
Trehalose: A sugar molecule involved in temperature stress management in plants
2024-03-07 01:51:36AlRzSvtBrwAtkurRmnProGrcCprrHbbFslSrCrCrstnFoyrKmbotSquRjvVrsny
The Crop Journal 2024年1期
Al Rz, Svt Brw M Atkur Rmn, Pro Grcí-Cprrós, M Hbb,Fsl S, Sr Cr, Crstn H.Foyr, Kmbot H.M.Squ*, Rjv K.Vrsny*
a College of Agriculture, Fujian Agriculture and Forestry University (FAFU), Fuzhou 350002, Fujian, China
b Department of Botany, School of Bioengineering and Biosciences, Lovely Professional University, Phagwara 144411, India
c Department of Botany, MCM DAV College, Kangra, Himachal Pradesh 176001, India
d Grassland and Forage Division, National Institute of Animal Science, Rural Development Administration, Cheonan 31000, Republic of Korea
e Agronomy Department of Superior School Engineering, University of Almería, Almería, Spain
f National Institute for Genomics and Advanced Biotechnology, National Agricultural Research Centre, Park Rd., Islamabad 45500, Pakistan
g Agriculture, Health and Environment Department, Natural Resources Institute, Faculty of Engineering & Science, University of Greenwich, Kent ME4 4TB, UK
h State Key Laboratory of Rice Biology, China National Rice Research Institute, Chinese Academy of Agricultural Sciences, Hangzhou, Zhejiang, China
i School of Biosciences, College of Life and Environmental Sciences, University of Birmingham, Edgbaston B15 2TT, UK
j The UWA Institute of Agriculture, The University of Western Australia, Crawley, Perth 6009, Australia
k WA State Agricultural Biotechnology Centre, Centre for Crop and Food Innovation, Food Futures Institute, Murdoch University, Murdoch, WA 6150, Australia
Keywords:Abiotic stress Gene expression Genetic engineering Osmolyte Trehalose-6-phosphate
ABSTRACT Trehalose (Tre) is a non-reducing disaccharide found in many species, including bacteria, fungi, invertebrates, yeast, and even plants, where it acts as an osmoprotectant, energy source, or protein/membrane protector.Despite relatively small amounts in plants, Tre concentrations increase following exposure to abiotic stressors.Trehalose-6-phosphate, a precursor of Tre, has regulatory functions in sugar metabolism, crop production, and stress tolerance.Among the various abiotic stresses, temperature extremes(heat or cold stress) are anticipated to impact crop production worldwide due to ongoing climate changes.Applying small amounts of Tre can mitigate negative physiological, metabolic, and molecular responses triggered by temperature stress.Trehalose also interacts with other sugars, osmoprotectants,amino acids, and phytohormones to regulate metabolic reprogramming that underpins temperature stress adaptation.Transformed plants expressing Tre-synthesis genes accumulate Tre and show improved stress tolerance.Genome-wide studies of Tre-encoding genes suggest roles in plant growth,development, and stress tolerance.This review discusses the functions of Tre in mitigating temperature stress—highlighting genetic engineering approaches to modify Tre metabolism, crosstalk, and interactions with other molecules—and in-silico approaches for identifying novel Tre-encoding genes in diverse plant species.We consider how this knowledge can be used to develop temperature-resilient crops essential for sustainable agriculture.
1.Introduction
The steady increase in global greenhouse gases,particularly carbon dioxide(CO2),has increased land and sea surface temperatures(‘global warming’).Climate change, characterized by long-term shifts in temperature and weather patterns, has brought about an increased frequency of heatwaves, severe cold spells, and drought periods[1,2].While climate change is a global phenomenon,some regions experience its effects more intensely (Fig.1), leading to various environmental stresses for agriculture (Fig.1A).The escalating temperatures caused by global warming and climate change(Fig.1B) adversely impact agriculture and crop productivity [1,2].According to current climate models (Fig.1C), mean global warming in the 21st century could exceed the Last Glacial Maximum and perhaps be even the hottest in the Holocene,consequently threatening the lives of the 2.8 billion people in agricultural societies[3].The primary concern is the detrimental effects of these temperature changes on plant development and food production [2,3].According to the Intergovernmental Panel on Climate Change 2018 report,if current warming trends persist,global temperatures will rise by 1.5°C between 2030 and 2052[4].Thus,we need a deeper understanding of the relationships between crop productivity and increased growing temperatures to implement preemptive approaches and measures to prevent global food insecurity.
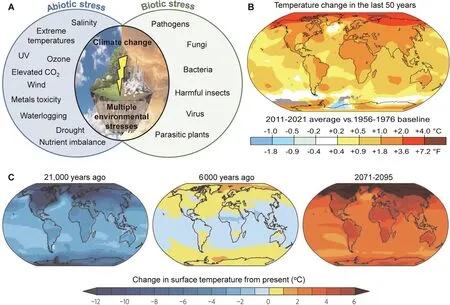
Fig.1.Effect of climate change and global warming on changes in environmental stress.(A) Current climate change events give rise to numerous environmental stresses,including abiotic and biotic factors,which could destructively affect crop yield and trigger food shortages.(B)Average surface air temperatures from 2011 to 2020 compared to 1951–1980.Source:NASA(https://data.giss.nasa.gov/gistemp/).(C)Model-simulated global temperature anomalies for the Last Glacial Maximum(21,000 years ago),mid-Holocene(6000 years ago),and forecast for 2071–2095.The forecast depicts the most severe case of possible climate change,with an anticipated world average temperature increase of 5–6°C by 2100 compared to pre-industrial conditions.There seems to be scientific agreement that unrestricted carbon pollution would result in global warming,with no less than several °C by 2100, posing significant dangers to human civilization and natural habitats at the urban, national, and international scales, impacting agricultural land and production worldwide.Source: Climate Change Knowledge Portal (https://climateknowledgeportal.worldbank.org/overview).
