
Prohibitin 1 inhibits cell proliferation and induces apoptosis via the p53-mediated mitochondrial pathway in vitro
2024-03-07 08:18:22JuanJuanShiYiKaiWangMuQiWangJiangDengNingGaoMeiLiYaPingLiXinZhangXiaoLiJiaXiongTaoLiuShuangSuoDangWenJunWang
Juan-Juan Shi,Yi-Kai Wang,Mu-Qi Wang,Jiang Deng,Ning Gao,Mei Li,Ya-Ping Li,Xin Zhang,Xiao-Li Jia,Xiong-Tao Liu,Shuang-Suo Dang,Wen-Jun Wang
Abstract BACKGROUND Prohibitin 1 (PHB1) has been identified as an antiproliferative protein that is highly conserved and ubiquitously expressed,and it participates in a variety of essential cellular functions,including apoptosis,cell cycle regulation,proliferation,and survival.Emerging evidence indicates that PHB1 may play an important role in the progression of hepatocellular carcinoma (HCC).However,the role of PHB1 in HCC is controversial.AIM To investigate the effects of PHB1 on the proliferation and apoptosis of human HCC cells and the relevant mechanisms in vitro.METHODS HCC patients and healthy individuals were enrolled in this study according to the inclusion and exclusion criteria;then,PHB1 levels in the sera and liver tissues of these participates were determined using ELISA,RT-PCR,and immunohistochemistry.Human HepG2 and SMMC-7721 cells were transfected with the pEGFPPHB1 plasmid and PHB1-specific shRNA (shRNA-PHB1) for 24-72 h.Cell proliferation was analysed with an MTT assay.Cell cycle progression and apoptosis were analysed using flow cytometry (FACS).The mRNA and protein expression levels of the cell cycle-related molecules p21,Cyclin A2,Cyclin E1,and CDK2 and the cell apoptosis-related molecules cytochrome C (Cyt C),p53,Bcl-2,Bax,caspase 3,and caspase 9 were measured by real-time PCR and Western blot,respectively.RESULTS Decreased levels of PHB1 were found in the sera and liver tissues of HCC patients compared to those of healthy individuals,and decreased PHB1 was positively correlated with low differentiation,TNM stage III-IV,and alphafetoprotein ≥ 400 μg/L.Overexpression of PHB1 significantly inhibited human HCC cell proliferation in a timedependent manner.FACS revealed that the overexpression of PHB1 arrested HCC cells in the G0/G1 phase of the cell cycle and induced apoptosis.The proportion of cells in the G0/G1 phase was significantly increased and the proportion of cells in the S phase was decreased in HepG2 cells that were transfected with pEGFP-PHB1 compared with untreated control and empty vector-transfected cells.The percentage of apoptotic HepG2 cells that were transfected with pEGFP-PHB1 was 15.41% ± 1.06%,which was significantly greater than that of apoptotic control cells (3.65% ± 0.85%,P < 0.01) and empty vector-transfected cells (4.21% ± 0.52%,P < 0.01).Similar results were obtained with SMMC-7721 cells.Furthermore,the mRNA and protein expression levels of p53,p21,Bax,caspase 3,and caspase 9 were increased while the mRNA and protein expression levels of Cyclin A2,Cy-clin E1,CDK2,and Bcl-2 were decreased when PHB1 was overexpressed in human HCC cells.However,when PHB1 was upregulated in human HCC cells,Cyt C expression levels were increased in the cytosol and decreased in the mitochondria,which indicated that Cyt C had been released into the cytosol.Conversely,these effects were reversed when PHB1 was knocked down.CONCLUSION PHB1 inhibits human HCC cell viability by arresting the cell cycle and inducing cell apoptosis via activation of the p53-mediated mitochondrial pathway.
Key Words: Prohibitin 1;Hepatocellular carcinoma cells;Apoptosis;Cell cycle;Mitochondrial pathway
lNTRODUCTlON
Prohibitin (PHB) proteins are antiproliferative proteins that are highly conserved and ubiquitously expressed in various eukaryotic cell types[1].The PHB proteins include PHB1 and PHB2;these proteins are encoded by genes that are located on different chromosomes and are associated with a large eukaryotic mitochondrial complex (PHB complex),which contains ring-like structures with chaperone-like functions or scaffolding activities and stabilizes mitochondrial proteins[2].PHB1 is a 30-kDa membrane protein that is predominantly located in the mitochondrium,nucleus,and plasma membrane and performs multiple cellular functions,such as functions related to apoptosis,cell cycle regulation,proliferation,metabolism,senescence,and differentiation[1,3-5].The biological activity of PHB1 appears to be complex,and this protein participates in various cellular pathways by specifically interacting with many proteins that are located in different subcellular environments[2,3,6].In the nucleus,PHB1 has been reported to interact with p53,E2F,retinoblastoma protein (Rb),and c-Myc,among other proteins,which are closely related to the inhibition of the cell cycle and the induction of apoptosis[6-9].
The tumour suppressor protein p53 causes a variety of responses,such as cell cycle inhibition and apoptosis induction,when it is activated by numerous stress signals,including DNA damage,hypoxia,and heat shock[10].P53 mediates cell apoptosis by activating mitochondrial apoptotic pathways that promote the release of cytochrome C (Cyt C) into the cytoplasm;the interaction of Cyt C with apoptosis protease-activating factor (Apaf-1) in the cytoplasm activates caspase signalling pathways that involve,for example,caspase 9,caspase 3,and caspase 7,and ultimately induces cell apoptosis[11].Cyt C release and Bcl-2 family member activation are particularly essential steps in the mitochondrial pathway.The Bcl-2 family is predominantly composed of antiapoptotic proteins,such as Bcl-2 and Bcl-XL,and proapoptotic proteins,including Bax,Bad,and Bek[11,12].The mitochondrial pathway is predominantly regulated by the p53 effectors Bcl-2 and Bax;p53 upregulates the expression of Bax and downregulates the expression of Bcl-2,resulting in the induction of apoptosis[12,13].Previous studies have demonstrated that PHB1 is overexpressed in gastric cancer cells and gynaecologic cancer cells,and the overexpression of PHB1 has been shown to activate the mitochondrial pathway;this subsequently upregulates Bax,Bak,caspase 9,and caspase 3 and downregulates Bcl-2,resulting in cancer cell apoptosis[14,15].Moreover,PHB1 downregulation delays abrin (ABR)-triggered cell apoptosisviatranslocation of the PHB1-p53 complex from the cytosol to the nucleus and activation of p53-mediated Bax transcription in human T leukaemia cancer cells[16].These findings suggest that PHB1 may induce apoptosisviathe p53-mediated mitochondrial pathway.
