
渭北旱塬不同年限撂荒地土壤酶活性及其化学计量变化特征
2024-02-29 09:20雷跻初刘小伟程积民梁1
水土保持研究 2024年1期
雷跻初, 刘小伟, 邓 军, 程 杰, 程积民, 郭 梁1,,7
(1.中国科学院 教育部水土保持与生态环境研究中心, 陕西 杨凌 712100;2.中国科学院大学, 北京 100049; 3.西北农林科技大学 草业与草原学院, 陕西 杨凌 712100;4.宁夏云雾山国家级自然保护区管理局, 宁夏 固原 756000; 5.国家林业和草原局西北调查规划设计院,陕西 西安 710048; 6.西北农林科技大学 黄土高原土壤侵蚀与旱地农业国家重点实验室,陕西 杨凌 712100; 7.中国科学院 水利部 水土保持研究所, 陕西 杨凌 712100)
黄土高原气候干旱、降水稀少、植被贫乏,加之不合理的土地利用加剧了土壤侵蚀,以致黄土高原成为我国乃至世界上水土流失最严重的地区之一,严重影响了当地的生态环境与社会经济。因此,我国于20世纪90年代在黄土高原开展了大量生态修复措施。其中,渭北旱塬坡耕地撂荒是该地区改善土壤条件和恢复退化环境的重要措施[1]。研究表明,耕地在撂荒后,地上植被盖度得到改善[2],水土流失情况有所缓解[3],土壤养分含量显著提高[4]。然而,长期撂荒在改变地上和地下生物群落的同时也会影响生态系统的养分平衡,加之植物与微生物对养分的竞争所导致的土壤微生物养分限制,又会对植被恢复造成负面影响[5],不利于土壤质量的改善和生态系统的稳定性维持。因此,监测撂荒地演替过程中土壤微生物养分限制,辨析其关键影响因素,具有重要研究价值。
土壤胞外酶活性较土壤基本理化性质指标能够更灵敏地反映土壤环境变化,参与碳(C)、氮(N)和磷(P)循环的胞外酶活性的相对比例能体现微生物的养分需求,表征微生物的养分限制情况[6]。土壤微生物主要通过分泌胞外酶转化分解土壤有机质中的C、N和P等元素,这一过程对土壤养分循环和能量流动具有重要调控作用[7]。土壤胞外酶作为土壤养分循环的关键驱动力[8],其中,与C循环相关的酶主要有:β-葡萄糖苷酶(β-1,4-glucosidase, BG)、纤维二糖水解酶(Cellobiohydrolase, CBH);与N循环相关的酶有:β-N-乙酰基氨基葡萄糖苷酶(β-1,4-N-acetylglucosaminidase, NAG)和亮氨酸氨基肽酶(Leucine aminopeptidase, LAP);碱性磷酸酶(Aalkaline phosphatase, AKP)是与P循环相关的关键酶,它们的活性及化学计量特征能有效反映土壤微生物的能量和养分代谢情况[9]。