
大别班达病毒的遗传进化机制研究
2024-03-15 00:59张宇涵王玉昊韦雪敏许一菲
中国人兽共患病学报 2024年2期
张宇涵,李 超,王玉昊,韦雪敏,许一菲,2
大别班达病毒(Dabie bandavirus, DBV)是一种新型的布尼亚病毒,2009年由于学杰等人在中国首次发现并分离鉴定[1]。DBV主要通过蜱虫叮咬传播给人类,引起发热伴血小板减少综合征(SFTS),临床表现为高热、血小板和白细胞计数减少等症状,并伴有中枢神经症状、胃肠道症状等[1]。SFTS的地方性散发流行期为4-10月。2011-2021年,SFTS年平均病死率为5.11%[2]。SFTS在中国主要流行于河南、湖北、山东、浙江、辽宁等省份[3-4]。
DBV属于Bunyavirales目,Phenuiviridae科,Bandavirus属。DBV是单股负链分节段的RNA病毒,包含L、M、S 3个片段,分别编码RNA依赖的RNA聚合酶(RdRp)、糖蛋白(Gp)、非结构蛋白(NSs)和核蛋白(NP)[5]。参与病毒的复制与转录的L基因编码的RdRp蛋白,是最保守的蛋白,进化速率也较慢;S基因编码的NSs蛋白主要参与宿主的免疫调节,其系统发育最为多样,是保守性最低的蛋白,进化速率也较快[6]。可根据地理位置和系统发育关系,将DBV划分为主要包括中国毒株的中国谱系(C1-C5),主要包括日本和韩国毒株的日本谱系(J1-J3)[6,8]。此外,两个潜在的亚谱系C6和J4在2017年的一项研究中得到了鉴定[9]。此前有研究表明,虽然中国和日本地区的DBV毒株可能源自同一祖先,但他们的亲缘关系较远,这暗示着很早以前,两个地区的毒株就已开始各自独立进化[8]。重配是分节段病毒特有的遗传进化事件,当多株不同型别的病毒感染同一宿主时,其RNA片段会进行混合,从而产生新特性的病毒。重组和重配事件会对病毒的传播力和致病性产生影响,是病毒进化的潜在动力[6,10]。选择压力指的是外界环境在生物进化过程中对其进化方向产生影响的作用力。病毒的进化选择压力来自于宿主的免疫压力和生态环境的变化。病毒容易通过自身的突变来逃避宿主免疫压力或适应新的环境,继而发生适应性变化,导致免疫防治或者药物治疗效果不佳。
本研究对截至2023年7月已发布的数据库全部的DBV各片段全长基因序列进行系统发育分析、基因重组和重配分析以及选择压力分析,阐明DBV的基因型流行分布特征,探究DBV进化机制的动态变化,为DBV的预防控制提供科学依据。
1 材料与方法
1.1 数据来源 在VIPR数据库和GenBank数据库下载DBV全长序列(下载于2023年7月),并记录序列分离时间、地点,以及样本类型。其中,L片段序列1 407条,M片段序列1 424条,S片段序列1 598条,构建以上3个数据集。在Refseq(https://www.ncbi.nlm.nih.gov/refseq/)下载毒株HB29为参考株(基因序列登录号:NC_018136.1-018138.1)。
1.2 方法