Although several functions of PHB1 have been described,little is known about the definitive functions and mechanisms of this protein in human hepatocellular carcinoma (HCC) cells.In the present study,we investigated the effect of PHB1-induced apoptosis and the underlying molecular mechanisms in HCC cells.Our findings demonstrated that PHB1 is a unique tumour suppressor and that its overexpression induces HCC cell apoptosisviathe p53-mediated mitochondrial pathway.
MATERlALS AND METHODS
Patients
The participants included well-documented HCC patients who were randomly enrolled from The Second Affiliated Hospital of Xi’an Jiaotong Uni-versity from November 2019 to October 2021.Healthy individuals who served as controls were randomly enrolled from the health examination centre of The Second Affiliated Hospital of Xi’an Jiaotong University.A total of 84 serum samples,including samples from 42 HCC patients and 42 healthy individuals,were included in this study.This study included liver tissue samples from 30 HCC patients and 30 healthy individuals.The healthy individuals had no abnormal aminotransferase activity,no history of liver disease,and no history of other tumour diseases,and they were negative for hepatitis B virus,hepatitis C virus,and human immunodeficiency virus infection.This study was performed in accordance with the principles of the Declaration of Helsinki,approved by the Institutional Review Board of The Second Affiliated Hospital of Xi’an Jiaotong University,and performed in accordance with university guidelines for the protection of participant information.All the participants provided written informed consent prior to blood sampling and liver surgery.
Cell lines and cell culture
The human HCC cell lines HepG2,SMMC-7721,and Huh 7 and the human normal liver cell line L-02 were obtained from the American Type Culture Collection (Manassas,VA,United States).The cells were grown in Dulbecco’s modified Eagle’s medium (DMEM;HyClone,Logan,UT,United States) supplemented with 10% (v/v) foetal bovine serum (HyClone),100 U/mL penicillin (Sigma-Aldrich,St.Louis,United States),and 100 μg/mL streptomycin sulfate (Sigma-Aldrich) at 37 °C in a 5% (v/v) CO2humidified atmosphere.The cell culture medium was replaced every other day.When the cells reached 80% confluence,the cells were washed with phosphate-buffered saline (PBS;137 mmol/L NaCl,2.7 mmol/L KCl,4.3 mmol/L Na2HPO4,1.4 mmol/L KH2PO4,pH 7.4) and then digested using 0.25% (w/v) tryptase (HyClone).The cells were harvested using DMEM followed by centrifugation at 800 rpm for 5 min.Cells in the logarithmic growth phase were resuspended in DMEM,plated at appropriate densities in appropriate plates,and subjected to passage as usual for further experimentation.
Construction of pEGFP-PHB1 recombination vector
The coding region ofPHB1was amplified by PCR,digested with Hind III and Kpn I (TAKARA,Japan),and directly cloned and inserted into the pEGFP-N1 plasmid (TAKARA,Japan) to generate the pEGFP-PHB1 plasmid.Briefly,the coding region of PHB1 (819 bp) was amplified using high-fidelity PCR (Thermo Fisher Scientific,United States).The coding regions of PHB1 and pEGFP-N1 were digested with the restriction enzymes Hind III and Kpn I.The digestion products were examined by electrophoresis on a 1% (w/v) agarose gel (Sigma-Aldrich,United States).Ligation reactions (Ligation Kit,TAKARA,Japan) were performed to join the DNA fragments,which were subsequently transformed into competentEscherichia colicells (E. coliTop 10,TAKARA,Japan) at 37 °C overnight (12-16 h).Colony selection was performed by PCR,and the amplicons were examined by electrophoresis using a 1% (w/v) agarose gel.Plasmid extraction from positive colonies was performed with an E.Z.N.A.®Endo-Free Plasmid Midi Kit (Omega,GA,United States),and the sequences were verified by sequencing.The primer pair that was used to amplify PHB1 was 5’-TGGGAGGTCTATATAAGCAGAG-3’ and 5’-CGTCGCCGTCCAGCTCG ACCAG-3’.
RNA interference
The PHB1-specific shRNA plasmid (shRNA-PHB1),its positive control plasmid (shRNA-GAPDH),and its negative control plasmid (shRNA-control) were purchased from Biomics Biotechnologies Co.,Ltd.(Jiangsu,China).The sequence of shRNA-PHB1 is 5’-GTAGCAAAGATTTACAGAA-3’,the sequence of shRNA-GAPDH is 5’-GTATGCAACAGCCTCAAG-3’,and the sequence of shRNA-control is 5’-GTGTTTGAGTCCATTGGCA-3’.
Transfection assay
HepG2 and SMMC-7721 cells in the logarithmic growth phase were seeded in 6-well plates at a density of 2 × 105cells/well in 2 mL of growth medium without antibiotics and incubated in 5% CO2at 37 °C for 24 h.The cells were then transfected with InvitrogenTMDMRIE-C Reagent (Thermo,CA,United States).Briefly,2 μg of DNA was diluted in 500 μL of serum-free Opti-MEM®I,and 8 μL of DMRIE-C was diluted in Opti-MEM®I.The resulting solutions were incubated at room temperature for 30-45 min.The diluted DNA was then combined with the diluted DMRIE-C,mixed gently,and incubated for 15-45 min at room temperature.The complexes were added to the cells,mixed gently by rocking the plate back and forth,and incubated at 37 °C in a CO2incubator for 4 h.The medium was replaced with 2 mL of growth medium supplemented with serum,and gene expression was measured after 24 h.
ELISA
The levels of PHB1 in patient serum were determined using an ELISA kit (CUSABIO,Wuhan,China) according to the manufacturer’s protocols.Serum samples were added to a 96-well plate coated with the capture antibody,and the plate was incubated with 100 μL of horseradish peroxidase (HRP)-conjugated secondary antibody for 60 min at 37 °C.The cells were then washed five times with 400 μL of wash solution.Fifty microlitres of chromogen solution A and 50 μL of chromogen solution B were added to each well,and the plate was incubated for 15 min at 37 °C.Finally,50 μL of Stop Solution was added to each well,and the optical density (OD) was measured within 15 min using a spectrophotometer (ND-1000,Thermo Fisher,United States) at double wavelengths of 450 nm and 630 nm.The concentrations of PHB1 were calculated according to a standard curve.
