
2020—2022年闽粤赣地区PRRSVORF5基因的分子流行病学调查
2024-03-18 13:45吕紫欣张鑫杰薛少华金艳冬陈林熙黄喜荣罗国清戴爱玲
福建农林大学学报(自然科学版) 2024年2期
吕紫欣, 张鑫杰, 薛少华, 金艳冬, 陈林熙, 黄喜荣, 罗国清, 蒙 宁, 陈 瑶, 戴爱玲,4
(1.福建农林大学动物科学学院,福建 福州 350002;2.龙岩学院生命科学学院,福建 龙岩 364012; 3.三明市动物疫病预防控制中心,福建 三明365001;4.福建省生猪疫病防控工程技术研究 中心/福建省家畜传染病防治与生物技术重点实验室,福建 龙岩 364012)
猪繁殖与呼吸综合征是由猪繁殖与呼吸综合征病毒(porcine reproductive and respiratory syndrome virus, PRRSV)引起的重要病毒性疫病之一,临床上主要导致妊娠母猪繁殖障碍以及其他阶段的猪群,尤其是仔猪出现呼吸障碍等症状[1]。PRRSV属于套式病毒目,是一种极易发生变异和重组的有囊膜的单股正链RNA病毒,包括至少10个开放阅读框(open reading frame, ORF)。其中,ORF5基因编码的GP5蛋白是PRRSV中最易变异的结构蛋白之一,通常作为PRRSV变异的主要依据,大量研究表明ORF5基因也可作为PRRSV基因组分型的重要依据[2-4]。
分子流行病学研究显示,我国流行的PRRSV可分为4个谱系:谱系8(JXA1-like株、CH-1a-like株)、谱系5(VR2332-like株、BJ-4-like株)、谱系3(GM2-like株、QYYZ-like株)、谱系1(NADC30-like株、NADC34-like株)[5]。近年来,PRRSV在我国的流行变异情况愈加复杂[6],2006年我国暴发由HP-PRRSV株引起的高致病性猪繁殖与呼吸综合征,2012年起我国陆续出现与NADC30-like株同源性较高的PRRSV,如NADC31-like株、QYYZ-like株、NADC34-like株[7]。NADC30-like株自2016年已超过HP-PRRSV株成为我国当前主要的流行毒株[8],给我国猪繁殖与呼吸综合征的防控带来了巨大的挑战。
为调查闽粤赣地区PRRSV的分子流行动态,本研究收集了2020—2022年闽粤赣地区1 475份发病猪组织和血清样品,采用实时荧光定量PCR(quantitative real-time PCR, qRT-PCR)对样品中的PRRSV进行了病原检测,对阳性样品ORF5基因进行了扩增、测序、序列比对及遗传进化分析。本研究旨在了解闽粤赣地区PRRSV的流行趋势和变异情况,对测序得到的62株PRRSVORF5基因开展遗传进化及分子流行病学分析,为闽粤赣地区猪繁殖与呼吸综合征的科学防控提供参考。