1.2.1 系统发育分析 对3个数据集的序列分别进行系统发育分析。使用MAFFT软件进行多序列比对,使用Iqtree软件以最大似然法分别构建基于L、M、S片段的系统发育树(bootstraP=1000)[11],并利用R软件包ggtree对获得的最大似然树进行可视化和修饰[12]。
1.2.2 重组和重配分析 本研究运用RDP4(Recombination Detection Program4)软件对1.1提及的3个数据集包含的所有序列中的重组事件进行检测。经RDP4内置的7种算法(RDP[13]、Bootscan[14]、himaera[15]、Seq[16]、GENECONV[17]、Lard、SiScan)检测,序列被4种以上方法检测为重组阳性且P< 0.05时,即被认为是潜在重组事件。
基于L、M、S 3个片段的系统发育分析结果,在含有DBV全长基因组的1 025株毒株中,筛选含有不同基因型的节段的潜在重配毒株。
1.2.3 选择压力分析 使用Bioedit进行L、M、S片段的核苷酸和氨基酸同源性分析,使用Mega11.0软件的Pairwise distance功能,选择p-distance模型,分别计算3个片段的遗传距离区间。
使用Mega11.0软件的Nei-Gojobori method(No.of Differences)模型分别计算L、M、S 3个片段的dN和dS的值。当dN/dS> 1时,认为发生正向选择作用;当dN/dS< 1时,认为发生负向选择作用。
2 结 果
2.1 DBV系统发育分析 L、M、S 3个片段的进化树结果显示,DBV可分为9种基因型别:C1-C6和J1-J3。L、M片段比S片段多出C6型。3个片段中属于C2、C4型的序列数量最多,C2、C3型涵盖的地域范围最广,J1、J2型包含的宿主类型最丰富。中国的DBV序列主要分布在C谱系,其中河南省和湖北省的优势基因型是C2,其次是C4型,山东省的优势基因型为C3型。日本、韩国的DBV序列以J谱系为主,且优势基因型均为J1型。同时,J1型的浙江部分毒株和日本毒株在同一个支系,J2型的河南部分毒株和韩国毒株在同一支系。

注:ABC分别代表系统发育进化树的L、M、S片段。
2.2 DBV重组和重配结果 经RDP4软件筛选后,在本研究数据集中发现88株毒株中存在98个潜在重组事件(表1),L、M、S片段分别发现49、41、28个重组事件。其中,L片段的重组率最高,为3.48%(49/1 407),M片段的重组率为2.88%(41/1 424),S片段重组率最低,为1.75%(28/1 598)。毒株2011YXX9的3个片段均发生了重组,另有11株毒株的两个片段发生了重组,如毒株13-China_Henan-275、毒株HNXY_31、毒株NB32/CHN/2013等。此外,在10株毒株中发现了存在多次重组事件的节段,例如毒株2011YSC60、毒株HB2012-171、毒株2011YXX9等。上述结果表明L片段发生重组的概率较高,S片段发生重组的概率较低。人类样本发生重组事件的数量较多(74/88),蜱虫和家畜样本较少(10/88)。L、M、S片段中,人类宿主检测到重组的概率分别为2.5%、2.9%、1.8%,动物(蜱虫和家畜)宿主检测到重组的概率分别为5.9%、3.0%、5.1%。在非人类宿主中检测到重组事件的比例高于人类宿主,且相较之前的研究(各片段均0.99%)[18]有所增加。