The sessile nature of plants requires tailoring their morphology,metabolism, and stress responses to withstand the adverse effects of climate change [2,5–9].Temperature extremes (i.e., cold stress(CS), such as chilling 0–15 °C and freezing < 0 °C, and heat stress(HS)>25°C)are major contributors to agricultural losses and a significant challenge to global food security [7,8,10,11].Temperature stress, alone or combined with other stresses, increases cellular oxidation and related signaling, leading to growth cessation and even programmed cell death[2,3,5].Heat and cold stresses trigger prolonged oxidation,adversely affecting cellular redox metabolism and triggering genetic pathways that limit crop growth[7,8,10,12,13].Extreme temperatures increase the production of reactive oxygen species(ROS),such as singlet oxygen(1O2),hydroxyl(OH•)radicals,superoxide(O2•-),and hydrogen peroxide(H2O2)[14].While ROS have the chemical reactivity to damage cellular components, current understanding suggests that their accumulation restricts plant development through transcriptional and posttranslational controls that regulate cell fate in response to temperature stress [8,14–17].
At the morphological level,CS and HS decrease crop productivity by causing shoot burning,stunted root and shoot development,leaf shedding,and fruit discoloration[2,5,8].Moreover,these stresses inhibit crucial physiological processes like photosynthesis,antioxidant activity,and nutrient uptake[2,5,8,11,18].By decreasing membrane fluidity and antioxidant enzyme activities, HS and CS disrupt the production of various proteins and secondary metabolites and perturb hormone signaling, ultimately causing cellular damage or cell death [10,19–21].HS also affects protein homeostasis by influencing protein synthesis,folding,quality control, and subcellular localization [22], while CS increases cellular membrane permeability and reduces unsaturated fatty acid contents [23], further diminishing plant efficiency.
Plants have evolved an intricate network of cellular stress response mechanisms to counteract the harmful effects of extreme temperatures,including repairing disrupted proteins by regulating stress-responsive genes and producing heat shock proteins(HSPs)/chaperones [24,25].The primary drivers of transcriptional regulation in plants under CS and HS include the upregulation of HSP/signaling pathways,ROS signaling,and the mitogen-activated protein kinase transduction pathway [25–27].Heat shock transcription factors (HSFs) are key components of the complex transcriptional regulatory network governing the heat stress response (HSR) in plants, as they activate downstream genes encoding HSR-induced transcription factors, metabolic enzymes, and HSPs [25,28,29].Changes in gene expression are a major outcome of extreme temperatures, leading to a decline in plant productivity [7,30].Plants also have a memory of adverse experiences, manifesting as transcriptional, somatic, intergenerational, and transgenerational stress memory[31,32],mediated by epigenetic changes regulating gene regulation expression in response to CS and HS [30,33,34].
Plants employ a comprehensive network of defense mechanisms to mitigate the harmful effects of stress[1,35].Stress modifies nearly every aspect of plant biology, including the synthesis and aggregation of osmolytes(e.g.,proline,glycine betaine,mannitol, glucose, sorbitol, fructose, and trehalose) [18,36–42].In addition, plants have a range of stress-responsive phytohormones that preserve organelle functions, boost antioxidant defenses, and mediate osmotic adjustments to prevailing environmental conditions [11,18,36–42].
Of the different plant osmolytes(mainly sugars),trehalose(Tre,a disaccharide and ‘sugar of life’) was first reported in rye (Secale cereale) and fungi (Claviceps spp.) [43–45].Trehalose has since been reported in many bacteria, fungi, invertebrates, and plants and is frequently associated with stress-smart phenotypes(i.e., traits that empower plants to successfully adapt and respond to diverse environmental factors such as temperature stress)[45–48].Trehalose protects cellular integrity in plants subjected to abiotic stress (e.g., heat, cold, salt, and drought) [48–50].However, precisely how Tre (or its precursor trehalose-6-phosphate,T6P)regulates stress management,particularly under temperature stress, requires further investigation.Trehalose stabilizes dehydrated enzymes (that have confronted reduced water levels because of environmental stresses like temperature or drought),lipid membranes,and proteins in plants subjected to abiotic stress[45], maintaining their structural integrity and functional movement[45].T6P is likely the more critical component,with Tre possibly functioning as a ROS mitigator rather than a profound regulator of resource allocation,as T6P does.Additionally,T6P triggers the expression of antioxidant systems that prevent excessive ROS accumulation during stress [48–51].Thus, exogenous Tre application has emerged as a promising strategy to increase endogenous Tre levels and enhance plant stress tolerance[49–51].Trehalose treatments improve stress tolerance by regulating physiological, biochemical, and molecular processes [48–51].
Trehalose is an osmolyte,energy house,and signaling molecule that coordinates adaptive temperature stress responses [48–52].Intriguingly, plants appear to have specific molecular sensors that enable them to detect changes in Tre levels under extreme temperature conditions.For instance, studies have reported that plants triggered explicit molecular pathways in response to elevated Tre levels [49–53].Trehalose-6-phosphate synthase (TPS) enzymes,involved in Tre biosynthesis, can be induced by CS, leading to increased Tre levels [54–56].Consequently, elevated Tre levels can serve as a signal to activate downstream cold-responsive genes,enhancing CS tolerance[51,54,55,57,58].Under HS,elevated Tre levels can trigger HS response pathways.Particularly noteworthy is the connection between Tre accumulation and pathway activation (e.g., protein kinase C1 (PKC1) pathway), highlighting how plants can detect Tre as a signal of impending stress, even in the absence of the primary stress factor [59].This ability of plants to sense Tre as an indicator of forthcoming stress demonstrates its versatile role in orchestrating cellular defenses against HS [59].Incorporating Tre sensitivity into the stress response system showcases the remarkable adaptability of plants to extreme temperatures.Understanding how plants recognize Tre as a signal for stress response provides novel insights into the intricate regulatory mechanisms that reinforce their capacity to thrive in challenging temperature conditions.Therefore, the following section explores how Tre application can improve crop yield sustainability under temperature stress.We propose that genome-wide identification and genetic engineering of Tre-encoding genes, Tre crosstalk, and interactions with other molecules can improve Tre metabolism and, hence, temperature stress tolerance.