土壤理化性质和养分分布在不同撂荒年限和不同土层间均具有差异[10],这些因素会对土壤酶活性及酶计量比造成直接或间接的影响[11],进而导致不同撂荒年限的土壤微生物养分限制状况有所不同,但具体影响因子及作用途径和机制尚不明确。为了探究渭北旱塬区不同撂荒年限土壤酶活性及其化学计量变化特征,本研究选取5个不同年限撂荒地为研究对象,通过分析土壤养分和胞外酶活性及生态化学计量随撂荒年限的变化,探究不同撂荒年限下土壤养分的变化及微生物养分限制情况及其驱动因素,旨在为渭北旱塬及黄土高原植被恢复和土地资源科学管理提供一定的理论依据。
1 材料与方法
1.1 研究区概况
研究区位于陕西省彬州市永乐镇高辉村(108°06′18″E,35°15′01″N),地处黄土高原中部,属典型渭北旱塬残塬沟壑区。该地区平均海拔为1 040 m,在气候划分上属于温带半干旱气候,年平均温度约9.7℃,平均无霜期180 d,年均降水量561 mm。土壤类型主要为黄绵土,极易受到侵蚀而造成水土流失。当地政府针对水土流失与生态退化进行了长期综合治理,自20世纪90年代开始实施退耕还林还草生态工程以来,大量坡耕地退耕后形成撂荒地,为本研究提供了良好的试验平台。
1.2 样地选择与样品采集
本研究通过空间代替时间的方法来探究撂荒年限对土壤酶活性及其化学计量比的影响。在研究区内选择坡向一致、地形相似、位置相邻的已撂荒5 a,10 a,20 a,25 a和33 a的5个不同年限的撂荒地作为试验样地,每块样地大小为30 m×30 m,各样地之间间距超过50 m,各样地的坡度坡向等情况见表1。样地植被类型多为以蒿类、禾草类为主的草本植物,主要优势种有:铁杆蒿(Lespedezacuneata)、本氏针茅(Stipabungeana)、阿尔泰狗娃花(Heteropappusaltaicus)、达乌里胡枝子(Lespedezadaurica)等。2021年9月在所选样地内进行土壤样品的采集,每个样地随机选取3个间距超过10 m的1 m×1 m的样方作为重复,用内径3.5 cm的土钻分层采集土壤样品,取样深度分别为0—10 cm,10—20 cm和20—40 cm共三层。去除土样中的石块、植物残体、根系和可见的土壤动物后过2 mm筛,将过筛后的土样混合均匀后分为两份,一份储藏于4℃冰箱内用于土壤微生物生物量及酶活性测定,另一份自然风干用于土壤理化性质的测定。