Histology and immunohistochemical analysis
The normal tissues (NT) from healthy individuals (n=30) as well as peritumoral tissues (PT) and tumour tissues (TT) from HCC patients (n=30) were fixed with 4% (w/v) paraformaldehyde for 15 min,washed five times with PBS,permeabilized with 0.1% (v/v) Triton X-100 (Sigma-Aldrich) for 5 min,embedded in paraffin,and cut into 5-μm sections.After deparaffinization,the histological sections were incubated at 100 °C for 15 min in sodium citrate buffer (10 mmol/L sodium citrate,0.05% (v/v) Tween 20,pH 6.0) for antigen retrieval and treated with 3% (v/v) hydrogen peroxide (H2O2) at room temperature for 15 min to block endogenous peroxidase activity in the tissues.Next,the tissues were washed five times with PBS and blocked with 5% (w/v) bovine serum albumin (BSA;Sigma-Aldrich) and 1% (v/v) normal donkey serum (Sigma-Aldrich) for 1 h at room temperature.The tissues were incubated with a rabbit monoclonal anti-PHB1 primary antibody (1:300 dilution,ab75766;Abcam,MA,United States) for 16 h at 4 °C,followed by incubation with horseradish peroxidase-conjugated anti-rabbit IgG (1:1000 dilution;Santa Cruz,CA,United States) for 1 h.The cell nuclei were stained with 4',6-diamidino-2-phenylindole (DAPI;Sigma-Aldrich),and the tissues were mounted with neutral balsam mounting medium.The tissues were visualized using an inverted microscope (Nikon,Tokyo,Japan).
Cell viability assay
Cell viability was determined using a 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetraazolium bromide (MTT;Sigma-Aldrich) assay as previously described[17].HepG2 and SMMC-7721 cells were plated in 96-well plates at a density of 6 × 103cells/well and incubated for 24 h;then,they were transfected with pEGFP-PHB1 or shRNA-PHB1.After 24,48,or 72 h of transfection,5 mg/mL MTT solution was added to each well and incubated for 4 h.The culture medium was then replaced with 150 μL of dimethyl sulfoxide.After shaking the culture plates for 5 min,OD was measured using an ND-1000 spectrophotometer at a wavelength of 570 nm.The cell viability percentages were calculated as follows: (ODtreated/ODcontrol) × 100%.
Cell cycle analysis
HepG2 and SMMC-7721 cells were incubated for 48 h after transfection,washed with ice-cold PBS twice,and fixed in 70% (v/v) ice-cold ethanol overnight at -20 °C.The cells were resuspended in Tris-HCl buffer (10 mmol/L Tris-HCl,pH 7.5) containing 20 mg/L ribonuclease A (RNase A;Sigma-Aldrich) for 15 min,followed by incubation with 50 mg/L propidium iodide (PI;Sigma-Aldrich) for 1 h at 37 °C in the dark.The cell cycle distribution was then determined using a FACscan flow cytometer (CALIBUR,BD,United States),and the data were analysed with ModFit LT 2.0 software (Verity Software House,United States).
Apoptosis assay
Cell apoptosis was analysed using Annexin V-FITC and PI double staining (Sigma-Aldrich) as previously described[18].Briefly,HepG2 and SMMC-7721 cells were incubated for 48 h after transfection and then harvested and stained with 10 μL of Annexin V-FITC and 5 μL of PI in cold binding buffer (50 mmol/L HEPES,700 mmol/L NaCl,12.5 mmol/L CaCl2,pH 7.4) for 15 min at room temperature in the dark.After washing,cell apoptosis was analysed using a FACscan flow cytometer (CALIBUR,BD,United States).
Cyt C release assay
HepG2 and SMMC-7721 cells were incubated for 48 h after transfection and then harvested and resuspended in 400 μL of mitochondrial protein isolation buffer (AMERSCO,Ohio,United States).The cells were homogenized with a polytron homogenizer on ice and then centrifuged at 1000 ×gat 4 °C for 10 min to pellet the nuclei and unbroken cells.The collected supernatants were centrifuged at 14000 ×gat 4 °C for 15 min to pellet the cytosolic proteins.The pellets were resuspended in mitochondrial protein isolation buffer and then centrifuged at 14000 ×gfor 1 min at 4 °C to pellet the mitochondrial proteins.The mitochondrial and cytosolic proteins were subjected to Western blot analysis as described below.
Quantitative real-time PCR analysis
HepG2 and SMMC-7721 cells were incubated for 48 h after transfection and then harvested.Total RNA was isolated using an E.Z.N.A.®Total RNA Kit (Omega,Norcross,GA,United States) as previously described[19].Briefly,1 μg of each RNA sample was reverse transcribed to synthesize complementary DNA (cDNA) with an RT-PCR kit (TAKARA,Japan).Quantitative real-time PCR was performed using a SYBR®Premix Ex TaqTMII kit (TAKARA,Japan) to analyse the relative expression ofPHB1,p53,p21,Cyclin A2,Cyclin E1,CDK2,Cyt C,Bax,Bcl-2,Caspase 3,andCaspase 9according to the manufacturer’s protocols.The RT-PCR mixture contained 12.5 μL of 2 × SYBR Ex Taq II,2 μL of RNA reaction,1 μL of forward primer,1 μL of reverse primer,and 8.5 μL of sterile distilled water in a final volume of 25 μL.The reaction was performed at 95 °C for 30 s for one cycle,and then at 95 °C for 5 s,60 °C for 60 s,and 72 °C for 60 s for 40 cycles.The PCR products were separated on a 2% agarose gel and stained with ethidium bromide (Sigma-Aldrich,United States).GAPDHwas used as an internal control.RT-PCR products were detected using an iQ5 Multicolor Real-Time PCR Detection System (Bio-Rad,Hercules,CA,United States).