1 材料与方法
1.1 材料
1.1.1 病料样品来源 1 475份发病猪的组织样品(扁桃体、淋巴结、肺脏、脾脏、肾脏,n=397份)和血清样品(n=1 078份),分别采自福建龙岩、泉州、漳州,广东汕头、梅州,江西赣州。样品信息如表1所示。所有组织和血清样品均由龙岩学院动物医学研究所保存于-80 ℃。
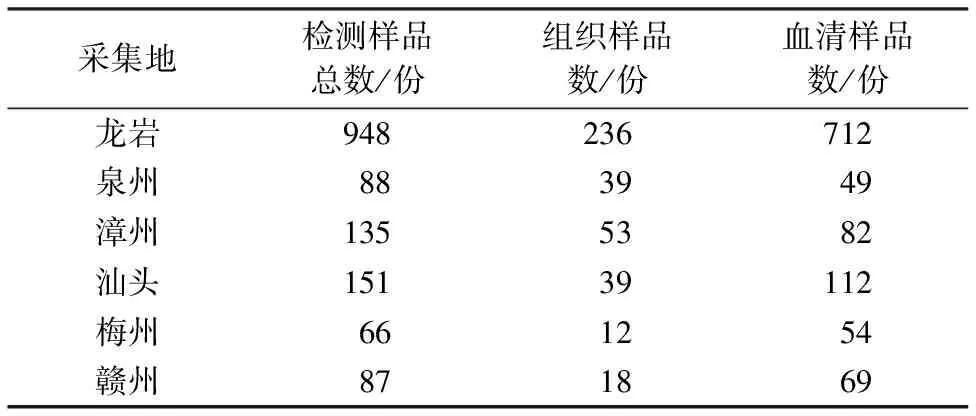
表1 样品来源信息Table 1 Information of sample source
1.1.2 试剂 反转录试剂、荧光定量PCR试剂和2×Phanta Flash Master Mix均购自南京诺唯赞生物科技有限公司。
1.2 引物的设计与合成
根据文献[9]的方法合成PRRSV qRT-PCR检测引物和探针。根据NCBI上的不同基因型PRRSV全基因组序列利用Oligo7软件设计扩增ORF5基因的特异性引物(上游引物序列:5′-GCRACGTTTTAGCCTGTCTT-3′;下游引物序列:3′-TGGCGTRTAKGTRATRGARAA-5′),预期引物扩增片段长度为750 bp。引物由生工生物工程(上海)股份有限公司合成。
1.3 组织和血清样品中PRRSV的qRT-PCR检测
从发病猪的扁桃体、淋巴结、肺脏、脾脏、肾脏等组织和血清样品中分别提取病毒总RNA,采用诺唯赞反转录试剂盒进行反转录以获得cDNA,获得的cDNA置于-80 ℃下保存,采用qRT-PCR检测cDNA。反应程序:55 ℃逆转录5 min;95 ℃预变性30 s;循环反应包括95 ℃ 10 s、60 ℃ 30 s,共设置45个循环。
1.4 PRRSV ORF5基因的扩增、克隆、测序
以“1.3”中获得的循环域值(Ct)较低的PRRSV cDNA为模板,使用ORF5上下游引物对PRRSV阳性病料进行单项PCR扩增。反应体系(25 μL):12.5 μL 2×TaqMaster Mix、2 μL引物对、2 μL样品cDNA、8.5 μL ddH2O。反应程序:95 ℃ 5 min;循环反应包括95 ℃ 30 s、55 ℃ 30 s、72 ℃ 30 s,共35个循环;72 ℃ 10 min。反应结束后取6.0 μL PCR扩增产物进行2%琼脂糖凝胶电泳检测,采用凝胶成像仪观察结果。对产物进行胶回收纯化,克隆于pMD19-T载体中,转化至大肠杆菌DH5α感受态细胞上,培养16~24 h后,挑取单克隆菌落经鉴定为阳性后送至广州睿博兴科生物技术有限公司进行测序。