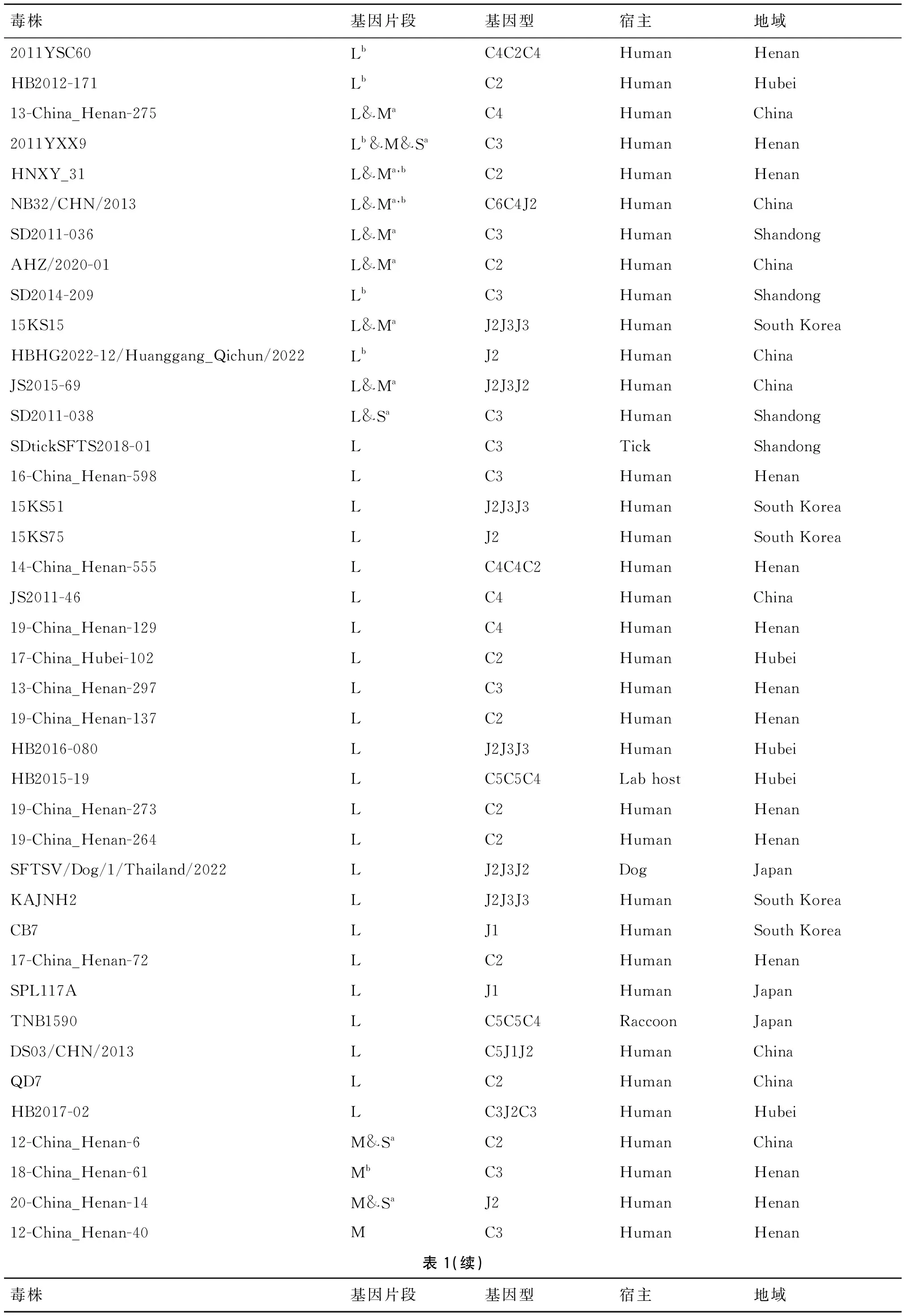
表1 DBV发生重组的毒株统计
根据L、M、S三个片段的系统发育分析结果,在148株重配株中发现了36种重配基因型(图2),其中20种类型已被先前的研究报道过[6-9]。主要的重配类型为C4C4C2 (32/148)、J2J3J3 (16/148)和C2C1C2(10/148)。绝大部分重配毒株发现于人源样本(133/148),有9株来源于家畜(5/148)、蜱虫(3/148)和小型哺乳动物(1/148),此外还有5株来源于实验室宿主。同时,大部分动物宿主来源的重配株均来自日本(3/9)。

图2 DBV 36种重配株的组合图Fig.2 Genetic constellation of the 36 potential DBV reassortants
同时,我们发现24株同时发生重组和重配的毒株(表1)。其中有两株毒株(SFTSV/Dog/1/Thailand/2022、TNB1590)分离自动物宿主。
2.3 DBV基因同源性及遗传距离分析 DBV毒株间表现出较高的同源性,L、M、S片段的核苷酸同源性分别为94.8%~99.9%,91.8%~99.9%,88.8%~99.9%;RdRp基因、Gp基因、NS基因、NP基因的氨基酸同源性分别为97.9%~100%,92.7%~100%,79.6%~99.6%,86.9%~100%(表2)。

表2 DBV基因核苷酸和氨基酸同源性结果
DBV 3个基因的总体遗传距离分别为0.005、0.015和0.047,L片段的遗传距离区间为0.00~0.02,M片段的遗传距离区间为0.00~0.06,S片段的遗传距离区间为0.00~0.11(表3)。
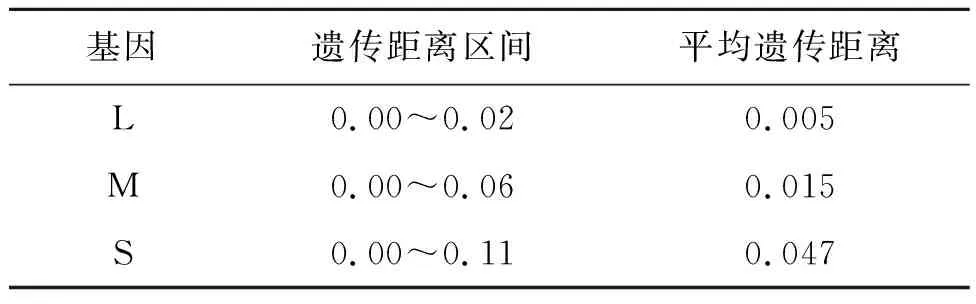
表3 DBV毒株遗传进化距离分析结果
结果表明S片段的核苷酸和氨基酸同源性均低于L、M片段,与其他两者相比,遗传距离也较大。
2.4 DBV选择压力分析 通过Mega 11.0软件对DBV编码的4个蛋白进行了全局dN/dS分析(如表4所示),结果显示,4个蛋白的dN/dS值均小于1,表明在进化过程中,4个蛋白均受到负选择压力的作用。而与L基因、M基因和NP蛋白相比,NS蛋白的dN/dS值最高为0.18。

表4 DBV蛋白的dN/dS比值结果
3 讨 论
由DBV感染引发的SFTS是我国目前急需解决的公共卫生问题,它是一种易诱发多种临床病症且病死率较高的蜱媒传染病,目前尚无获得临床批准的SFTS特效抗病毒药物,支持治疗是SFTS患者的主要临床治疗方式[19-21]。自最初发现以来,DBV已在中国、日本[22]、韩国[23]等超过25个国家被发现,在美国[24]和澳大利亚[25]也相继发现了一些新白蛉病毒与DBV存在着密切进化关系。2017年,DBV被世界卫生组织(WHO)列为十大最受关注的病原之一[26]。该研究通过测定公共数据库的全部DBV基因组序列,对DBV的遗传进化从全序角度进行了深入系统的研究。