Western blot analysis
HepG2 and SMMC-7721 cells were incubated for 48 h after transfection and then harvested.The cells were washed with ice-cold PBS,and cell lysates were prepared by lysing the cells with ice-cold RIPA lysis buffer [20 mmol/L Tris,150 mmol/L NaCl,1% (v/v) Triton X-100,1% (w/v) digestive phosphatase inhibitors,1% (w/v) protease inhibitors,and 1% (w/v) phenylmethyl sulfonylfluoride,pH 7.5 (Sigma-Aldrich)].The cell lysates were centrifuged at 14000 ×gat 4 °C for 15 min,after which the total protein concentrations in the supernatants were determinedviathe BCA assay (Kangweishiji,Beijing,China) according to the manufacturer’s protocol.Thirty micrograms of protein samples per lane was separated on 12% (w/v) SDS-PAGE gels (Sigma-Aldrich) and electrically transferred to polyvinylidene difluoride membranes (Sigma-Aldrich).The membranes were blocked with 5% (w/v) BSA (Sigma-Aldrich) in Tris-buffered saline (TBS;0.1 M,pH 7.4) before incubation with primary antibodies overnight at 4 °C.The primary antibodies that were used included antibodies against PHB1 (1:5000 dilution,ab75766;Abcam),p53 (1:1000 dilution,ab31333;Abcam),p21 (1:1000 dilution,ab47452;Abcam),Cyclin A2 (1:1000 dilution,ab 181591;Abcam),Cyclin E1 (1:1000 dilution,ab33911;Abcam),CDK2 (1:1000 dilution,ab32147;Abcam),Caspase 3 (1:500 dilution,ab13847;Abcam),Caspase 9 (1:1000 dilution,ab63488;Abcam),Cyt C (1:5000 dilution,ab 133504;Abcam),Bax (1:1500 dilution,ab10813;Abcam),and Bcl-2 (1:1000 dilution,ab59348;Abcam).After washing,the membranes were incubated with HRP-conjugated goat anti-rabbit secondary antibodies (Santa Cruz) for 1 h at room temperature with agitation.The proteins were detected using the ECL Western Blotting Kit Reagent (Thermo Fisher,United States) and analysed using Syngene G:Box tools (Syngene,United Kingdom).β-Actin (Santa Cruz) was used as an internal control.
Statistical analysis
Representative data are presented as the mean ± SD and were tested for normality and equal variance.The statistical analyses were performed using Student'sttests or one-way ANOVA with Bonferronipost hoccorrection with SPSS 15.0 software (SPSS,Inc.,United States).All the experiments were repeated at least three times with similar results.Differences were considered significant atP< 0.05.
RESULTS
Low expression of PHB1 is associated with tumour progression in HCC patients
To investigate PHB1 expression in HCC patients,serum and liver tissue samples were harvested from HCC patients.The mean serum PHB1 levels were significantly lower in patients with HCC than in healthy individuals (154.86 ± 23.10 pg/mLvs399.48 ± 39.1 pg/mL,P< 0.001;Figure 1A).Next,the mRNA expression levels ofPHB1in the liver samples of the HCC patients (n=30) and healthy individuals (n=30) were examined using RT-PCR.ThePHB1mRNA expression in the PT and TT of HCC patients was significantly lower than that in the NT of healthy individuals (P< 0.01 orP< 0.001;Figure 1B).

Figure 1 Prohibitin 1 expression in hepatocellular carcinoma patients.A: Serum levels of Prohibitin 1 (PHB1) in hepatocellular carcinoma (HCC) patients (n=42) and healthy individuals (n=42) were measured by ELISA;B-D: PHB1 mRNA levels investigated using real-time PCR (B),rate of positive PHB1 expression (C),and immunohistochemical staining for PHB1 measured via immunohistochemistry (D) in normal tissues from healthy individuals (n=30) as well as in peritumoral tissues and tumour tissues from HCC patients (n=30,200 × and 400 ×).The data are presented as the mean ± SEM of three independent experiments with similar results.aP < 0.05,bP < 0.01,cP < 0.001 vs healthy individuals.PT: Peritumoral tissues;TT: Tumour tissues;NT: Normal tissues;HCC: Hepatocellular carcinoma;PHB1: Prohibitin 1.
To confirm the RT-PCR findings,we performed immunohistochemistry on TT and NT from the individuals mentioned above (n=30).PHB1 was located mainly in the cytoplasm and nucleus of HCC cells.The PT and TT of HCC patients exhibited lower levels of PHB1 staining than did the NT of healthy individuals (P< 0.05 orP< 0.001;Figure 1C and D),and lower levels of PHB1 were positively correlated with low differentiation,TNM stage III-IV,and alpha-fetoprotein ≥ 400 μg/L (P< 0.05;Table 1).

Table 1 Clinicopathological analyses of the Prohibitin 1 expression in tumor tissues of 30 primary hepatocellular carcinoma patients
PHB1 inhibits HCC cell proliferation
Because low expression levels of PHB1 were observed in the HCC patients,we next examined changes in PHB1 expression in HepG2,SMMC-7721,Huh 7,and L-02 cellsin vitro.PHB1 mRNA and protein levels were lower in HepG2 and SMMC-7721 cells than in Huh 7 and L-02 cells (Figure 2A).Therefore,HepG2 and SMMC-7721 cells were used for all the subsequent experiments.
