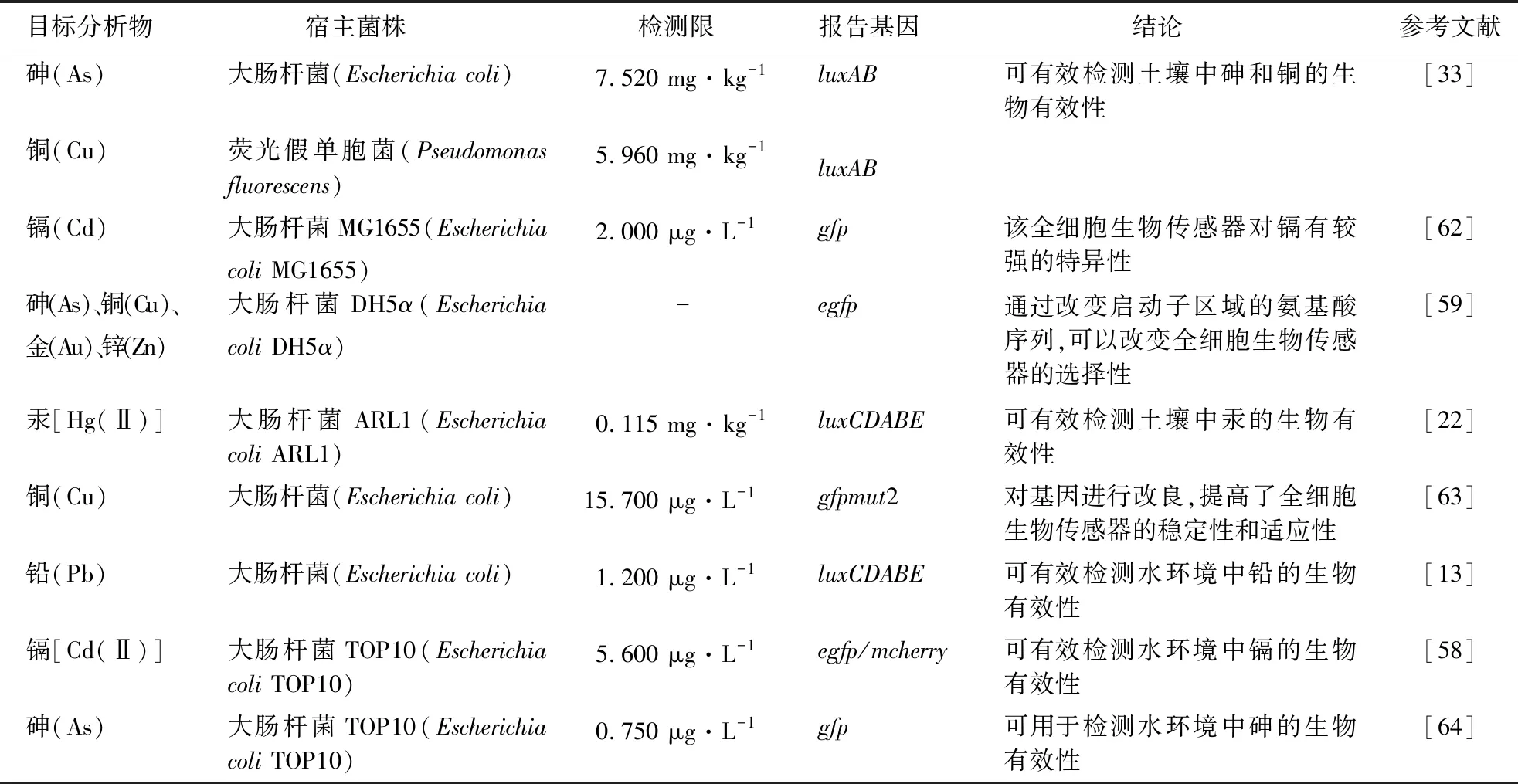
表1 全细胞生物传感器检测重金属生物有效性的相关研究1)Table 1 Recent studies on detection of heavy metal bioavailability by whole-cell biosensor
研究表明,重金属(尤其是生物非必需金属)在极低浓度下对大多数细菌都具有毒害作用[65]。因此,需要对全细胞生物传感器进行优化以提高其在较高浓度重金属环境下的检测性能。研究人员发现,在细胞内加入解毒基因能够增强其对重金属的耐受性[21]。例如,zntA和cadA是两个典型的金属转运蛋白编码基因,可用于构建能够在极端环境中工作的全细胞生物传感器[66]。Zhang et al[67]研究表明,基于zntA构建的全细胞生物传感器可以在1.2 μg·L-1~12.5 mg·L-1的质量浓度范围内检测铅的生物有效性。
Li et al[59]研究发现,全细胞生物传感器可以同时通过两个模式检测铅的生物有效性和毒性,而其毒性的检测是基于常规风险评估方法的生物配体模型。试验条件(如pH、细胞密度和培养基成分)的细微差异可能会对全细胞生物传感器的检测限造成影响[13]。Delatour et al[68]发现,通过改变细胞培养液中的营养物质如葡萄糖和木糖中氨基酸的浓度,可以对全细胞生物传感器的响应时间及生物发光信号的形态进行精细调整。特定的金属结合蛋白也可显著提高全细胞生物传感器的选择性和灵敏度。如全细胞生物传感器可通过在大肠杆菌中融合可编码调控蛋白MerR的汞抗性基因和启动子mer提高对汞的选择性[22]。此外,将全细胞生物传感器与基因信号放大器耦联也是一种比较常见的优化方式,在细胞内添加放大器有利于荧光响应信号的输出,进而提高全细胞生物传感器的灵敏度和特异性。研究表明,添加了“T7-RNA 聚合酶”放大器的全细胞生物传感器对镉离子响应的速度及特异性明显提高[69]。
3.2 有机污染物检测
有机污染物可以通过多种方式进入环境中,如未经处理的工业或生活污水的直接排放、船舶或海上勘探造成的石油泄漏,以及杀虫剂等污染物渗入土壤等[70]。研究人员通过构建不同的全细胞生物传感器,分别对苯酚、阿莫西林、氯仿等多种有机污染物的生物有效性进行了评估[71-73]。表2总结了应用全细胞生物传感器检测有机污染物生物有效性的相关研究。
Patel et al[81]发现,全细胞生物传感器对工业废水样品中的多种芳香烃化合物具有高灵敏性,检测限在0.1~1.0 μmol·L-1。为了维持全细胞生物传感器在复杂环境下检测的稳定性,研究人员用聚多巴胺包裹细胞,使得检测结果更加准确、有效,该传感器可用于检测土壤中的甲苯等有机物[82]。抗生素是一种新型有机污染物,其大量使用对环境造成了潜在的威胁。已有研究表明,以抗生素抗性基因启动子与报告基因融合构建的全细胞生物传感器可以用于评估抗生素在环境中的毒害作用[73]。