1.5 PRRSV ORF5的遗传进化分析
应用DNAStar软件包中的MegAlign软件对获得的62条PRRSV与GenBank上已发表的2020—2022年闽粤赣地区18条PRRSV和127条PRRSV国内外参考株的ORF5基因序列(参考毒株信息如表2所示)进行核苷酸和氨基酸序列比对分析,并应用Mega 11.0软件中的最大似然法构建PRRSVORF5基因遗传进化树。
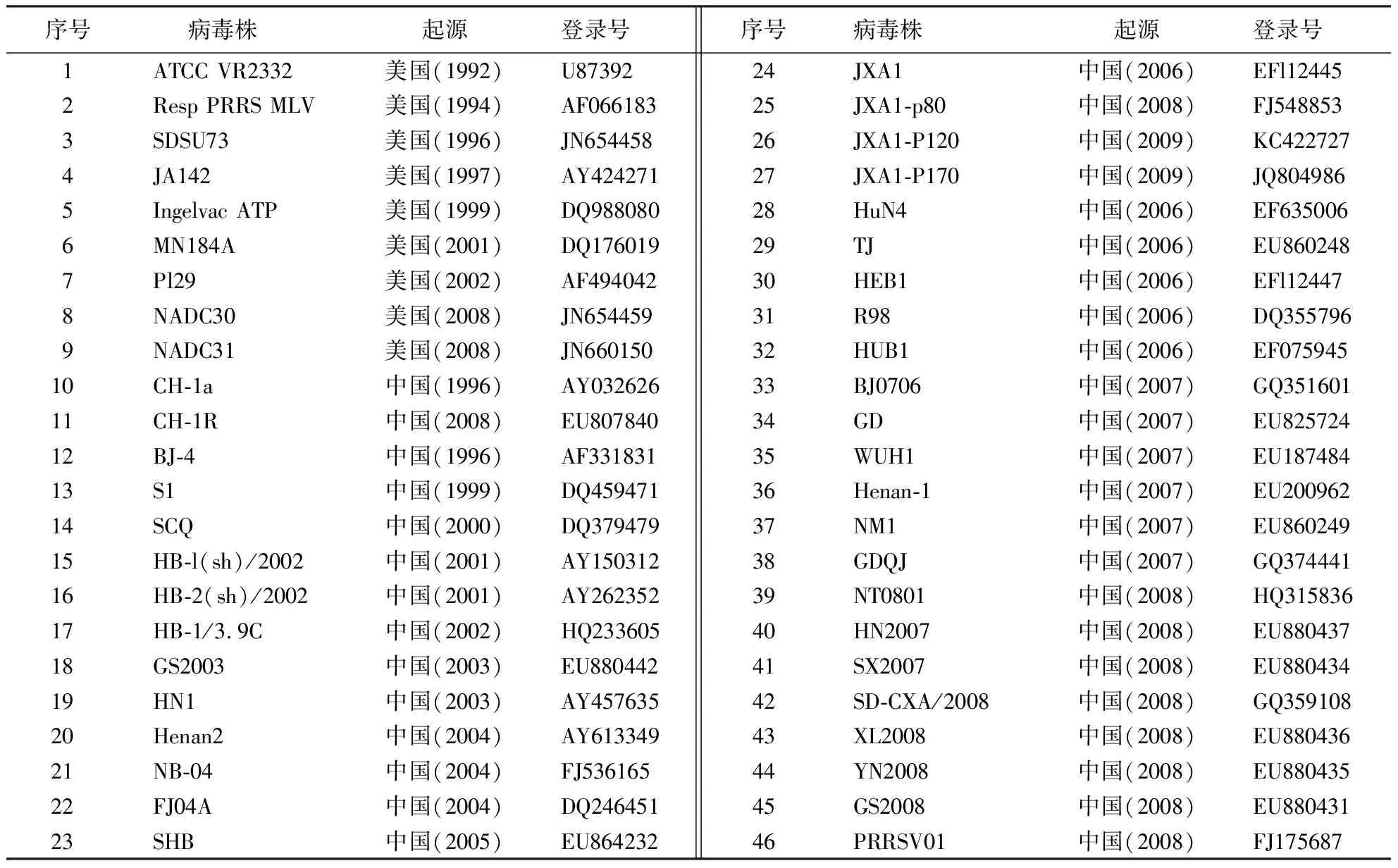
表2 参考毒株信息Table 2 Information of reference strains
2 结果与分析
2.1 组织和血清样品中PRRSV的qRT-PCR检测结果
应用qRT-PCR对来自闽粤赣地区的1 475份组织和血清样本cDNA进行病原检测。结果显示:2020、2021、2022年PRRSV的阳性率分别为24.23%(95/392)、27.69%(162/585)、18.07%(90/498);福建龙岩、泉州、漳州,广东汕头、梅州,江西赣州阳性率分别为23.20%(220/948)、11.36%(10/88)、31.11%(42/135)、24.50%(37/151)、22.73%(15/66)、26.44%(23/87);血清与组织样本阳性率分别为21.42%(231/1 078)、29.21%(116/397)。表明PRRSV广泛流行于不同地区,其中,漳州地区感染PRRSV的情况最为严重,阳性率高达31.11%(42/135)。
2.2 PRRSV OFR5基因的RT-PCR扩增与鉴定
挑选86份来自不同猪场的PRRSV阳性样品,对其ORF5基因进行扩增、测序,经NCBI比对验证,成功获得了62株PRRSVORF5基因序列。MegAlign软件分析显示,80株PRRSVORF5基因间核苷酸序列同源性为81.2%~100%,氨基酸序列同源性为78.6%~100%。