2.Trehalose biosynthesis and metabolism
The chemical structure of Tre (α-D-glucopyranosyl-[1,1]-α-Dglucopyranoside) comprises two glucose monomers linked by an α,α-1,1′-O-glycosidic bond.Except for vertebrates, all kingdoms produce this non-reducing disaccharide [60], which is chemically inserted with high hydrophilicity and very low bond energy(4.2 kJ mol-1) [49].Trehalose has a molecular mass of 342 Da,whereas the phosphorylated form, trehalose-6-phosphate (T6P),has a molecular mass of 422 Da [61].Trehalose is resistant to hydrolysis and degradation at elevated temperatures [54].

Fig.2.Trehalose metabolism in plants.G6P,glucose-6-phosphate;Pi,inorganic phosphate;TPS,trehalose-6-phosphate synthase;T6P,trehalose-6-phosphate;TPP,trehalose-6-phosphate-phosphatase; Tre, trehalose; UDPG, uridine diphosphate glucose; UDP, uridine diphosphate.
Trehalose biosynthesis in plants is catalyzed in a two-step reaction pathway involving trehalose-6-phosphate synthase (TPS) and trehalose-6-phosphate-phosphatase (TPP), localized in the cytosol(Fig.2).These enzymes catalyze the condensation reaction of uridine diphosphate glucose (UDPG) and glucose-6-phosphate (G6P)to T6P and the subsequent dephosphorylation of T6P to Tre.Finally,Tre is hydrolyzed by trehalase (TRE) into two glucose (Glu) molecules [62].Trehalose breakdown via Tre phosphorylase can also generate Glu and glucose-1-phosphate [63,64].Together, these enzymes generate signaling molecules by regulating the different physiological states of Tre homeostasis [65].Large gene families encode TPS and TPP in plants.For example, Arabidopsis thaliana has 11 TPS genes and 10 TPP genes [66].The Tre, TPP, and T6P levels in different cell types depend on the relative abundance of Tre and TPS expression and enzyme activity [53].
Other Tre-synthesis pathways occur in nature.While the TPS–TPP pathway occurs in higher plants, other organisms (including some plants),such as Escherichia coli,use T6P synthase(also known as OtsA) to transfer Glu from UDP-glucose to G6P to generate the intermediate T6P; then, TPP (OtsB in E.coli) dephosphorylates T6P to Tre [45].A different Tre pathway (TreZ–TreY), reported in thermophilic archaea of the genus Sulfolobus, involves the conversion of maltooligosaccharides or starch to Tre, catalyzed by maltooligosyl trehalose synthase (TreY) and maltooligosyl trehalose trehalohydrolase (TreZ) [67].This TreT pathway occurs in many extremophiles, including Thermococcus litoralis and Thermotoga maritima, where trehalose glycosyl-transferring synthase (TreT)catalyzes the synthesis of Tre from ADP-glucose and Glu [68].
Trehalose and T6P are present in low levels in plants, with reported values of<10 μmol g-1FW or 10 nmol g-1FW,depending on the species [69].These metabolites accumulate under abiotic stress conditions, especially in desiccation-tolerant plants [70].However, the accumulation level depends on the type of stress.T6P is localized predominantly in the cytoplasm (71%–72%) of A.thaliana leaves,with lower amounts reported in the chloroplasts(16%–22%) and vacuoles (7%–11%) [71].The accumulation of T6P depends on the relative rates of synthesis(TPS) and dephosphorylation (TPP), which, in turn, are regulated by transcriptional and post-translational controls linked to cellular sucrose (Suc) status[72].TPS enzymes are phosphorylated by SNF1-related protein kinase (SnRK1), with sophisticated feedback control, such as T6P inhibiting SnRK1.Class II TPSs,in particular,play a vital role in regulating plant growth and development [73], inhibiting SnRK1.Development and cell type are also crucial factors in determining T6P levels and responses to T6P [74,75].T6P acts as a Suc signal metabolite, detecting Suc availability to control the pathways of Suc anabolism and catabolism depending on local Suc availability[70].Hence,T6P:Suc ratios uniquely govern carbohydrate metabolism related to plant development and defense.Changes in T6P:Suc ratios within specific cell types are crucial for controlling plant productivity and yield [72].
3.Temperature stress management by trehalose application
3.1.Cold stress
Cold stress (i.e., chilling and freezing temperatures) adversely affects crop productivity and yield [5,7,10].Long-term CS causes ROS accumulation and perturbs cellular redox metabolism[11,51,76].Furthermore, CS decreases photosynthesis and mem-brane fluidity, adversely affecting crop productivity and, in some cases,causing programmed cell death[21].Trehalose accumulates in CS-tolerant crop plants such as Tripogon loliiformis,suggesting a positive role for this sugar in CS tolerance[77].The signaling function of Tre as a ROS mitigator and T6P as a signal is important in this regard (Table 1; Fig.3), influencing metabolic pathways that benefit survival under CS [50,72,78].Moreover, modulation of these pathways (Tre hydrolysis or T6P synthesis and hydrolysis)can improve crop yield and biomass production [49].

Table 1 Summary of key temperature stress management studies using exogenous trehalose application in diverse plant species.
3.1.1.Trehalose modulates plant growth and development under cold stress
Chilling stress affects plant growth and development, with reduced photosynthesis and restrictions in growth-regulating pathways contributing to the accumulation of excess carbon that cannot be used for growth, ultimately affecting plant productivity[5,21].For example,T6P regulated maize growth and development by improving carbon availability under CS [92].Similarly, priming rice (Oryza sativa L.) seedlings by supplying exogenous Tre (0.5, 1,and 2 mmol L–1) and spermidine (Spm) improved shoot parameters and seedling dry weight under CS(15°C)[93].Chickpea(Cicer arietinum L.) seedlings also exhibited improved growth under CS(13/10°C),associated with priming-mediated increases in Tre content [94].
Exogenous Tre application (10 mmol L–1) to tomato (Solanum lycopersicum L.) plants significantly alleviated CS-dependent inhibition of growth and biomass accumulation [57].The increased activities of enzymes associated with carbohydrate storage in photosynthetic tissues may explain the Tre-mediated increase in plant biomass production [95].Trehalose regulates ROS accumulation,turgor pressure, and mineral balance by activating molecular and metabolic components involved in these processes [78].A range of root parameters such as root surface area, length, and fresh dry weight increased in maize (Zea mays L.) seedlings following Tre application(12 mmol L–1)under CS(5±0.5°C)[92].Similarly,pre-treating wheat (Triticum aestivum L.) with Tre (1, 10, and 50 mmol L–1)significantly alleviated CS-induced decreases in grain number per spike by preventing floret degeneration and enhancing floret fertility in apical spikelets [96].