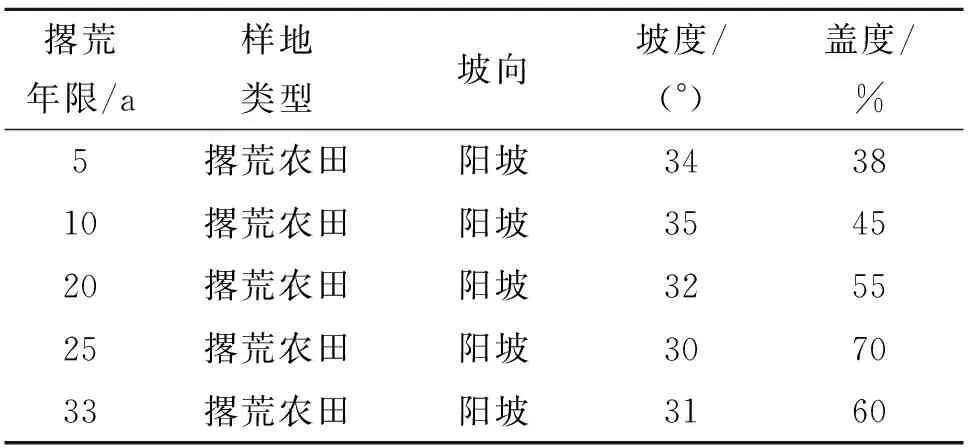
表1 样地概况Table 1 General situation of the sample plots
1.3 测定指标及方法
用重铬酸钾外加热法测定样品中土壤有机碳(Soil organic carbon, SOC)含量,可溶性有机碳(Dissolved organic carbon, DOC)采用TOC法测定[12],采用凯式定氮仪测定总氮(Total nitrogen, TN)含量,总磷(Total phosphorus, TP)含量用浓硫酸—高氯酸钼锑抗比色法测定,用碱解扩散法和NaHCO3浸提—钼锑抗比色法分别测量土壤速效氮(Available nitrogens, AN)和速效磷(Available phosphorus, AP),以上土壤指标测定具体过程参考《土壤农化分析》[13]。
4℃保存的新鲜土样用于测定土壤中5种参与C、N和P循环的酶活性。其中β-葡萄糖苷酶(BG)和纤维二糖水解酶(CBH)是与C循环相关的酶;β-N-乙酰基氨基葡萄糖苷酶(NAG)和亮氨酸氨基肽酶(LAP)是与N循环相关的酶,碱性磷酸酶(AKP)是与P循环相关的酶;所有酶活性采用微孔板荧光法测定[14],详细的酶活性测定方法见参考文献[15]。
1.4 数据处理与分析
通过酶计量矢量模型计算酶化学计量的向量长度(Vector length, VL)和向量角度(Vector angle, VA),量化土壤微生物C、N和P限制,计算公式如下:
x=(BG+CBH)/(BG+CBH+AKP)
(1)
y=(BG+CBH)/(BG+CBH+LAP+NAG)
(2)
(3)
VA=DEGREES〔ATAN2(x,y)〕
(4)
式中:x表示参与C和P循环酶的相对活性比;y表示参与C和N循环酶的相对活性比。向量长度(VL)越长,表明微生物受到的C限制越大;向量角度(VA)小于45°表示微生物受土壤N限制,角度大于45°表示微生物受到土壤P限制,微生物N限制随着角度减小而增大,P限制随着角度增大而增大[16]。