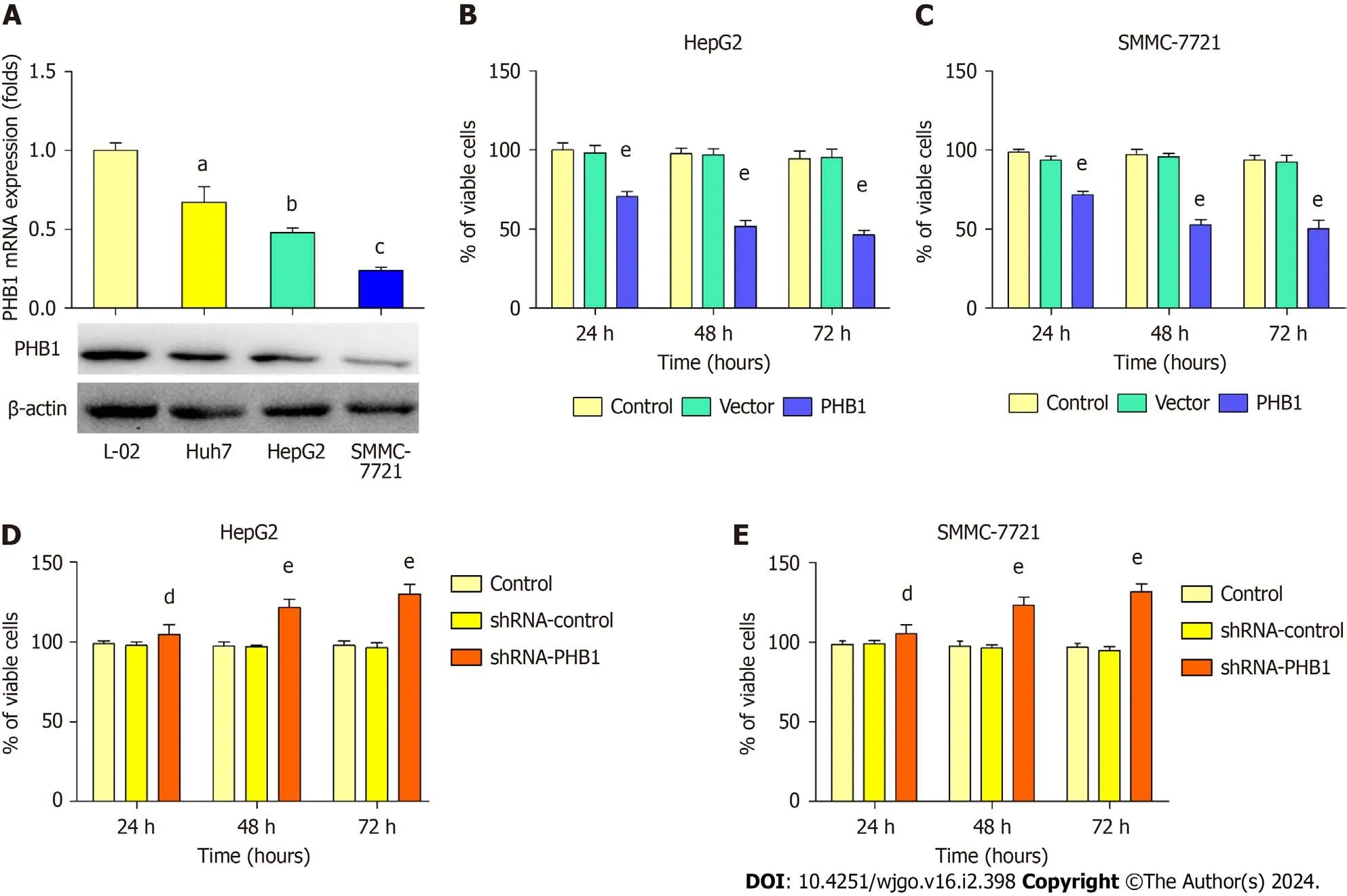
Figure 2 Prohibitin 1 inhibits the proliferation of hepatocellular carcinoma cells. A: The Prohibitin 1 (PHB1) mRNA and protein levels in hepatocellular carcinoma cells were investigated using real-time PCR (upper panel) and Western blot (lower panel),respectively;B-E: HepG2 and SMMC-7721 cell viability was determined using MTT assays after transfection with pEGFP-PHB1 (B and C) or PHB1-specific shRNA (shRNA-PHB1;D and E) for 24,48,or 72 h.The data are presented as the mean ± SD of three independent experiments with similar results.aP < 0.05,bP < 0.01,cP < 0.001 vs L-02 cells.dP < 0.05 and eP < 0.01 vs control and empty vector/shRNA-control,respectively.PHB1: Prohibitin 1.
HepG2 and SMMC-7721 cells were successfully transfected with the pEGFP-PHB1 plasmid and PHB1-specific short hairpin RNA (shRNA-PHB1).Compared with that of the control cells and the empty vector (pEGFP-N1)-transfected cells,the viability of the HCC cells was significantly decreased in a time-dependent manner after transfection with pEGFPPHB1 for 24,48,or 72 h (P< 0.01;Figure 2B and C).However,the HCC cell viability was significantly increased in a timedependent manner when PHB1 was knocked down with shRNA-PHB1 (Figure 2D and E).
PHB1 alters the cell cycle in HCC cells
Because the proliferation of HCC cells was inhibited by PHB1 in a time-dependent manner,we investigated whether PHB1 plays a role in the cell cycle (Figure 3).HepG2 cells were transfected with pEGFP-PHB1 for 48 h,and an increase in the proportion of cells in the G0/G1 (67.27% ± 2.13%vs56.25% ± 1.60% or 58.60% ± 0.91%) and a decrease in the pro-portion of cells in the S phase (14.94% ± 1.60%vs25.40% ± 1.22% or 24.34% ± 1.35%) were observed in HepG2 cells compared with those in untreated control or empty vector-transfected cells (P< 0.01;Figure 3A and B).However,compared with those in untreated control and shRNA-control cells,the proportion of HepG2 cells in the G0/G1 was significantly lower (49.27% ± 1.17%vs54.25% ± 1.21% or 56.60% ± 2.90%),and the proportion of HepG2 cells in the S phase was increased (31.94% ± 1.42%vs25.4% ± 1.60% or 24.34% ± 1.10%) when PHB1 was knocked down with shRNAPHB1 (P< 0.01;Figure 3D and E).Similar results were also obtained with SMMC-7721 cells (Figure 3C and F).

Figure 3 Effect of Prohibitin 1 expression on cell cycle progression of hepatocellular carcinoma cells. A-F: HepG2 and SMMC-7721 cells were transfected with pEGFP-PHB1 (A-C) or shRNA-PHB1 (D-F) for 48 h.The cell cycle distribution was determined by flow cytometry,and the histograms present the mean percentage of the cell population in each phase of the cell cycle.The data are presented as the mean ± SD of three independent experiments with similar results.aP < 0.05 and bP < 0.01 vs control and empty vector/shRNA-control,respectively.PHB1: Prohibitin 1.
PHB1 regulates the cell cycle via the p21/p53-mediated pathway in HCC cells
Because the overexpression of PHB1 arrested the cell cycle in the G0/G1 phase,we investigated the expression levels of cell cycle-related molecules in HCC cells (Figure 4).RT-PCR and Western blot analyses revealed that the p21 and p53 mRNA and protein expression levels were significantly increased but the Cyclin A2,Cyclin E1,and CDK2 mRNA and protein expression levels were significantly decreased in PHB1-overexpressing HepG2 (Figure 4A,E,and G) and SMMC-7721 cells (Figure 4B,E,and G).Conversely,the p21 and p53 mRNA and protein expression levels were lower and the Cyclin A2,Cyclin E1,and CDK2 mRNA and protein expression levels were higher in PHB1-knockdown HepG2 (Figure 4C,F,and H) and SMMC-7721 (Figure 4D,F,and H) cells.