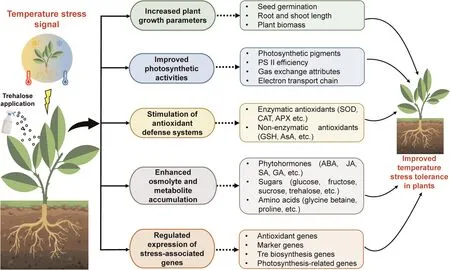
Fig.3.Diagrammatic representation of trehalose-mediated temperature stress alleviation in plants.APX, ascorbate peroxidase; CAT, catalase; PSII, photosystem II; SOD,superoxide dismutase; Tre, trehalose.
3.1.2.Trehalose improves photosynthesis and photosynthetic apparatus under cold stress
Cold stress impairs photosynthesis in many plant species [97].Exposure to CS decreases the amounts of photosynthetic pigments and disrupts chloroplast membrane and stromal functions,decreasing photosynthate production and altering Suc–sink relationships[98–100].Exogenous Tre application(10 mmol L–1)improved plant photosynthesis in melon (Cucumis melo L.) seedlings under CS,leading to increased net photosynthesis rates (Pn), transpiration(Tr), stomatal conductance (gs), and water use efficiency (WUE)[101].Moreover, Tre application regulated water absorption and use and improved the structural and functional stability of mesophyll cell chloroplasts [101].Exogenous Tre application(10 mmol L–1) in melon increased the maximal quantum yield of photosystem II(PSII)photochemistry under CS and decreased electrolyte leakage[80].Similarly,pepper(Capsicum annuum L.)plants had higher chlorophyll contents following Tre application(5%,10%,and 15%) under CS (4 °C) [102].Tre-mediated increases in photosynthesis observed under CS are attributed to various factors,including improved chlorophyll synthesis, activities of ribulose-1,5-bisphosphate (RuBP) carboxylase/oxygenase, PSII efficiencies,and antioxidant defenses[80,101,102].
3.1.3.Trehalose improves antioxidant defense systems and detoxifies ROS under cold stress
Cold stress causes prolonged ROS accumulation and profoundly influences redox regulation and signaling[8,10,11].ROS are oxidative signals that arise from primary and secondary metabolism and provide essential information on metabolism and the energy state of different cell compartments [14].In addition, stress-triggered apoplastic ROS burst (oxidative burst) is essential for cell-to-cell communication and forms the foundation of systemic stress responses called systemic acquired acclimation to abiotic stress[14].All ROS molecules have the potential to serve as signaling functions, with the signaling pathway depending on the presence of redox-regulated sensors and processing systems in each cellular compartment[14,103].Periods of high ROS accumulation in specific cellular compartments,particularly the apoplast,are required to transmit stress signals effectively.Each cellular compartment contains low molecular weight antioxidants such as ascorbate and glutathione (GSH), antioxidant enzymes, and ROS-processing enzymes that limit ROS accumulation and signaling [104–106].Ascorbate and GSH concentrations, antioxidant enzyme activities,and the abundance of ROS-processing enzymes are regulated in each intracellular compartment to allow appropriate regulation of the cell-to-cell ROS signaling wave and other redox signaling pathways.This antioxidant system serves to police ROS accumulation and associated changes in redox signaling [14,103,107] that regulate growth and defense[16,17],particularly under stress conditions [14,15].Plants often synthesize antioxidant enzymes [superoxide dismutase (SOD), catalase (CAT), and peroxidase (POD)]to overcome ROS-induced oxidative damage[42,76,108].Trehalose application can reduce stress-induced ROS accumulation by activating antioxidant defense mechanisms [50,109].
For example, Tre application (20 mmol L–1) increased antioxidant enzyme activities [SOD, CAT, POD, and ascorbate peroxidase(APX)] and decreased malondialdehyde (MDA) levels in rapeseed(Brassica napus L.)seedlings under CS(4 and–4°C)[51].Cold stress(–3.5 ± 0.5 °C) increased MDA and H2O2levels in wheat florets,while Tre application (1, 10, and 50 mmol L–1) increased antioxidant enzyme activities [APX, glutathione reductase (GR), dehydroascorbate reductase(DHAR), monodehydroascorbate reductase(MDHAR), glutathione peroxidase (GPX), and glutathione-stransferase] and non-enzymatic antioxidants (ascorbic acid and GSH)to minimize the accumulation of these metabolites[96].Similarly, Tre application (5%, 10%, and 15%) increased POD and CAT activities in pepper plants subjected to CS (4 °C) and reduced MDA levels and electrolyte leakage [102].Therefore, Tremediated tolerance to low temperature could be attributed to the modulation of the redox state by ROS scavenging and regulation of the ascorbate–glutathione cycle [110].
Cold stress affects membrane integrity by decreasing membrane fluidity and causing protein denaturation and aggregation.However, Tre actively maintains cell membrane integrity by promoting the formation of phosphate groups and hydrogen bonds that hinder protein aggregation and degradation [111].A notable case of this defensive mechanism was observed under salinity stress,in which Tre maintained the structural integrity of the lipid bilayer and enzyme functioning by conserving ion pumps that remove sodium ions from chloroplasts[64].More in-depth studies,mainly under CS, are needed to gain further insights into this defensive mechanism.
3.1.4.Trehalose upregulates Tre- and stress-associated genes under cold stress
The expression of Tre-synthesis genes, such as those of the TPP gene family, increased under CS, following exposure to salt stress,and in response to exogenous abscisic acid(ABA)application in different organs and at different growth stages in plants such as rapeseed [112] and wheat [113].Exposure to a combination of CS,drought, and exogenous ABA also induced OsTPP1 and OsTPP2 expression in the roots and shoots of rice seedlings [55].Furthermore, salt stress and ABA treatments rapidly increased OsTPP1 expression, while CS increased the expression of this gene more slowly [114].In addition, the overexpression of TPS and TPP increased Tre accumulation and improved CS tolerance in transgenic tobacco, Arabidopsis, and rice [56,58,115,116].