80株PRRSVORF5基因序列与NADC30-like株、NADC34-like株等为代表的谱系1毒株间的核苷酸序列同源性为82.1%~95.0%,氨基酸序列同源性为81.6%~95.5%;与GM2-like株、QYYZ-like株等为代表的谱系3毒株间的核苷酸序列同源性为81.1%~94.5%,氨基酸序列同源性为80.1%~95.5%;与VR2332-like株、BJ-4-like株等为代表的谱系5毒株间的核苷酸序列同源性为82.1%~99.8%,氨基酸序列同源性为78.1%~98.5%;与JXA1-like株、CH-1a-like株等为代表的谱系8毒株间的核苷酸序列同源性为82.4%~95.0%,氨基酸序列同源性为80.1%~99.0%。
2.3 PRRSV ORF5基因遗传进化树的构建
为进一步分析闽粤赣地区PRRSV主要流行毒株的变异情况,利用Mega 11.0软件对80株PRRSVORF5基因序列进行系统发育树的构建。结果(图1)显示,80株PRRSV均属于美洲型,且分布在谱系1、3、5、8上。其中:40株PRRSV分布在谱系1上,与NADC30-like株属于同一分支;12株PRRSV分布在谱系3上,与QYYZ-like株属于同一分支;7株PRRSV分布在谱系5上,与VR2332-like株属于同一分支;21株PRRSV分布在谱系8上,与JXA1-like株属于同一分支。表明闽粤赣地区2020—2022年的流行毒株以谱系1为主。

●为本研究所扩增的PRRSV ORF5基因。
2016年以来以NADC30-like株为代表的谱系1在检出数量上已明显超过以HP-PRRSV株为代表的谱系8,表明NADC30-like株已取代HP-PRRSV株成为我国主要的流行毒株[8]。福建省多个地区还相继报道该亚型新出现的分支NADC34-like株[10],同时有研究表明NADC34-like株具备与NADC30-Like株类似的在我国流行的条件,因此需要加大监测力度[11]。
2.4 PRRSV ORF5基因编码的氨基酸序列
对闽粤赣地区的80株PRRSVORF5基因进行氨基酸序列比对,结果(图2)显示,ORF5基因编码200个氨基酸,没有氨基酸的插入与缺失。GP5蛋白的氨基酸序列突变主要集中在信号肽区(第1~31氨基酸残基处)和高变区(第32~35、57~61氨基酸残基处),在第62~120、130~160氨基酸残基处相对保守。有研究发现,第13、151氨基酸位点为推导的潜在毒力相关位点[12]。谱系8中仅有部分毒株在2个位点发生了突变,其中,1株发生了R13Q的突变,7株发生了第151位点的变异,1株发生了R151T的突变,2株发生了R151G的突变,3株发生了R151I的突变,其余15株保持了与CH-1a、JXA1等强毒株一致的序列特征。谱系3中有12株PRRSV在中和表位区发生了L39S的变异,谱系8中有20株PRRSV发生了L39I的变异。PRRSV疫苗株和野毒株可根据GP5蛋白上的第137位氨基酸进行区分[12],谱系5中PRRSV GP5蛋白第137位氨基酸与疫苗株Resp PRRSV MLV一样均为A,表明处于谱系5的7株PRRSV可能都来源于疫苗株。