采用R4.1.0软件对试验数据进行统计分析。运用单因素方差分析法(One-way ANOVA)分析不同撂荒年限土壤理化性质、胞外酶活性及其化学计量比差异,采用LSD法多重比较同一土层下各变量在不同撂荒年限间的差异。对酶活性、酶化学计量比与土壤理化性质进行主坐标(PCoA)分析,并利用vegan包中“envfit”函数将土壤理化因子与PCoA轴得分做相关分析以探究影响微生物养分限制的关键土壤因子。
2 结果与分析
2.1 不同撂荒年限土壤养分含量及化学计量比特征
不同撂荒年限下土壤养分含量及化学计量比的结果见表2,撂荒年限对土壤SOC,TN,TP,AN和AP等指标均有显著影响。在0—10 cm土层间,土壤SOC和TN含量随撂荒年限的增加而显著增加,撂荒10 a,20 a,25 a和33 a土壤SOC含量分别较撂荒5 a时增加了17.62%,32.47%,25.55%,2.90%;土壤TN含量分别增加了24.45%,40.42%,31.91%,5.32%。土壤SOC与TN含量均在撂荒20 a达到峰值后有所下降,但总体仍呈现增加的趋势。TP含量则随撂荒年限的增加而显著减少(p<0.05)。土壤AN与AP含量随撂荒年限的增加呈先增后减趋势,总量上仍较撂荒5 a时有所增加。在10—20 cm及20—40 cm土层间,土壤SOC,TN含量均随撂荒年限的增加呈不显著的下降趋势,TP则显著减少(p<0.05)。整体而言,撂荒显著增加了土壤SOC与TN含量,但TP含量会随撂荒年限的增加而减小。
随着撂荒年限的增加,土壤C∶N在各土层中未发生显著改变。土壤C∶P在0—10 cm土层间随撂荒年限的增加而显著增大,在10—20 cm土层间随撂荒年限的增加呈不显著的增加趋势,在20—40 cm土层中随年限的增加而显著降低。土壤N∶P变化随撂荒年限变化并不显著。
2.2 不同撂荒年限的土壤酶活性变化
撂荒年限和土层深度对C、N和P获取的胞外酶活性均具有显著影响(图1)。对0—10 cm土层而言,表征C循环的酶(BG+CBH)和N循环的酶(NAG+LAP)活性在撂荒前期逐年增加,在撂荒20 a达到最大值后开始显著下降,总体呈现出显著的下降趋势。随着撂荒年限的增加,表征P循环的酶(AKP)的活性则呈现升高趋势。在10—20 cm土层,随着撂荒年限的增加,土壤BG+CBH,NAG+LAP酶呈现出先增后减的变化趋势,与浅层(0—10 cm)土壤酶活性变化趋势具有一致性,AKP酶活性随着撂荒年限的增加而显著降低。在更深层土壤(20—40 cm)间,各类土壤胞外酶活性均随着撂荒年限的增加而显著降低,相较于表层,深层土壤酶活性普遍偏低。