In tomato subjected to CS, Tre application (10 mmol L–1)increased Cu/Zn SOD, CAT1, and RESPIRATORY BURST OXIDASE 1(RBOH1) expression due to increased activities of these enzymes but decreased APX5 and GR1 expression due to decreased activities of these enzymes [57].Notably, APX and GR play key roles in the ascorbate–glutathione cycle,which is involved in alleviating oxidative stress[110].The controlled decrease in their expression under increased stress conditions could redirect other defense mechanisms, ultimately improving the adaptive responses of plants to CS.Similarly, the expression of antioxidant genes (CAT12, POD34,and FSD7), CS-responsive marker genes [CBF1, CBF2, CBF4 (Crepeat binding factor), COR6.6, COR15, COR25 (cold-responsive),COL1 (CONSTANS LIKE 1), and KIN1], and Tre-biosynthetic genes(TPS4,TPS8,and TPS9)increased in Tre-treated(20 mmol L–1)rapeseed seedlings under CS (4 and –4 °C) [51].Priming wheat seeds with Tre (0.5, 1, and 2 mmol L–1) increased the expression of cold-responsive (CBFs and CORs) and Tre-synthesis (TPP1 and TPP2) genes under CS (15 °C) [93].
3.2.Heat stress
Heat stress triggers ROS accumulation in the nucleus and other cellular compartments [117] and alters cell metabolism, physiology, and gene expression [5,10,13,118,119].Prolonged HS exposure disrupts membrane integrity and cell functions [120].Trehalose contributes to the physiological, biochemical, and molecular mechanisms that help plants overcome the adverse effects of HS (Table 1; Fig.3).
3.2.1.Trehalose modulates plant growth and development under heat stress
Heat stress adversely affects plant growth and development by inhibiting seed germination, root and shoot lengths, plant biomass,fruit set,and pollen viability[121].The oxidative conditions induced by HS inhibit seed germination.The buildup of Tre in this situation becomes a significant source of energy for germinating seeds to use for their recovery.The Tre precursor(T6P),crucial for controlling the glycolytic process of carbohydrate metabolism, also stimulates germination [53].The effect of exogenous Tre application on plant growth can vary depending on the plant species and stress response mechanisms.For instance,exogenous Tre application(1.5 mmol L–1)inhibited wheat seedling growth by decreasing root length, vertical height, leaf area, and leaf length under HS (40 °C) [122].Plants shorten their vegetative stage when confronting abiotic stresses to sustain and redistribute energy,improving their chances of survival[123],implying that Tre-treated wheat seedlings have more energy to participate in stress tolerance due to delayed development.Overall, it can be concluded that the influences of Tre application on growth are inconsistent and can include both improvement and inhibition, depending on the plant species and the complexities of their stress response mechanisms.Some species may display increased growth under similar stress conditions, highlighting the complexity of the impact of Tre application on plant physiology and stress tolerance/adaptation.
On the contrary, exogenous Tre application (0.5 mmol L–1)reduced the degree of leaf wilting and yellowing caused by HS(42°C)in wheat seedlings,improving their tolerance to HS by protecting the membrane system and decreasing cell membrane damage [90].Similarly, Pereira et al.[124] reported that Tre improved HS tolerance by sustaining biomembrane and protein function.The three major mechanisms for Tre-mediated stabilization of biological structure and function under HS are (1) stabilizing the native conformation of biomolecules by Tre as it forms H-bonds with the polar heads of lipids and proteins to function as water replacement in dehydrated conditions, (2) maintaining the structure and stability of biomolecules by entrapping residual water molecules around their surface,and(3)generating fewer reactive and protective structures that are stable under HS [125,126].
A Tre treatment enabled wheat plants to resist HS by improving root phenotypic properties with more robust vitality [87].In another study, Liu et al.[127] found that Tre addition (5, 10, 15,20, and 30 g L–1) improved the mycelial growth of Pleurotus pulmonarius under HS (40 °C).Trehalose-mediated plant growth and biomass increases could be due to improved relative water content and photosynthetic performance and stimulated antioxidant systems resulting from decreased MDA and H2O2accumulation.Trehalose application (10 μmol L–1) increased chickpea seedling length and survival rate under HS (45/40 °C), indicating that Tre also works downstream of ABA, partly influencing the protective effects of ABA[128].Hence,Tre functions as a central coordination factor in carbohydrate synthesis and recycling by interacting with the ABA signaling pathway [128].
3.2.2.Trehalose improves photosynthesis and photosynthetic apparatus under heat stress
Heat stress adversely affects respiration and photosynthesis by perturbing the mitochondrial and chloroplast electron transport chains[119].Disrupted thylakoid function and decreased activities of stromal enzymes such as RuBisCo activase that cause heatinduced inactivation of the Calvin–Benson–Bassham cycle decrease carbohydrate production [129–131].Exogenous Tre application (1.5 g L–1) decreased HS (43/37 °C)-induced losses in chlorophyll and improved photosynthetic gas exchange characteristics, including photosynthetic rate per unit leaf area (Pn), stomatal conductance (gs), and instantaneous carboxylation efficiency,in Emmenopterys henryi Oliv [132].In addition, Tre-treated plants had higher PSII efficiencies, chlorophyll fluorescence parameters(Fv/Fmratios), photochemical quenching, and electron transport than non-treated plants[132].Similarly,Tre application(30 mmol L–1) increased Pn, intercellular CO2concentration (Ci), and Fv/Fmratios in Paeonia lactiflora Pall.under HS(40°C).These results suggest that Tre protects photosynthesis against HS-induced inhibition.In addition, Tre-treated Paeonia lactiflora leaves showed better protection of chloroplast structures and higher antioxidant activities and photosynthetic capacity under HS than untreated leaves [133].
Trehalose can also form films on leaves, influencing stomatal behavior and reducing unnecessary water loss through transpiration, ultimately improving photosynthetic efficiency [77,134,135].Trehalose application (1.5 mmol L–1) protected the proteins in wheat thylakoid membranes from degradation, leading to an improved chloroplast ultrastructure [136].Trehalose application can prevent HS-induced destabilization of thylakoid membrane lipids and the accompanying loss of photosynthetic pigments,decreasing the activities of chlorophyll-degrading enzymes[137,138].Exogenous Tre application (0.5, 1.5, and 10 mmol L–1)improved gas exchange parameters, Fv/Fmratios, and WUE in wheat and maize seedlings exposed to 42 °C [91].