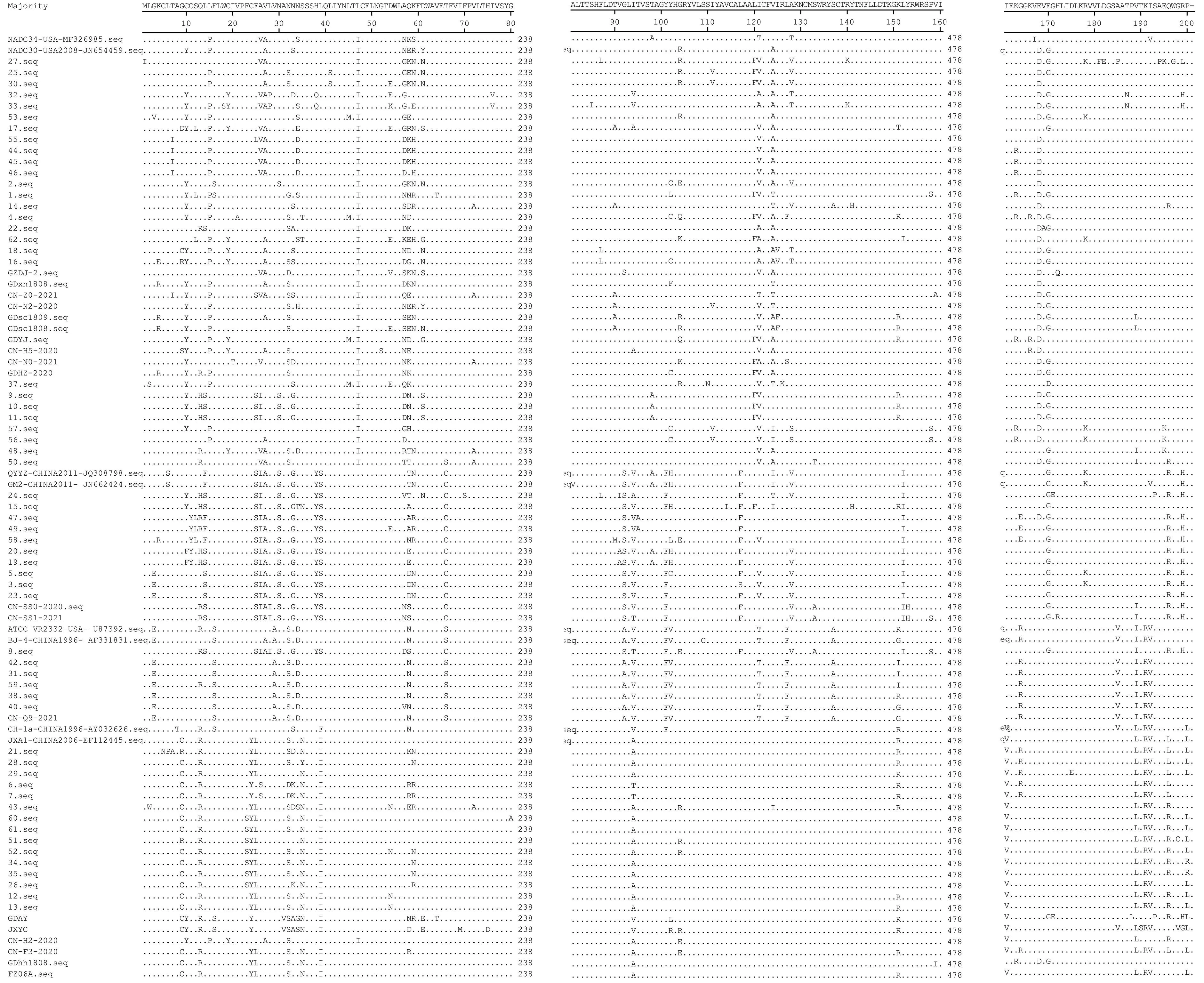
图2 PRRSV GP5蛋白氨基酸序列的比对Fig.2 Amino acid sequence alignment of PRRSV GP5 protein
3 讨论
PRRSV在我国持续性大规模暴发并导致猪群的高死亡率[5]。1996年郭宝清等[13]首次在国内暴发的繁殖与呼吸综合征猪群流产胎儿中分离到PRRSV,确认我国猪群存在PRRSV感染。2006年HP-PRRSV株对我国养猪业造成了巨大危害,因PRRSV基因组不断发生变异,自2016年起NADC30-like株逐渐替代HP-PRRSV株成为我国的主要优势毒株[7]。当前国内流行的PRRSV基因型种类复杂,一猪场可存在多种基因型PRRSV毒株,加剧了各基因型PRRSV毒株之间的重组变异情况,给猪繁殖与呼吸综合征的防控带来了巨大挑战。因此实时了解PRRSV流行株分子变异情况十分必要。
本研究对2020—2022年闽粤赣地区规模化猪场的1 475份病料样品进行PRRSV病原检测,共检测出347份PRRSV阳性样品,阳性率为23.5%,表明闽粤赣地区均能检测到PRRSV,虽在检出率上有所差异,但能说明各地区感染PRRSV已十分普遍;纵向观察显示,PRRSV阳性率从2021年的27.69%下降至2022年的18.07%,其大幅度下降可能与中华人民共和国农业农村部取消猪瘟病毒疫苗和PRRSV疫苗强制免疫计划有关,从而促使养殖场更加注重科学免疫以降低疫病对猪只的感染风险;从分布地区上发现,漳州市PRRSV阳性率占比最高,提示应对该地区加强猪繁殖与呼吸综合征的防控,以降低疫病带来的经济损失。自1996年PRRSV首次在我国报道后,迅速传播至各个省份,对全国养猪业造成巨大的经济损失。如:李文忠等[14]报道,2021—2022年天津地区养殖场疑似发病猪PRRSV阳性样品检出率为37.3%;吴瑕等[15]报道,2017—2020年江西省PRRSV阳性率为42.55%;高明艳等[16]报道,2021年河北省部分地区PRRSV阳性率为48.39%。经调查,近年来全国各地PRRSV阳性率仍处于较高水平,且PRRSV极易变异的特性给我国猪繁殖与呼吸综合征的防控带来了严峻的挑战。