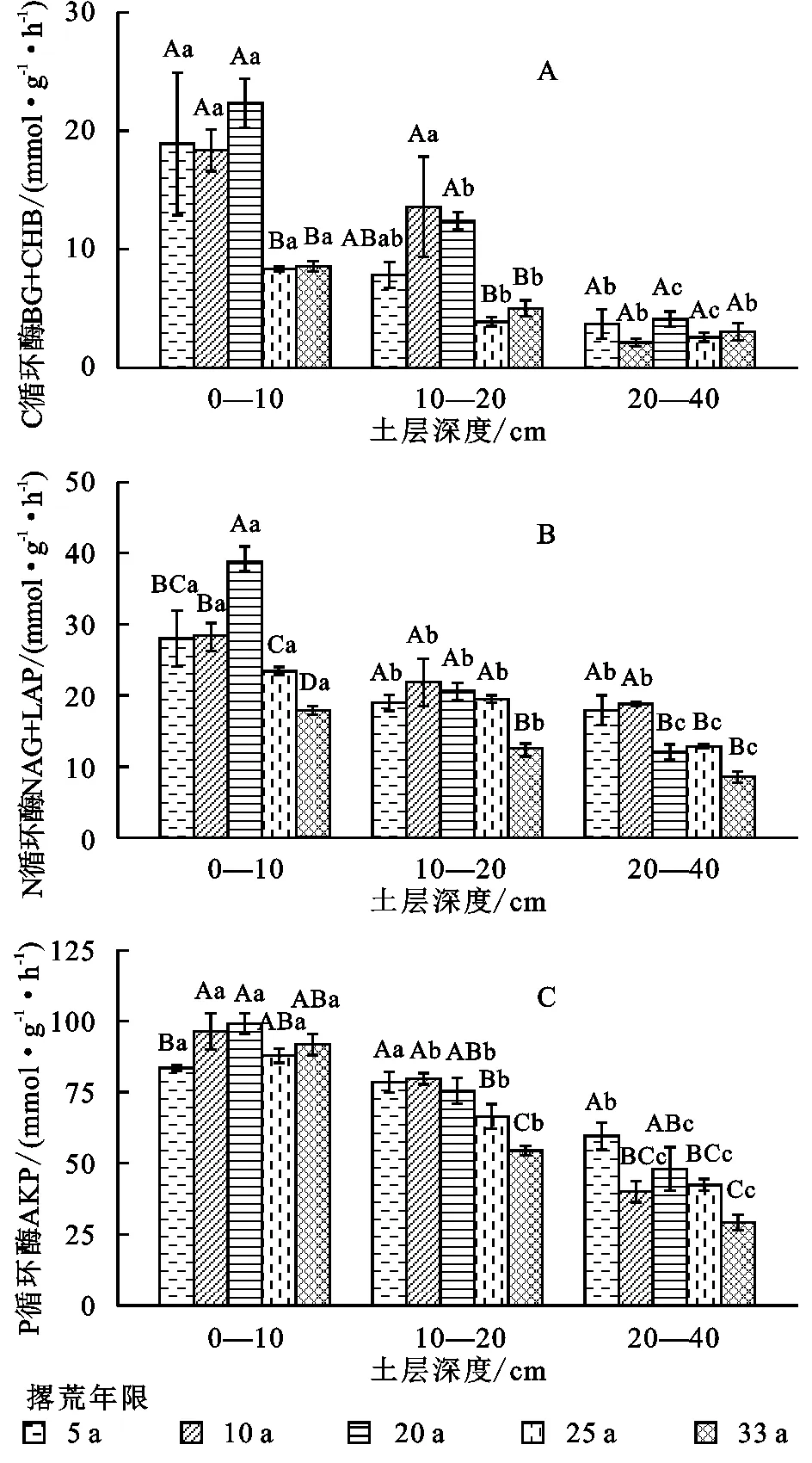
注:不同大写字母表示变量在相同土层不同撂荒年限间差异显著,不同小写字母表示变量在相同撂荒年限不同土层间差异显著(p<0.05),下同。
2.3 不同撂荒年限土壤酶化学计量比变化
对不同撂荒年限下的酶化学计量比进行分析,结果表明:撂荒年限对酶化学计量具有显著影响。整体上,C和N酶活性比(C∶NEEA)、C和P酶活性比(C∶PEEA)和N和P酶活性比(N∶PEEA)在不同撂荒年限土壤中的变化范围分别为0.25~0.87,0.20~0.67,0.69~0.80(图2),且均随着撂荒年限的增加呈显著的降低趋势(p<0.05),这与土壤各类胞外酶活性变化趋势基本一致。
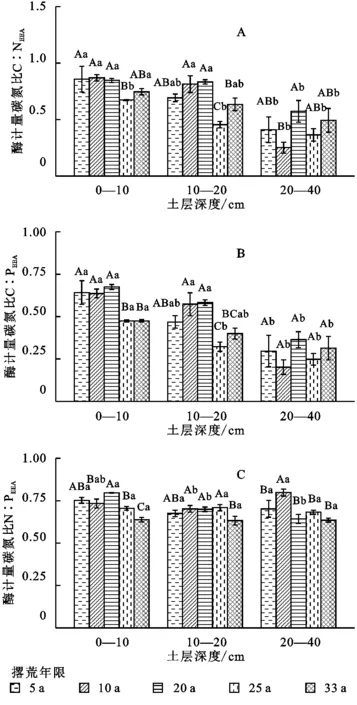
图2 不同撂荒年限土壤酶计量在不同土层中的变化
撂荒年限显著影响土壤微生物养分限制特征(图3)。其中,向量长度(变化范围0.114~0.427)和夹角(变化范围63.27°~75.23°)在不同撂荒年限间差异显著(p<0.05),表明撂荒年限影响了土壤微生物养分限制。在不同土层间,随撂荒年限的增加,向量长度总体呈减小趋势(图3A),表明微生物受到C限制的程度随撂荒年限的增加而减弱。不同撂荒年限下的土壤酶化学计量在不同土层间的向量角度均大于45°,且随撂荒年限的增加而增加,这表明微生物受到强烈的P限制,且其限制程度随撂荒年限增加呈现出增加趋势(图3B,C)。
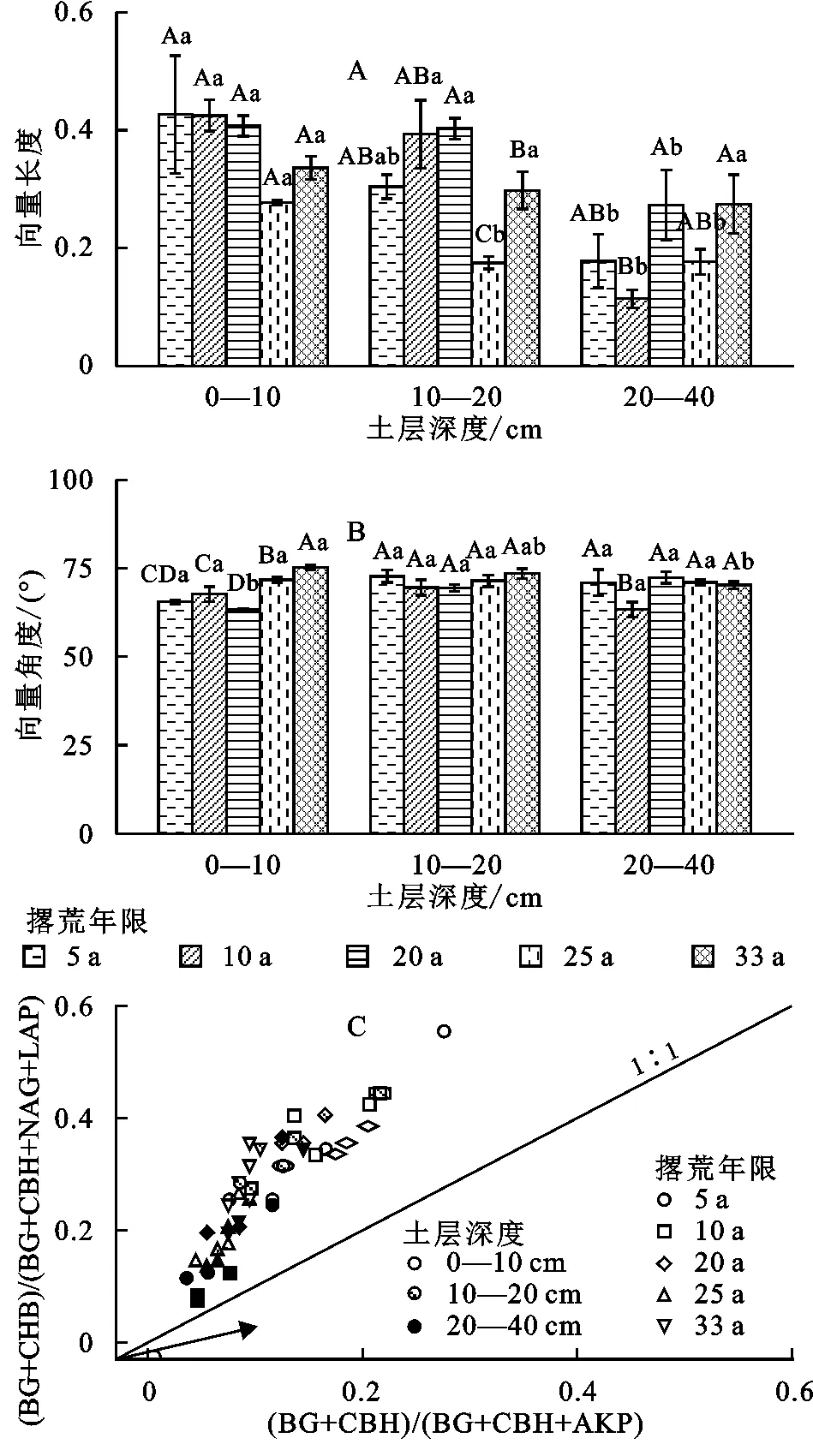
图3 不同撂荒年限土壤酶化学计量的向量长度(VL)和角度(VA)及其计量关系
2.4 不同撂荒年限土壤理化因子对土壤酶活性及其化学计量比的影响
分别以土壤酶活性及酶计量比为响应变量,以土壤理化性质为解释变量进行主坐标(PCoA)分析后,将土壤理化因子与PCoA轴得分做相关分析,得到各土层间影响不同撂荒年限土壤微生物养分限制的关键因子,结果如图4所示。在0—10 cm土层间,影响土壤酶活性及其化学计量比的土壤理化因子为DOC、AN和TP(图4A,D);在10—20 cm土层间,影响土壤酶活性的主要土壤理化因子为TP、AN和AP(图4B),影响土壤酶化学计量比的主要土壤理化因子为DOC和AN;在20—40 cm土层间,影响土壤酶活性的主要土壤理化因子为TP,影响土壤酶化学计量比的主要土壤理化因子为AP。
