
两种入侵植物在海南省的分布格局与风险分析
2024-02-09 00:00:00李晓霞曾力旺曾安逸董定超
南方农业学报 2024年11期
关键词:海南省





摘要:【目的】预测巴西含羞草和无刺含羞草2种入侵植物在海南省的潜在地理分布,确定2种入侵植物在海南省的入侵风险等级,为海南省入侵植物管理和防治工作提供参考。【方法】利用MaxEnt模型和GIS地理信息系统评估制约巴西含羞草和无刺含羞草的主要环境变量,模拟并划分2种入侵植物在海南省的潜在入侵地理分布;开展2种入侵植物定性定量入侵风险分析,确定其危险级别。【结果】模拟结果显示,巴西含羞草和无刺含羞草受试者工作特征曲线面积(AUC)分别为0.956和0.996,结果远高于随机预测值0.500,表明模型预测结果具有较高的可信度。模型预测结果显示,2种入侵植物在我国的适生区主要分布在南方,尤其以广东、海南、福建等省份分布最为密集;其中,在海南省的潜在适宜区域主要集中在东北部,并有向西南扩散的趋势,与实际调查结果分布区域基本吻合。纬度分布预测结果发现,巴西含羞草主要分布在北纬50º~南纬50º,无刺含羞草则主要分布在北纬10º~30º。根据风险指标体系准则层赋值,计算得出巴西含羞草风险值(R)=69.15,高于61.00,为特别危害物种;无刺含羞草R=58.00,介于29.50~61.00,为高度危害物种。【结论】巴西含羞草为特别危害物种,应严格禁止引入;无刺含羞草属于高度危害,必须严格限制引入。由于2种入侵植物已经传入我国,应采取必要的应急措施,开展检疫、监测和灭除等工作,防止其进一步扩散和造成更大的危害。
关键词:巴西含羞草;无刺含羞草;MaxEnt模型预测;入侵风险分析;海南省
中图分类号:S45 文献标志码:A 文章编号:2095-1191(2024)11-3324-09
The distribution patterns and risk analysis of 2 invasive plants in Hainan
LI Xiao-xia, ZENG Li-wang, ZENG An-yi, DONG Ding-chao*
(Institute of Scientific and Technology Information, China Academy of Tropical Agricultural Sciences/Key Laboratory of
Applied Research on Tropical Crop Information Technology of Hainan, Haikou, Hainan 571101, China)
Abstract:【Objective】To predict the potential geographic distribution of Mimosa diplotricha C. Wright and Mimosa diplotricha var. inermis( Adelbert) Veldkamp in Hainan, and to determine the invasion risk level of the 2 invasive plants in Hainan, so as to provide reference for the management of invasive plants and control of invasive plants in Hainan. 【Method】The MaxEnt model and geographic information system( GIS) were used to assess the main environmental varia-bles governing Mimosa diplotricha C. Wright and Mimosa diplotricha var. inermis (Adelbert) Veldkamp, and to simu‐late and delineate the potential geographic distribution of the 2 invasive plants in Hainan; qualitative and quantitative inva‐sion risk analyses of the 2 invasive plants were carried out to determine the level of their risk. The simulation results showed that area under a receiver operating characteristic curve( AUC) of Mimosa diplotricha C. Wright and Mimosa dip‐lotricha var. inermis( Adelbert) Veldkamp were 0.956 and 0.996 respectively, which was much higher than the random prediction value( 0.500), indicating that the prediction results of the model had high confidence.【 Result】The model pre‐diction results showed that in China,the suitable habitats for 2 invasive plants were mainly distributed in the southern re‐gions, particularly in provinces such as Guangdong, Hainan and Fujian. In Hainan, the potential suitable areas were mainly concentrated in the northeastern part and showed a trend of spreading towards the southwest,which was basically consistent with the distribution areas of actual survey results. According to the latitude distribution prediction results, Mi‐mosa diplotricha C. Wright mainly distributed between 50º northern latitude and 50º southern latitude,while Mimosa dip‐lotricha var. inermis (Adelbert) Veldkamp mainly distributed between 10º northern latitude and 30º northern latitude. Based on the risk index system criteria layer assignment, the calculated risk value R for Mimosa diplotricha C. Wright was 69.15, which was higher than 61.00, classifying it as a particularly harmful species; the risk value R for Mimosa dip‐lotricha var. inermis( Adelbert) Veldkamp was 58.00, falling between 29.50 and 61.00, classifying it as a highly harmful species. 【Conclusion】Mimosa diplotricha C. Wright is a particularly hazardous species and its introduction should be strictly prohibited; Mimosa diplotricha var. inermis( Adelbert) Veldkamp is highly hazardous and its introduction must be strictly restricted. Since the 2 invasive plants have been introduced into China, necessary emergency measures should be taken to carry out quarantine, monitoring and eradication, to prevent them from further spreading and causing greater harm.