基于PRRSV表面的主要囊膜蛋白GP5存在遗传多样性的特点,本研究对闽粤赣地区所检出的阳性PRRSV样品ORF5基因进行扩增及遗传进化分析。结果显示,闽粤赣地区PRRSV存在谱系1、3、5、8的混合流行,其中以NADC30-like株和NADC34-like株为代表的谱系1占比最高,为50.00%。80株PRRSVORF5基因间的核苷酸和氨基酸序列同源性分别为81.3%~100%、78.6%~100%;与谱系1毒株间的核苷酸序列同源性为82.1%~95.0%,氨基酸序列同源性为81.6%~95.5%。研究发现,2016—2018年PRRSV NADC30-like株已成为我国华北和西南地区的主要流行毒株(分别占比52.83%[17]、70.3%[18])。此结果与本研究结果高度一致,表明目前NADC30-like株仍然是我国PRRSV的优势流行株,因此亟需加强NADC30-like株的防控。
PRRSV疫苗株和野毒株可根据GP5蛋白的第137位氨基酸位点进行区分,而强、弱毒株可根据第13、151位氨基酸位点进行区分[12]。本研究对80株PRRSVORF5基因进行氨基酸序列比对,发现63株PRRSVORF5基因编码的第137位氨基酸位点均为S,与CH-1a-like株、JXA1-like株、NADC30-like株、GM2-like株相同;谱系5中7株PRRSVORF5基因编码的第137位氨基酸位点与疫苗株Resp PRRSV MLV一样均为A,表明其可能为疫苗株;谱系8中大部分毒株保持了强毒株的序列特征。此外,本研究还发现33株PRRSV存在L39I/L39S的变异,这一突变可能会影响GP5蛋白的免疫原性从而大幅提高病毒逃避疫苗免疫诱导中和作用的概率,进而降低疫苗免疫保护的效率[15,19],给猪繁殖与呼吸综合征的综合防控带来困难。因此,只有实时监测和掌握PRRSV的变异情况及流行动态,才能有效预防猪繁殖与呼吸综合征疫情的发生。
猜你喜欢
艺术品鉴(2022年16期)2022-07-09
科学大观园(2022年2期)2022-01-23
河池学院学报(2021年1期)2021-07-10
景德镇陶瓷(2021年1期)2021-03-24
趣味(数学)(2020年4期)2020-07-27
支部建设(2020年15期)2020-07-08
东方考古(2017年0期)2017-07-11
动物医学进展(2015年10期)2015-12-07
百科知识(2015年18期)2015-09-10
特产研究(2014年4期)2014-04-10