3.2.3.Trehalose improves antioxidant defense systems and prevents ROS accumulation under heat stress
Trehalose boosts antioxidant defenses in plants exposed to environmental stress [49,50].For example, Tre application (1 and 2 mmol L–1)increased CAT,APX,and POX activities in wheat seedlings and decreased heat (35 °C)-induced increases in H2O2and MDA [139].Similarly, maize seedlings treated with Tre (5, 10, 15,20, and 25 mmol L–1) had lower MDA contents with reduced electrolyte leakage under HS (47 °C) than untreated seedlings [140].Therefore, Tre is an important activator of antioxidant systems in plants exposed to stress.
Winter wheat plants treated with Tre had improved HS tolerance and lower levels of HS-induced electrolyte leakage, MDA and ROS accumulation, and lipoxygenase (LOX) activity than untreated plants [136].Trehalose application (5, 10, and 15 g L–1)decreased HS (28 °C)-induced growth inhibition and MDA levels in oyster mushroom (Pleurotus ostreatus) [141].Trehalose treatments (10, 30, and 50 mmol L–1) increased SOD activity in wheat plants under HS (22, 45, 55, and 65 °C) [142].Wheat seedlings pre-treated with Tre (1.5 mmol L–1) had enhanced CAT and APX activities and non-enzymatic antioxidant(ascorbate and GSH)contents under HS (40 ± 0.5 °C), reducing ROS production [143].Trehalose could increase H2O2scavenging by shielding CAT from heat inactivation.In conclusion,Tre-mediated reductions in oxidative stress might be due to antioxidant mechanism regulation,ROS scavenging, and membrane stabilization to eliminate ROS.
3.2.4.Trehalose upregulates Tre- and stress-associated genes under heat stress
Trehalose protects cellular membranes against HS by stimulating the PKC1 pathway, a dynamic intracellular signaling cascade[59] associated with stress responses and cell survival.In this setting, stimulation of the PKC1 pathway results from Tre accumulation rather than HS itself[59].Trehalose application(1.5 mmol L–1)enhanced the expression of SOD,CAT,POD,and APX genes in wheat seedlings exposed to HS[143].Trehalose-treated Arabidopsis plants had increased expression of vacuolar ATP synthase catalytic subunit A, a stress-related gene [144].Similarly, Luo et al.[87]reported that Tre application (0.5 mmol L–1) significantly altered the transcriptome signature of wheat roots under HS(42°C).Gene expression changes in Tre-treated wheat varieties, leading to HS tolerance,positively correlated with secondary metabolite synthesis and carbohydrate metabolism[90].These findings suggest that Tre regulates gene expression in response to abiotic stress.
4.Temperature stress management by engineering Tre-encoding genes
Numerous studies have focused on creating transgenic plants that overexpress genes involved in Tre metabolism [49,50] to improve resilience to temperature stress (see reviews by[49,50,142].For example, the expression of chimeric yeast TPS and TPP in A.thaliana under the control of the 35S promoter or stress-regulated rd29A promoter improved heat, salt, drought,and freezing tolerance [145].Similarly, transgenic alfalfa plants expressing yeast TPS1 and TPP2 genes were more resilient to HS than the wild-type [146].Transgenic tomato plants expressing an E.coli gene that encodes trehalose-6-phosphate synthase/phosphatase (TPSP) had increased amounts of Tre in seeds [84].These transgenic lines also had higher post-HS germination rates(>50%)than the wild-type(17%).Furthermore,the TPSP transgenic seeds expressed numerous HS genes more rapidly following exposure to high temperatures than wild-type seeds.Therefore, the accumulation of Tre and related metabolites promotes the expression of HS-responsive genes [84].Functional analysis revealed an underlying link between TPS1 and HS-dependent raffinose synthesis in Arabidopsis [147].Whereas the tps1-1 null mutant (TILLING line) with functionally manifest TPS1 isoforms differentially influenced carbohydrate metabolism under severe HS.Notably,increased TPS1 activity was associated with reduced endogenous Suc levels and improved HS tolerance in Arapidopsis; interference of T6P signaling caused a superior increase of transitory starch and Suc and was linked with improved HS tolerance [147].
Arabidopsis plants overexpressing Hevea brasiliensis HbTPS1 also enhanced heat and freezing tolerance [148].
Administering Tre to growing oyster mushrooms enhanced HS responses by promoting mycelial growth.Overexpression of the TPS gene in P.ostreatus mycelia increased the amount of Tre following exposure to HS[141].Exogenous Tre application and the overexpression of tps1 alleviated HS in P.ostreatus by increasing PPP and G6PDH activities that prevent lactate accumulation [88].Several reports have documented how the expression of Tre genes can improve tolerance to stresses other than temperature extremes.For example, overexpression of TPP increases high light tolerance in Parachlorella kessleri by increasing Tre contents [149].OsTPP3-overexpressing rice plants had a higher tolerance to drought than wild-type plants [150].Similarly, ClTPS3 overexpression in A.thaliana significantly improved salt tolerance [151].In rice, phospho-OsbHLH002 instantly triggered OsTPP1 expression (responsible for Tre biosynthesis), increasing Tre and chilling tolerance [152].
Gene editing is a highly flexible technology that can increase genetic variation precisely [153–157].Arabidopsis plants containing the Tre substrate-binding domain (25 amino acid conserved signature ‘PGXRFXEXYXWDSXW,’ with notable conservation between diverse plant taxa) had a higher tolerance to drought stress than the wild-type [158].Conversely, knocking out Tre(NtTRE)in tobacco using CRISPR/Cas9 technology increased HS sensitivity compared to wild-type plants [159].