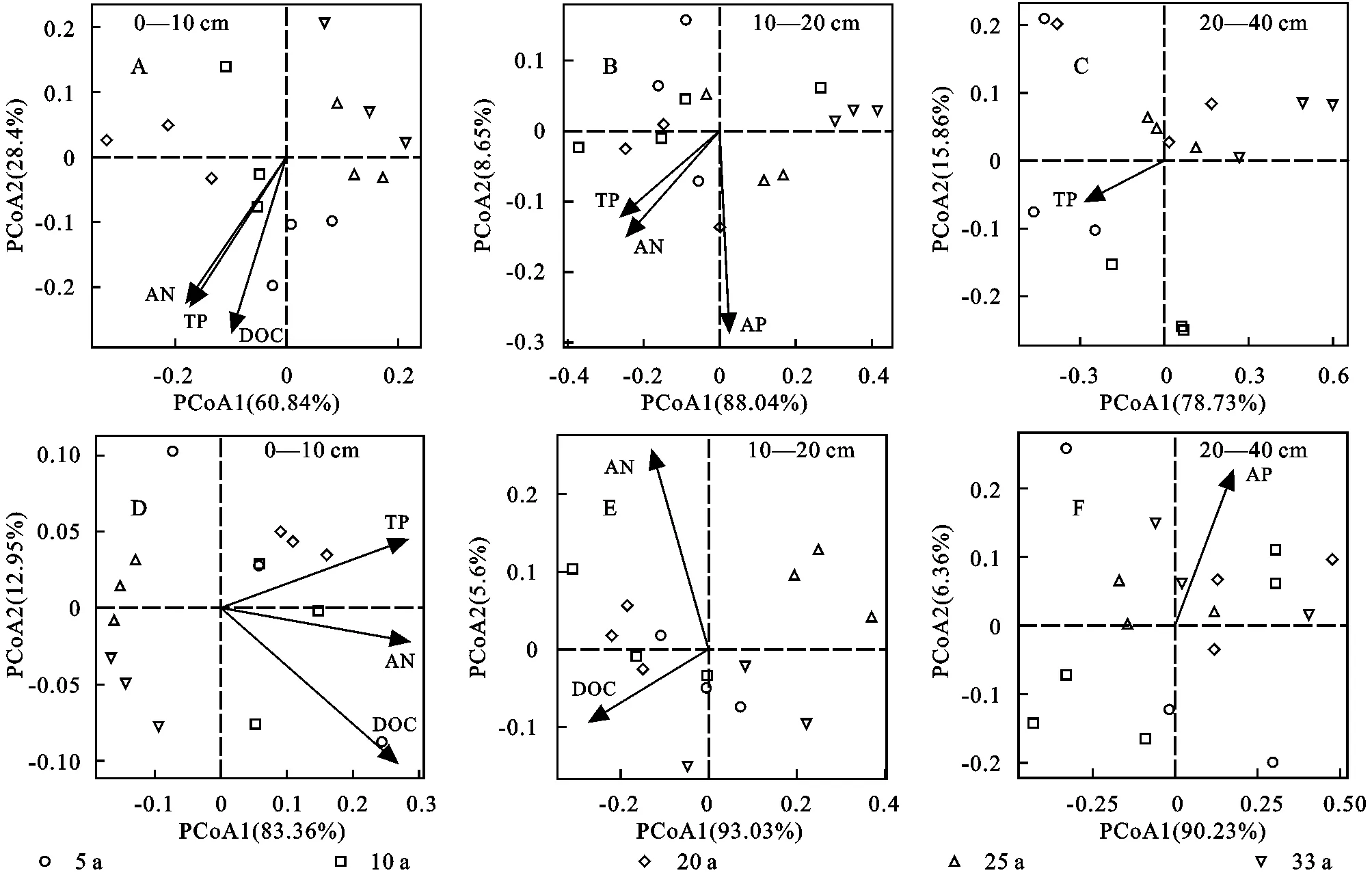
图4 不同撂荒年限土壤酶活性及其化学计量比与土壤养分及其计量比的主坐标分析
3 讨 论
3.1 不同撂荒年限对土壤养分的影响
本研究选取了土壤SOC、TN和TP等指标来反映渭北旱塬地区不同年限撂荒地土壤养分的演变规律(表2),发现土壤养分含量在不同退耕年限之间存在显著差异(p<0.05)。在撂荒前期(5~20 a),土壤SOC和TN含量显著增加,与前人研究结果基本一致[17]。这可能是由于撂荒前期,耕地刚刚停止施肥,土壤有机质及其他养分含量高,增加了地上植物物种多样性和植被覆盖度。同时地上植物凋落物的腐烂分解增加了土壤养分含量,故而土壤C和N含量呈上升趋势。但SOC和TN含量在撂荒20 a达到峰值后又呈现下降趋势,这种动态变化可能是由地上群落在植被演替过程中的变化引起的,有研究表明地上物种多样性和地上生物量在撂荒中期时达到最高,而后下降[18],这与本研究中土壤SOC和TN的变化趋势一致,说明在撂荒后期地上生物量减少,导致土壤有机质输入量降低,造成土壤SOC和TN含量在撂荒20 a后下降,但仍高于撂荒初期的土壤养分含量。土壤TP含量随着撂荒年限的增加而逐年降低,这可能是由于恢复过程植物的生长繁殖造成了土壤P含量的大量消耗,但凋落物分解过程中向土壤输入的P含量较低,导致土壤TP含量下降。
撂荒地土壤SOC、TN和TP等由表层到下层呈现逐层递减的趋势,且表层土壤养分含量显著高于下层,这与前人研究结果一致[19]。这是因为表层土壤受植被枯落物养分归还的影响,因此这些养分在土壤中存在表聚现象。综合来看,渭北旱塬坡耕地撂荒至33 a,土壤C和N含量随着撂荒年限的增加呈现先增后减但总体增加的趋势,而土壤P含量则显著减少。
撂荒年限影响土壤养分变化,不同撂荒年限的土壤C、N和P化学计量比也反映了这种变化。但在本研究中,不同土层间的土壤C∶N变化并不显著,这与现有的研究结果基本一致[20-21]。结果表明,土壤C和N含量间存在内稳态[22],因此土壤C∶N在不同撂荒年限间的变化相对稳定。而土壤C∶P和N∶P总体上呈现增加趋势,意味着土壤P随着撂荒年限的增加而愈发匮乏,从侧面反映出土壤微生物受到了强烈的P限制。
3.2 不同撂荒年限对土壤酶活性及化学计量变化的影响与关键驱动因子