Key words: Mimosa diplotricha C. Wright; Mimosa diplotricha var. inermis( Adelbert) Veldkamp; MaxEnt model prediction; invasion risk anlysis; Hainan province
Foundation items: Hainan Natural Science Foundation(624MS105); Hainan Philosophy and Social Science Plan‐ning Project[HNSK(YB)23-75]; Young Scholars Project of Hainan Natural Science Foundation(323QN299); Open Fund Project of Key Laboratory of Applied Research on Tropical Crop Information Technology of Hainan in 2023(ZDSYS-KFJJ-202315)
0 引言
【研究意义】随着全球气候变化和人类活动不断扩展,外来物种入侵的数量呈上升趋势,给全球范围内造成了巨大的经济损失(Paini et al.,2016;陶永祥等,2017;Bacher et al.,2018;Masood,2018)。由于外来入侵植物治理难度大,对其进行潜在分布区预测以及建立有效的预警机制,是控制其种群扩张的关键手段和重要基础(李慧琪等,2015;Zhang et al.,2021)。海南是我国最大的热带岛屿,拥有丰富的生物多样性和独特的生态系统,是我国的重要生态安全屏障和生物资源库,同时也是我国入侵植物的重灾区。当前,巴西含羞草(Mimosa diplotricha C. Wright)和无刺含羞草[Mimosa diplotricha var. inermis (Adelbert) Veldkamp]已在海南省海口、儋州、澄迈和三亚等多个市(县)分布,并呈迅猛扩散态势,对海南省的农业生产和生态环境造成了严重损害。因此,了解和评估巴西含羞草和无刺含羞草2种入侵植物在海南省的分布格局和风险状况,对于制定有效的防治策略和保护海南省的生物多样性和生态安全具有重要意义。【前人研究进展】目前,生态位模型[最大熵模型(MaxEnt)、BIOCLIM模型和DOMAIN模型]已被广泛用于预测物种的潜在适生区(Song et al.,2020;Anand et al.,2021)。与其他生态模型相比,MaxEnt模型能准确地模拟大部分外来入侵物种的潜在分布区域,是目前具有较高预测精度的生态位模型(王聪等,2018;Xian et al.,2023)。研究显示,通过MaxEnt模型预测外来入侵物种的潜在分布区域,可以更准确地评估其可能带来的风险和危害,进而制定针对性的防控措施。Yang等(2018)研究入侵植物银胶菊(Galinsoga quadrira‐diata)扩张对气候变化的响应,结果发现,在气候变化条件下银胶菊在中国的适宜区可能减少并向北移动,东北地区入侵风险增加,需紧急采取预防措施。塞依丁·海米提等(2019)运用MaxEnt模型预测刺苍耳(Xanthium spinosum)在新疆的潜在分布格局,结果发现,在2种不同的气候变化情景下,至2070年,刺苍耳分布区中心可能向新疆伊犁州的奎屯方向移动。Arogoundad等(2020)结合MaxEnt模型、遥感数据和环境变量,对夸祖鲁—纳塔尔省的银胶菊进行了模拟,并基于此预测了5种潜在的外来入侵植物在内布拉斯加州北普拉特河地区蔓延的可能性。Zhang等(2021)研究桉树枝瘿姬小蜂(Leptocybe invasa)在中国的潜在分布核心,预测结果有助于识别和遏制其扩散趋势,为制定有效的控制措施提供理论支持。潘铭心等(2022)利用MaxEnt模型预测加拿大一枝黄花(Solidago canadensis L.)