5.Trehalose crosstalk and interaction with other molecules under stress

Fig.4.Overview of trehalose-involving crosstalk and interactions with other molecules helping plant acclimatization and tolerance against temperature stress.The genes COR,KIN1,CBFs,HsFA2,HSBP2,and OsNAC5,and transcription factors CBF/DREB1,AP2EREBP,HSFs,and NAC regulate in response to temperature stress signals and/or trehalose.ABA,abscisic acid;APX,ascorbate peroxidase;Ca2+,calcium;CAT,catalase;CK,cytokinin;ET,ethylene;GA,gibberellic acid;IAA,indole-3-acetic acid;NO,nitric oxide;POD,peroxidase; ROS, reactive oxygen species; SA, salicylic acid; SOD, superoxide dismutase.
Trehalose interacts with numerous pathways involving other signaling molecules, such as sugars (e.g., Suc, glucose, fructose),amino acids (e.g., proline, hydroxy-proline, and glycine betaine),phytohormones (e.g., ABA, indole acetic acid, ethylene, cytokinins,gibberellic acid),and polyamines(e.g.,putrescine,spermidine,and spermine),to boost secondary metabolism and carbohydrate metabolism and thus enhance tolerance to environmental stresses[50,160,161], as illustrated in Fig.4.The T6P/SnRK1 module might act as a sensor,controlling the metabolic reprogramming required to optimize fitness under different environmental conditions[161,162].In contrast,the signaling functions of T6P mainly inhibit SnRK1 [162].However, T6P, glucose 1-phosphate (G1P), and G6P have similar signaling responses, which control SnRK1 functions.Sucrose and/or Tre mediate the regulation of hexokinase (HXK)and target rapamycin (TOR) functions via the HXK-autonomous Suc-specific and T6P/SnRK1 pathways, respectively [52].
Complex signaling systems trigger plant responses to temperature stress (Fig.4), including changes in membrane fluidity that activate temperature signaling pathways [163], such as calcium(Ca2+) channels (Fig.4) that trigger numerous downstream processes, including the regulation of gene expression [164].Regulating nitric oxide(NO)and ROS generation is particularly important,as these signaling molecules,together with Ca2+,activate transcription factors that promote growth [163].
Transgenic Arabidopsis plants expressing TPS (otsA) or TPP (otsB)had increased T6P levels when supplied with an exogenous Suc source[165].T6P regulates starch metabolism[166],modulating leaf Suc and starch accumulation during the day and starch degradation at night[167].TPS1 modulates long-term HS tolerance due to its controlling function in carbon partitioning and Suc homeostasis[147].Exogenous Tre supplementation induced starch accumulation in Arabidopsis by affecting the starch-synthesizing enzyme ADP-glucose pyrophosphorylase(AGPase).T6P promoted AGPase activation through thioredoxin(Trx)-mediated processes [168].Trehalose supplementation significantly increased soluble protein, sugar, and proline contents and enhanced the expression of Tre-synthesis genes (TPS4, TPS8, and TPS9) in rapeseed seedlings, improving cold tolerance [51].
6.Genome-wide characterization of Tre-encoding genes for crop improvement
Two classes of TPS genes are present in plant genomes.Comparative genome analysis identified 7 genes with 16 introns in class I TPS genes and 27 genes with two introns in the protein-coding region of class II TPS genes in the Arabidopsis,rice,and Populus genomes [169].An analysis of protein–protein interactions in 26 TPS genes from rapeseed revealed roles in galactose metabolism, glycolysis/gluconeogenesis, carbohydrate transmembrane transport,fructose and mannose metabolism, pentose phosphate pathway,and trehalose-phosphatase activity[112].In Arabidopsis,TPS genes regulate growth and carbohydrate utilization [170], hypocotyl growth [171], and flowering time [172].Maize TPS expression increased in response to cold and salt stresses [173].
The number of Tre-encoding genes varies with genome size in different plant species (Table 2).A genome-wide in-silico analysis revealed that differential regulation of specific TPS and TPP gene classes occurs in response to abiotic stresses such as extreme temperatures, drought, and salinity (Table 2).
Several genome-wide approaches revealed putative Treencoding gene functions (Table 2; Fig.5).For example, the evolutionary relationships and duplication events of 31 wheat TaTPP genes were involved in leaf senescence,development,and stress tolerance [65].Promoter analysis revealed cis-regulatory elements in the TaTPP genes linked to hormone functions, stress, and plant growth and development [65].Transcriptional analysis of TaTPP genes in wheat revealed differential expression patterns during different developmental phases and in various organs [65].Moreover,qRT-PCR-based expression analysis revealed that numerous TaTPP genes were induced under salinity and drought stresses [65], providing basic foundations for wheat improvement by engineering TaTPP genes.Comparative genome-wide analysis of the wheat TPP gene family identified 31 TPP genes divided into 6 sub-families[113].Furthermore, RNA-seq analysis identified 26 TaTPP differentially expressed genes with roles in ABA metabolism and signaling related to cold and drought stresses.Overexpression of TaTPP11 in Arabidopsis delayed growth and enhanced drought tolerance [113].
7.Overview of mechanisms underlying Tre-mediated temperature stress responses and tolerance in plants
The mechanisms underlying Tre-mediated temperature stress responses and tolerance in plants are multifaceted and involve various physiological and molecular processes.Below is an overviewof these mechanisms, with detailed case-by-case examples discussed above and in Table 1.

Table 2 Summary of up-to-date genome-wide studies to unravel the crucial role of trehalose genes in the genomes of different plant species.
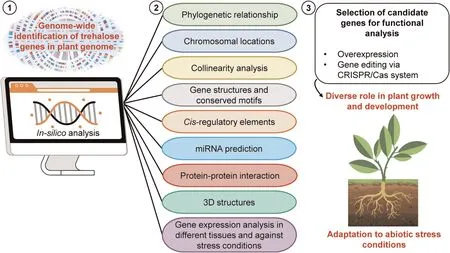
Fig.5.Overview of in-silico tools used for genome-wide identification and characterization of trehalose genes in diverse plant genomes in three main steps: (1) gene identification in a particular plant genome,(2)detailed characterization of identified genes,and(3)selection of novel candidate genes for functional characterization to reveal their mechanistic role in crop improvement under stress.
Osmoprotection and metabolic regulation: Tre is an osmoprotectant that helps maintain cellular osmotic balance under temperature stress.It prevents cellular dehydration, stabilizes macromolecules,and safeguards cellular structures.Trehalose regulates metabolic pathways, particularly those related to energy metabolism and stress-responsive genes, by providing an additional energy source for cells to cope with stress conditions.