胞外酶在有机质分解和养分循环中起着重要的作用,能在一定程度上反映微生物的生长和代谢过程中的能量(C)和养分限制(N和P)情况[7]。本研究中,在撂荒前期(前20 a),随着撂荒年限的增加,参与土壤C、N和P循环的胞外酶活性显著增加,这与王兴等[23]对黄土高原农田撂荒过程中酶活性变化特征的研究结果一致。有研究表明,胞外酶活性与有机质的分解有关[24],因此耕地撂荒后进入自然恢复阶段转化为草地后,植物凋落物等有机质输入量增加,土壤有机质逐年累积,增加了土壤微生物的养分来源,进而刺激微生物分泌大量的胞外酶[25]。在撂荒20 a以后,群落演替至稳定阶段,地面植被及物种多样性开始降低,且随着撂荒年限的增加土壤养分含量下降,限制了土壤微生物的生长代谢,因此土壤胞外酶活性也随之降低。
土地利用的转变会改变植被盖度和土壤理化性质,微生物代谢受到生物和非生物因子的调节,进而对土壤酶活性及其化学计量比产生影响[26]。本研究发现TP、AN、AP和DOC显著影响土壤酶活性及其化学计量比,说明在不同撂荒年限间这4个因素在调节黄土丘陵区撂荒地土壤酶活性及酶计量比的变化中发挥了主要的驱动作用(图4)。在本研究中,酶化学计量的向量长度随撂荒年限的变化说明微生物受到C限制的程度随着撂荒年限的增加得到了一定程度的缓解,这说明表明耕地撂荒会显著提高土壤SOC含量,进而缓解了微生物C限制。酶化学计量的向量角度变化表明,随撂荒年限的增加,微生物受P限制的程度也随之增加。这是由于黄土高原生物生长对P元素吸收利用率不高[27],同时随撂荒年限的增加而增加的土壤C∶P和N∶P也说明了土壤P含量较之C和N含量的相对不足,进一步加剧了微生物受到P限制的情况。Xiao等[28]的研究表明,在黄土高原次生演替过程中,土壤养分含量与微生物C和P限制显著相关,这与本研究结果一致。
研究发现,随着土层深度的加深,影响土壤酶活性的因子逐层减少。这主要与凋落物和细根残体在土壤中的垂直分布密切关联[29],因为表层土壤较深层土壤含有更多的微生物,会加速植物枯落物的分解,导致土壤中的DOC含量较高,故而DOC成为表层土壤(0—10 cm,10—20 cm土层)酶活性随撂荒年限变化的关键驱动因子,这与DOC在不同土层间的分布情况(表2)相符。同时,土壤DOC的含量会影响土壤微生物可利用C的供给情况[30],本研究中土壤DOC含量随着撂荒年限的增加而逐年减少(表2),促使微生物需要分泌更多的C获取酶来维持自身养分平衡,从而使C循环酶即(BG+CBH)活性在撂荒前期随着撂荒年限的增加而增加(图1A)。