全细胞生物传感器对不同的有机污染物具有不同的敏感性,这一特性使其可被用于单种或混合有机污染物遗传毒性的高通量筛选[71]。自然环境条件会影响菌株的活性,进而影响全细胞生物传感器的检测结果。Plotnikova et al[82]对比了不同环境条件下全细胞生物传感器对萘生物有效性的最低检测限,结果表明,在气相中最低检测限为50 nmol·L-1,在水相中为0.5 μmol·L-1。萘的高挥发性是导致差异的主要原因。Zhang et al[83]研究了pH值对全细胞生物传感器检测性能的影响,发现pH值较高时宿主细胞可在铅浓度较低的情况下表现出毒性反应。这是因为pH值会影响宿主细胞的活性,进而影响全细胞生物传感器的检测结果。因此,如何克服环境因素的影响是发展该技术的关键。Hurtado-Gallego et al[84]研究表明,以蓝细菌作为宿主细胞的全细胞生物传感器可以有效检测污染源附近被污染的自然水体中污染物的生物有效性。蓝藻细胞易于培养,且具有较强的环境适应性,使其能够在更恶劣的环境条件下保持自身的活性。这说明宿主细胞对全细胞生物传感器的重要性。此外,全细胞生物传感器对实验室中标准检测物质的检测结果并不能完全代表该物质在自然环境下的生物有效性。例如,使用全细胞生物传感器对H2O2生物有效性进行检测时,自然环境中产生的甲基紫精的O2-会干扰检测的准确性,无法保证其特异性[85]。因此,如何保证全细胞生物传感器在自然环境下对污染物的检测性能值得重视。目前,研究人员已提出了解决方案,包括冷冻干燥、真空干燥、连续培养和固定在有机或无机源的生物相容性聚合物中[86-87]。将全细胞生物传感器封装在天然或合成聚合物中已被证明可用于检测环境污染物,聚合物基质提供了一个水合环境,其中包含细胞所需的营养物质及细胞活性和生长的辅助因子,被包裹的宿主细胞在环境中能免受有毒物质的侵害,并保持质粒的稳定性[88]。
4 总结
与传统方法相比,全细胞生物传感器具有快速、经济、操作简便等优点,是检测环境中污染物生物有效性的理想工具。然而,对于该技术的应用仍有几个问题需要解决,包括进一步缩短响应时间、提高细胞灵敏度和选择性,以及进一步推进其标准化应用等。未来应通过建立并完善全细胞生物传感器的标准检测方法,促进其商业化应用。
猜你喜欢
今日农业(2021年11期)2021-11-27
环境科学研究(2021年6期)2021-06-23
环境科学研究(2021年4期)2021-04-25
少儿科学周刊·儿童版(2021年23期)2021-03-24
矿产综合利用(2020年1期)2020-07-24
中成药(2018年8期)2018-08-29
当代化工研究(2016年6期)2016-03-20
中国资源综合利用(2016年3期)2016-01-22
中国药理学通报(2015年2期)2016-01-12
热带农业科学(2015年9期)2015-10-14