本研究验证了绝大多数DBV型别的分布具有地域性,在DBV的两大谱系中,C谱系主要包括中国毒株,J谱系主要包括韩国和日本毒株[27]。小部分毒株的基因型存在跨地域分布,Yun Y等将中日韩间的DBV基因型分布与候鸟迁徙路线结合分析,认为候鸟可能在DBV的跨地域传播中发挥着重要作用[6,28]。
本研究进一步证实了先前研究报道过的重组情况[28-29],并在人类宿主与动物宿主分离出的DBV毒株中均发现重组的现象,且动物宿主的重组率高于人类宿主。大部分重组DBV毒株是来自人源样本,而小部分动物宿主来源的毒株也作为亲本序列促进或参与了重组的发生,说明动物宿主也提供了进行重组的场所。在蜱虫、狗、猫、刺猬等不同动物宿主来源的毒株中都发现DBV重组现象,并且近年来动物宿主发生重组的比例在增加,这表明DBV可以发生广泛的跨物种传播和感染。本研究发现了16种未被报道过的重配型别,提示DBV的毒力和流行传播能力正在发生改变,此发现对疾病的控制具有重要意义。重配事件所涉及的不同的地域,表明DBV可能在不同的地域之间进行迁移。蜱虫曾被认为可能是适合DBV重配株的来源[26],本研究中重配株的宿主多样性提示,除蜱虫外,家畜或伴侣动物也可能会加强DBV重配现象的产生。越来越多的动物宿主如狗、猫、牛等参与DBV的重组、重配事件中,使其进化机制的丰富度不断提高,这对DBV疫苗的设计和SFTS的预防产生了更大的挑战。
基因组全编码区的选择压力分析对其疫苗和药物的开发具有较大的指导意义。本研究结果表明影响DBV进化的主要选择压力是负选择压力,这一结论与以往的研究一致[30]。4个基因的dN/dS值均小于1,其中NS基因的dN/dS值高于其他3个基因,这可能与NS基因编码的蛋白有关。NSs能够通过拮抗干扰素而影响毒力,进而帮助病毒逃避宿主的抗病毒反应,从而参与宿主的免疫调节[9]。NSs为保守性最低的蛋白,其进化速率也较快。相比于其他两个片段,NS基因较低的同源性和较远的平均遗传距离进一步验证了NSs基因片段具有较高突变能力。由于蛋白中的氨基酸变化可能影响其功能,进而影响DBV的感染性和致病力[6],基因型特异性的突变位点是否存在还需要进一步被探索。
利益冲突:无
引用本文格式:张宇涵,李超,王玉昊,等.大别班达病毒的遗传进化机制研究[J].中国人兽共患病学报,2024,40(2):171-178. DOI:10.3969/j.issn.1002-2694.2024.00.027
猜你喜欢
科学大观园(2022年2期)2022-01-23
科学(2020年3期)2020-11-26
当代水产(2020年3期)2020-06-15
实用皮肤病学杂志(2015年4期)2015-12-22
动物医学进展(2015年10期)2015-12-07
医学研究杂志(2015年12期)2015-06-10
现代检验医学杂志(2015年6期)2015-02-06
特产研究(2014年4期)2014-04-10
郑州大学学报(理学版)(2014年3期)2014-03-01
实验动物与比较医学(2014年5期)2014-02-28