撂荒后土壤N∶P在各土层均随撂荒年限的增加而显著增加(表2),这表明在植被恢复期间P是该生态系统的主要限制因素[31],且土壤酶矢量模型结果也表明微生物受到了较为强烈的P限制,这也与土壤TP含量是限制酶活性和酶计量比的主要因子这一结果吻合。这可能是由于自然演替过程中,随着植被多样性的提高,植物群落对土壤养分的需求量变得更大,导致微生物可利用的养分不足[32],进而促使微生物分泌更多的胞外酶来缓解由于植物竞争引起的养分限制情况。P元素之所以会成为限制酶活性的关键因子,是因为在生态系统中,C和N都可以通过生物途径来进行外源补充,但P元素的含量主要与成土母质有关[1],且其扩散到土壤中的速率较低,随撂荒年限的变化并不显著。但长时间的植被生长会消耗更多的P,这就导致微生物P限制随着撂荒年限的增加而不断加剧。
4 结 论
(1) 撂荒能够显著提高土壤C和N含量,随撂荒年限增加土壤SOC和TN含量呈现先增后减的趋势,且二者均在撂荒20 a时达到峰值后下降,但总体上仍显著增加;土壤P含量随撂荒年限的增加显著降低。
(2) 撂荒显著改变了土壤酶活性:随着撂荒年限的增加,土壤C获取(BG+CBH)、N获取酶(NAG+LAP)活性显著降低,P获取酶(AKP)活性在0—10 cm土层间显著增加,但在更深土层(10—20 cm, 20—40 cm)间显著降低。
(3) 撂荒可以缓解土壤微生物受C限制的程度,但随着撂荒年限的增加,微生物受到P限制的程度反而加剧。土壤DOC、TP和AN含量是驱动酶活性及其计量比变化的关键因子。
综上,本研究通过探究胞外酶活性及其化学计量比与撂荒年限的关系,揭示了自然撂荒条件下植被恢复过程中的微生物养分限制特征。研究表明,撂荒对土壤状况具有改善作用,且浅层土壤中的表现更为显著,但撂荒时间过久(20 a以上)就会加剧微生物P限制,因此对经过长年撂荒的土地应当适量施用磷肥,以期改善其土壤状况。
猜你喜欢
建材发展导向(2022年10期)2022-07-28
现代畜牧科技(2021年4期)2021-07-21
江苏农业科学(2020年2期)2020-04-17
当代陕西(2020年24期)2020-02-01
当代陕西(2020年24期)2020-02-01
当代陕西(2020年24期)2020-02-01
江苏农业科学(2017年10期)2017-07-21
中成药(2017年6期)2017-06-13
动物营养学报(2015年3期)2016-01-07
湖北农业科学(2015年17期)2015-10-09