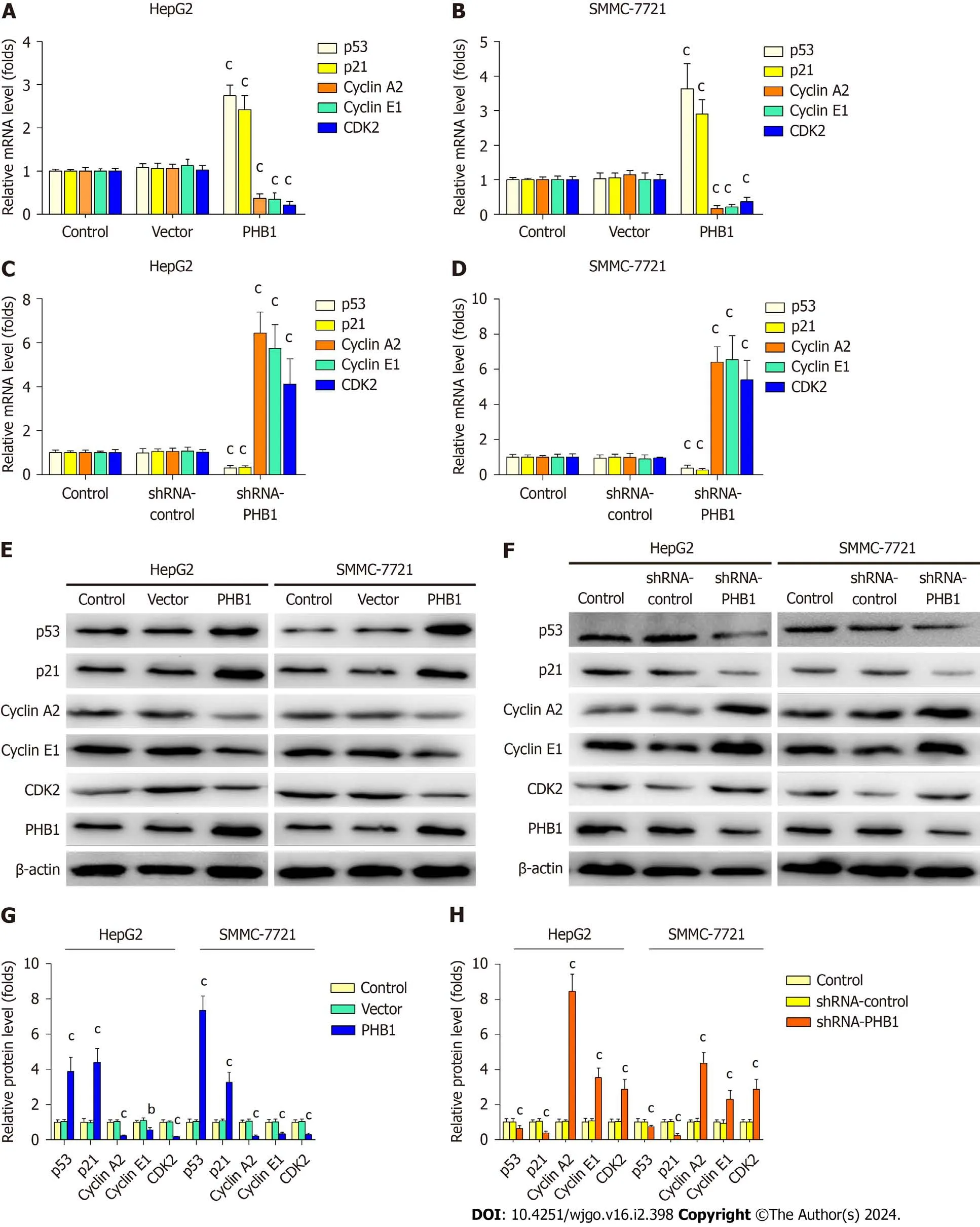
Figure 4 Prohibitin 1 regulates cell cycle progression in hepatocellular carcinoma cells. A-H: The p53,p21,Cyclin A2,Cyclin E1,and CDK2 mRNA and protein levels in HepG2 and SMMC-7721 cells transfected with pEGFP-PHB1 or shRNA-PHB1 for 48 h were determined by real-time PCR (A-D) and Western blot (E-H),respectively.The data are presented as the mean ± SD of three independent experiments with similar results.aP < 0.05,bP < 0.01,and cP < 0.001 vs control and empty vector/shRNA-control.
PHB1 induces apoptosis in HCC cells
Because the proliferation of HCC cells was inhibited by PHB1,we investigated whether PHB1 plays a role in the cell apoptosis (Figure 5).HepG2 cells were transfected with pEGFP-PHB1 for 48 h,and the percentage of apoptotic cells was 15.41% ± 1.06%,which was significantly higher than those of apoptotic untreated control cells (3.65% ± 0.85%,P< 0.01) and empty vector-transfected cells (4.21% ± 0.52%,P< 0.01;Figure 5A and B).However,the percentage of apoptotic cells decreased (6.41% ± 0.36%vs7.65% ± 0.55% and 8.01% ± 0.72%,P< 0.05) in the PHB1-knockdown HepG2 cells (Figure 5D and E).Similar results were also obtained with SMMC-7721 cells (Figure 5C and F).

Figure 5 Effect of Prohibitin 1 expression on hepatocellular carcinoma cell apoptosis. A-F: HepG2 and SMMC-7721 cells were transfected with pEGFP-PHB1 (A-C) or shRNA-PHB1 (D-F) for 48 h.Cell apoptosis was investigated using Annexin V-FITC and PI and was analysed by flow cytometry.The histograms present the mean percentage of apoptotic cells.The data are presented as the mean ± SD of three independent experiments with similar results.aP < 0.05 and bP < 0.01 vs control and empty vector/shRNA-control,respectively.
PHB1 induces apoptosis through activation of the p53-mediated mitochondrial signalling pathway
To determine the mechanism underlying PHB1-mediated apoptosis,we performed RT-PCR and Western blot to investigate the expression levels of apoptosis-related molecules in HepG2 and SMMC-7721 cells (Figure 6).Cyt C release from the mitochondrium into the cytosol is the essential step in the mitochondrial pathway.The protein expression levels of Cyt C were significantly increased in the cytosol and decreased in the mitochondrium when PHB1 was upregulated by pEGFP-PHB1 in HCC cells (Figure 6A and C).However,the expression of Cyt C was not significantly altered in HCC cells when PHB1 was knocked down with shRNA-PHB1 (Figure 6B and C).PHB1 overexpression enhanced the mRNA and protein expression levels of p53,Bax,caspase 3,and caspase 9.In contrast,the Bcl-2 mRNA and protein expression levels were significantly reduced in these HCC cells (Figure 6D,E,H,and J).Conversely,the mRNA and protein expression levels of p53,Bax,caspase 3,and caspase 9 were significantly decreased,and the Bcl-2 mRNA and protein expression levels were increased in HepG2 cells when PHB1 was knocked down with shRNA-PHB1 (Figure 6F,G,I,and K).