Stomatal regulation and WUE: Tre affects stomatal activity,which is crucial for adjusting water loss during temperature stress.It regulates stomatal conductance,affecting transpiration rates and improving WUE.
Defense against oxidative stress:Tre plays a vital role in inducing antioxidant properties within plants, essential for combating oxidative stress caused by extreme temperatures.It helps scavenge ROS and interacts with ROS-scavenging antioxidant enzymes,reducing oxidative damage to cellular components.
Gene expression and signaling networks: Tre exerts control over gene expression and signaling networks, which are critical for stress responses and adaptation.It can trigger/repress the expression of stress-responsive genes.It can also interact with other signaling pathways and regulatory molecules involved in Tre-mediated stress responses and tolerance mechanisms.
8.Conclusion and future directions
Trehalose is a versatile and multifunctional molecule in plants,serving as an osmoprotectant, storage reserve, and stress protectant.It plays a crucial role in maintaining cellular osmotic equilibrium, stabilizing macromolecules, and safeguarding cellular structures, particularly under temperature stress.It also regulates metabolic pathways,including those associated with energy metabolism and stress-responsive genes.Trehalose can influence stomatal regulation,enhancing WUE by adjusting transpiration rates.Its role in defense against oxidative stress is vital,as it helps scavenge ROS and interacts with antioxidant enzymes.Furthermore, Tre or T6P acts as a key player in gene expression and signaling networks,influencing the expression of stress-responsive genes and crosstalk with other signaling pathways and regulatory molecules.While it is clear that Tre interacts with various phytohormones and other signaling molecules, the precise mechanisms that facilitate the coordinated control of fitness and productivity in response to changing environmental conditions remain poorly understood.Most research on Tre has focused on Arabidopsis and a few crop systems, mainly related to temperature stress.Further studies on a broader range of species are needed to understand the importance of Tre in plant biology and agriculture.
The T6P/SnRK1/TOR signaling system is essential in regulating environmental stress responses,functioning as a molecular switch for environmental stress signaling.Further research is needed to decipher this system’s intricate crosstalk and regulatory mechanisms.With T6P, Tre controls plant growth, development, and defense, promoting biosynthetic pathways linked to growth and yield.For example, enhanced T6P levels promote yield enhancement and stability, while reduced levels enhance the deployment of carbon assets and carbon mobilization linked to stress responses[196].Recent years have witnessed significant yield improvements in cereal crops such as maize,rice,and wheat by directly modifying T6P levels through overexpression of the OsNAC23 NAC transcription factor, simultaneously increasing T6P accumulation and repressing Tre levels [197,198].Furthermore, a close relationship exists between T6P and Suc levels.The T6P:Suc ratio critically controls overall Suc levels to direct plant growth and development.Hence, further investigations are required to establish the precise nature of this regulatory mechanism in different tissues and organs, mainly in response to temperature stress.
The literature supports the conclusion that engineering Treencoding genes can improve plant growth and productivity,particularly under stress.Therefore, efforts are needed to produce Tregene-based cultivars for use in agriculture.Studies are also needed to identify positive and negative regulators of Tre metabolism that directly or indirectly modify its accumulation in plants to enhance our understanding of how Tre modifies stress tolerance and growth.
Trehalose is a versatile biomolecule that can be exogenously applied to crop plants to improve growth, yield, and stress tolerance.The chemistry between exogenous and endogenous Tre and its role in augmenting stress tolerance is a captivating area of study, and differences in exogenous and endogenous Tre levels(see main text and Table 1) are influenced by species-specific responses and the energetic nature of Tre metabolism under stress conditions.There is enormous potential for Tre application in nanotechnology.For example, Tre can be used as a functional group with magnetic nanoparticles to improve protein stability in biological systems.Poly(Tre)-nanoparticles have already been used to inhibit amyloid/polyglutamine aggregation in treating disease.Advances in Tre application methods could further improve crop growth and productivity by optimizing Tre application rates and management for different crops under various stresses in the field.
Trehalose is an attractive signaling molecule strongly influencing gene expression,particularly genes encoding proteins involved in plant antioxidant defense.However, the precise mechanisms facilitating this control are largely undocumented, presumably due to the assumption that T6P/SnRK1/TOR interactions exert this regulation.More studies are needed to determine Tre’s precise molecular and metabolic effects in different cell types under diverse environmental conditions.In addition, few studies have investigated the signal transduction systems that regulate Tre metabolism in response to different stresses or the transcriptional and post-translational modifications that ensure appropriate responses to different stresses.An improved understanding of the full scope of Tre-associated functions in plants will undoubtedly contribute to the success of future crop improvement programs.
CRediT authorship contribution statement
Ali Raza:Conceptualization, Visualization, Writing – original draft, Writing – review & editing.Savita Bhardwaj:Writing –review&editing.Md Atikur Rahman:Writing–review&editing.Pedro García-Caparrós:Writing – review & editing.Madiha Habib:Writing–review&editing.Faisal Saeed:Writing–review& editing.Sidra Charagh:Writing – review & editing.Christine H Foyer:Writing–review&editing.Kadambot H.M.Siddique:Conceptualization, Visualization, Writing – review & editing, Supervision.Rajeev K.Varshney:Conceptualization, Visualization,Writing – review & editing, Supervision, Funding acquisition.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgments
Rajeev K.Varshney is thankful to the Food Futures Institute of Murdoch University for financial support.This work was supported by the Food Futures Institute of Murdoch University to Rajeev K.Varshney.

- The Crop Journal的其它文章
- Corrigendum to ‘‘GmTOC1b negatively regulates resistance to Soybean mosaic virus”.[Crop J.11 (2023) 1762–1773]
- Decoding the inconsistency of six cropland maps in China
- Wetting alternating with partial drying during grain filling increases lysine biosynthesis in inferior rice grain
- OsDA1 positively regulates grain width in rice
- A polygalacturonase gene OsPG1 modulates water homeostasis in rice
- The ABA synthesis enzyme allele OsNCED2T promotes dryland adaptation in upland rice