Figure 6 Prohibitin 1 is involved in intrinsic mitochondrial signalling pathway activation. A-K: HepG2 and SMMC-7721 cells were transfected with pEGFP-PHB1 or shRNA-PHB1 for 48 h.Cytochrome C protein levels in the cytosol (C) and mitochondrium (M) were determined via Western blot analyses (A-C);p53,Bax,Bcl-2,caspase 3,and caspase 9 mRNA and protein levels were determined by real-time PCR (D-G) and Western blot (H-K),respectively.The data are presented as the mean ± SD of three independent experiments with similar results.aP < 0.05,bP < 0.01,cP < 0.001 vs control and empty vector/shRNA-control.Cyt C: Cytochrome C.
DlSCUSSlON
Abnormal expression of proteins,especially the PHB1 protein,may be closely related to the progression of HCC[20].Several studies have indicated that PHB1 mRNA and protein levels are higher in liver-specificPhb1KO (Mat1aknockout) mice and in normal human hepatocytes,and liver-specific PHB1 deficiency results in marked liver injury,fibrosis,and HCC[21].Further research has shown that PHB1 acts as a negative regulator in murine liver and human HCC cellsviathe WNT-beta-catenin signalling pathway[22].In MHCC97-H cells,which are highly metastatic liver cancer cells,the overexpression of PHB1 inhibits cell proliferation and increases migration[23].However,whether PHB1 is overexpressed in HCC patients is still largely unknown.We examined PHB protein levels in the sera and liver tissues of HCC patients and healthy participants and first found that PHB1 protein expression was significantly higher in HCC patients than in healthy individuals,which is consistent with previous results in patients with gastric cancer[24,25].Because low levels of PHB1 expression were observed in HCC patients,we next observed overexpression of PHB1 in HCC cells,which is consistent with previous results from other cancer cells[14,15,20].
PHB1 plays critical roles in cell survival and apoptosis,but these effects are complex in terms of cell proliferation and apoptosis.Several previous studies have reported that silencing PHB1 induces or accelerates apoptosis in transformed cell lines,which contradicts the conclusions of other studies[26-29].Interestingly,PHB1 knockdown blocks cell proliferation and induces apoptosis in human hepatoma PLC/PRF/5 cellsviathe NF-κB signalling pathway[30].However,PHB1 deficiency results in spontaneous HCC formation in mice,and the knockdown of PHB1 in the mouse normal hepatocyte cell line AML12 increases cyclin D1 and E2F transcription factor expression to accelerate proliferation[21].Therefore,we investigated the effects of PHB1 knockdown and overexpression in HepG2 and SMMC-7721 cells.In the present study,the overexpression of PHB1 inhibited cell proliferation in a time-dependent manner and altered cell cycle progression,but these effects were reversed when PHB1 was knocked down with shRNA-PHB1 in HepG2 and SMMC-7721 cells.
Cell cycle arrest driven by p53 requires the transcription ofGADD45,14-3-3σ,andp21,which are downstream target genes of p53[31,32].Specifically,thep21gene encodes a cyclin-dependent kinase inhibitor,and its transcription is induced by increased p53 protein levels,leading to cell cycle arrest at the G0/G1 phase.G0/G1 phase arrest is principally regulated by p21[32].Several studies have indicated that silencing PHB1 promotes cell proliferation,decreases the proportion of cells in the G0/G1 phase,and increases the proportion of cells in the S phase in the human breast adenocarcinoma cell line MCF-7[33,34].The proportion of cells in the G0/G1 phase was increased and the proportion of cells in the S phase was decreased in PHB1-overexpressing HepG2 and SMMC-7721 cells,which indicates that the overexpression of PHB1 arrested the HCC cell cycle in the G0/G1 phase and thereby inhibited cell proliferation.We further found that the overexpression of PHB1 induced greater expression of p53 and p21,but the knockdown of PHB1 blocked the expression of these molecules in HepG2 and SMMC-7721 cells.The exact mechanism is unclear;however,previous studies on other cell cycle regulatory proteins have shown that rocaglamide (a flavagline) selectively binds to PHB1 and further downregulates cyclin D3,CDK4 (cyclin-dependent kinase 4),and CDK5,which are G0/G1 regulatory proteins,resulting in cell cycle arrest in the G0/G1 phase[35].Therefore,we further investigated whether Cyclin A2,Cyclin E1,and CDK2 mRNA and protein expression levels were significantly decreased following the overexpression of PHB1 in HepG2 and SMMC-7721 cells.PHB1 mainly bound to p53 and increased its expression,which in turn induced p21 transcription and resulted in cell cycle arrest in the G0/G1 phase.
Furthermore,we found that PHB1 induced apoptosis in HepG2 and SMMC-7721 cells.To further investigate the mechanism underlying PHB1-induced apoptosis,we measured the expression levels of apoptosis-related molecules in HCC cells.Because PHB1 plays an important role in the stability of mitochondrial proteins,we speculated that PHB1 induced cell apoptosisviathe mitochondrial signalling pathway.Our studies indicated that the overexpression of PHB1 increased p53,Bax,caspase 3,and caspase 9 mRNA and protein expression but decreased Bcl-2 mRNA and protein expression in HepG2 and SMMC-7721 cells.Conversely,the mRNA and protein levels of p53,Bax,caspase 3,and caspase 9 were significantly decreased,while those of Bcl-2 was increased when PHB1 was knocked down.However,when PHB1 was upregulated in HepG2 cells,Cyt C expression increased in the cytosol and decreased in the mitochondrium.This pattern was indicative of Cyt C release from the mitochondrium into the cytosol,which is the essential step in the mitochondrial pathway[11].Accumulating evidence suggests that p53 can activate the mitochondrial apoptosis pathway by inducing the transcription of the proapoptotic protein Bax through direct protein-protein interactions[36,37].It was recently reported that combined treatment with tetraarsenictetrasulfide and arsenic trioxide induces apoptosis in acute promyelocytic leukaemia cells by increasing the levels of p53 and its target gene Bax[38].The overexpression of PHB1 induced p53 expression and upregulated Bax expression to activate the mitochondrial signalling pathway,further enhancing Cyt C release into the cytosol and activating caspase 9 and caspase 3;these findings are consistent with previous results from other cancer cells[14,15].For the first time,in the present study,we demonstrated that the overexpression of PHB1 significantly inhibited cell proliferation and induced cell cycle arrest and apoptosis in human HCC cells.Our results revealed a novel mechanism by which PHB1 inhibits HCC and showed that the intrinsic mitochondrial pathway is involved in PHB1 overexpression-induced apoptosis in human HCC cells.
CONCLUSlON
In conclusion,we have showed that PHB1 inhibits the viability of human HCC cells by arresting the cell cycle and inducing cell apoptosisviaactivation of the p53-mediated mitochondrial pathway.We propose a working model of the role of PHB1 in inducing apoptosis and activating the p53-mediated mitochondrial pathway.In this model,PHB1 induces the expression of p53 in the nucleus and upregulates Bax expression to activate the mitochondrial signalling pathway,which further enhances Cyt C release into the cytosol and activates caspase 9 and caspase 3.Although additional studies are needed to determine the precise mechanisms by which PHB1 regulates cell apoptosis,our findings reveal a formerly undescribed function of PHB1 in the induction of apoptosis in human HCC cells.
ARTlCLE HlGHLlGHTS
Research background
Prohibitin 1 (PHB1) has been identified as an antiproliferative protein that is highly conserved and ubiquitously expressed,and it participates in a variety of essential cellular functions,including apoptosis,cell cycle control,proliferation,and survival.Emerging evidence indicates that PHB1 may play an important role in the progression of hepatocellular carcinoma (HCC).However,the role of PHB1 in HCC is controversial.
Research motivation
This study aimed to provide a theoretical basis for the use of PHB1 as a possible therapeutic target for liver cancer.
Research objectives
To investigate the effects of PHB1 on cell proliferation,cell cycle progression,and apoptosis in human HCC cells as well as the relevant mechanisms.
Research methods
ELISA,immunohistochemistry,flow cytometry,quantitative real-time PCR,Western blot,and transfection assays were used in this study.
Research results
Low expression of PHB1 was associated with tumour progression in HCC patients.PHB1 inhibited the viability of human HCC cells by arresting the cell cycle.PHB1 induced p53 expression and upregulated Bax expression to activate the mitochondrial signalling pathway,which further enhanced cytochrome C release into the cytosol and activated caspase 9 and caspase 3 to induce apoptosis.
Research conclusions
PHB1 inhibits the viability of human HCC cells by arresting the cell cycle and inducing cell apoptosisviaactivation of the p53-mediated mitochondrial pathway.Our results revealed a novel mechanism by which PHB1 inhibits HCC and showed that the intrinsic mitochondrial pathway is involved in PHB1 overexpression-induced apoptosis in human HCC cells.
Research perspectives
Although our findings reveal a formerly undescribed function of PHB1 in the induction of apoptosis in human HCC cells,additional studies are needed to understand the precise mechanisms by which PHB1 regulates cell apoptosis.
FOOTNOTES
Co-corresponding authors:Shuang-Suo Dang and Wen-Jun Wang.
Author contributions:Dang SS,Wang WJ,and Shi JJ designed and coordinated the study;Shi JJ,Wang YK,Wang MQ,Gao N,Li M,Zhang X,and Liu XT performed the study;Shi JJ,Wang WJ,Li YP,and Jia XL acquired and analysed the data;Shi JJ wrote the manuscript;all authors were involved in the critical review of the results and have contributed to,read,and approved the final manuscript.Dang SS and Wang WJ contributed equally to this study as co-corresponding authors.There were two reasons for designating Dang SS and Wang WJ as co-corresponding authors.First,the research was performed as a collaborative effort,and the designation of co-corresponding authorship accurately reflects the distribution of responsibilities and burdens associated with the time and effort required to complete the study and the resultant paper.This also ensures effective communication and management of postsubmission matters,ultimately enhancing the paper's quality and reliability.Second,Dang SS and Wang WJ contributed efforts of equal substance throughout the research process.The choice of these researchers as co-corresponding authors acknowledges and respects this equal contribution,while recognizing the spirit of teamwork and collaboration of this study.
Supported bythe Key Research and Development Program of Shaanxi,No.2021SF-227 and No.2020SF-297;and the Natural Science Basic Research Program of Shaanxi,No.2023-JC-YB-770.
lnstitutional review board statement:This study was approved by the Institutional Review Board of The Second Affiliated Hospital of Xi’an Jiaotong University and by the Institutional Review Board of the Second Affiliated Hospital of Xi’an Jiaotong University.
Conflict-of-interest statement:The authors have no conflicts of interest to declare.
Data sharing statement:No additional data are available.
Open-Access:This article is an open-access article that was selected by an in-house editor and fully peer reviewed by external reviewers.It is distributed in accordance with the Creative Commons Attribution NonCommercial (CC BY-NC 4.0) license,which permits others to distribute,remix,adapt,build upon this work non-commercially,and license their derivative works on different terms,provided the original work is properly cited and the use is non-commercial.See: https://creativecommons.org/Licenses/by-nc/4.0/
Country/Territory of origin:China
ORClD number:Juan-Juan Shi 0000-0002-5626-9821;Yi-Kai Wang 0000-0001-6709-1777;Mu-Qi Wang 0000-0003-2314-8983;Jiang Deng 0000-0001-8645-3825;Ning Gao 0000-0003-1018-3990;Mei Li 0000-0001-6295-4270;Ya-Ping Li 0000-0002-0900-5559;Xin Zhang 0000-0002-5966-0471;Xiao-Li Jia 0000-0001-8865-9771;Xiong-Tao Liu 0000-0002-7367-6488;Shuang-Suo Dang 0000-0003-0918-9535;Wen-Jun Wang 0000-0001-9861-1763.
S-Editor:Lin C
L-Editor:Wang TQ
P-Editor:Xu ZH
 World Journal of Gastrointestinal Oncology2024年2期
World Journal of Gastrointestinal Oncology2024年2期
- World Journal of Gastrointestinal Oncology的其它文章
- Does enhanced recovery after surgery programs improve clinical outcomes in liver cancer surgery?
- Cardiotoxicity induced by fluoropyrimidine drugs in the treatment of gastrointestinal tumors
- Effect of screening colonoscopy frequency on colorectal cancer mortality in patients with a family history of colorectal cancer
- Preoperative controlling nutritional status as an optimal prognostic nutritional index to predict the outcome for colorectal cancer
- Tumour response following preoperative chemotherapy is affected by body mass index in patients with colorectal liver metastases
- Expression of cyclin-dependent kinase 9 is positively correlated with the autophagy level in colon cancer