在我国的潜在适生区,结果发现,其高度适生区集中在多个省份和沿海地区,为防控该物种入侵提供了重要依据。陈思明(2023)、方铧等(2023)利用MaxEnt模型预测互花米草(Spartina alterniflora)在我国潜在适生区的研究发现,福建漳江口红树林等保护区面临较高的入侵风险。Xian等(2023)对紫茎泽兰(Ageratina adenophora)入侵中国后生态位扩张的研究发现,紫茎泽兰在墨西哥(原产地)和中国(入侵地)之间的生态位重叠度较低,在中国潜在地理分布主要集中在西南地区的热带和亚热带地区,其地理分布格局由降水和温度变量共同决定。李晓霞等(2024)运用MaxEnt模型对小花十万错(Asystasia gangetica subsp. micrantha)的研究发现,小花十万错潜在分布受气候变暖影响,预测其将北迁且集中于华南沿海及高海拔地区,因此,应针对高温多雨、温差小区域制定小花十万错重点防治措施。林敬梧和桂东伟(2024)运用MaxEnt模型预测火炬树(Rhus typhina)在北美和中国的空间分布差异,结果表明,火炬树在北美和中国适宜分布区集中于北纬30°~40°湿润地带,入侵中国后其气候空间和生态位发生较大变化,需加强防疫监测和监管以保护生物多样性。【本研究切入点】野外调查发现,巴西含羞草和无刺含羞草具有广泛的生态适应性和繁殖能力,且能在多种环境中形成单一群落,对本地植物的生长和生态平衡造成威胁。然而,对于巴西含羞草和无刺含羞草在海南省的具体分布格局、风险状况及其与环境因子的关系等方面的研究仍需进一步深入。【拟解决的关键问题】利用MaxEnt模型和GIS地理信息系统评估制约巴西含羞草和无刺含羞草2种入侵植物的主要环境变量,模拟并划分其在海南省的潜在入侵地理分布,并开展2种入侵植物定性定量入侵风险分析,确定其危险级别,为海南省入侵植物管理和防治工作提供参考。
1 材料与方法
1. 1 数据来源及处理
1. 1. 1 巴西含羞草和无刺含羞草分布数据的收集与处理 通过检索Inaturalist、全球生物多样性信息共享平台以及野外实地调查,分别搜集到巴西含羞草标本记录1502条、无刺含羞草标本记录45条。将收集的数据排布至Excel表格中,去除无效样本(相同采集地点和时间的重复样本、地理位置和时间不清楚的样本)。将坐标通过ArcGIS 10.8映射在世界地图上,并通过SDMtoolbox工具的减小空间自变量功能处理,最终得到巴西含羞草位置记录1225条、无刺含羞草位置记录35条,并转化成csv格式进行保存。
1. 1. 2 环境变量的筛选和数据处理 环境气候数据来源于WorldClimate全球气候数据库(https://www.worldclim.org/),包含历史条件下的生物气候变量和未来(2060—2080年)的气候数据变量,气候数据精度为30″。通过联合国粮农组织土壤门户网站(https://www.fao.org/)下载海拔(GloElev)和土壤质量(Sq1~Sq7)等数据。19个生物气候变量环境因子之间有一定的相关性,7个土壤质量因子与植被分布存在一定相关性。利用ArcGIS 10.8中的SDMtoolbox插件Conversion Tools将环境图层转换成MaxEnt可用的ASCII格式。
1. 2 风险评估方法
本研究基于层次分析法(Analytic hierarchy pro‐cess,AHP)原理,参照张瑞海等(2019)、张国良等(2020)提出的外来植物风险指标体系和判断标准,包括4个主要指标层、17个具体指标和32个问题进行量化赋值(张岳等,2022),并计算出入侵植物的风险值(R),即R=0.40R1+0.15R2+0.20R3+0.25R4,式中,R1为入侵性、R2为适应性、R3为扩散性、R4为危害性。根据R值确定危害级别。
2 结果与分析
2. 1 巴西含羞草和无刺含羞草适生区分析
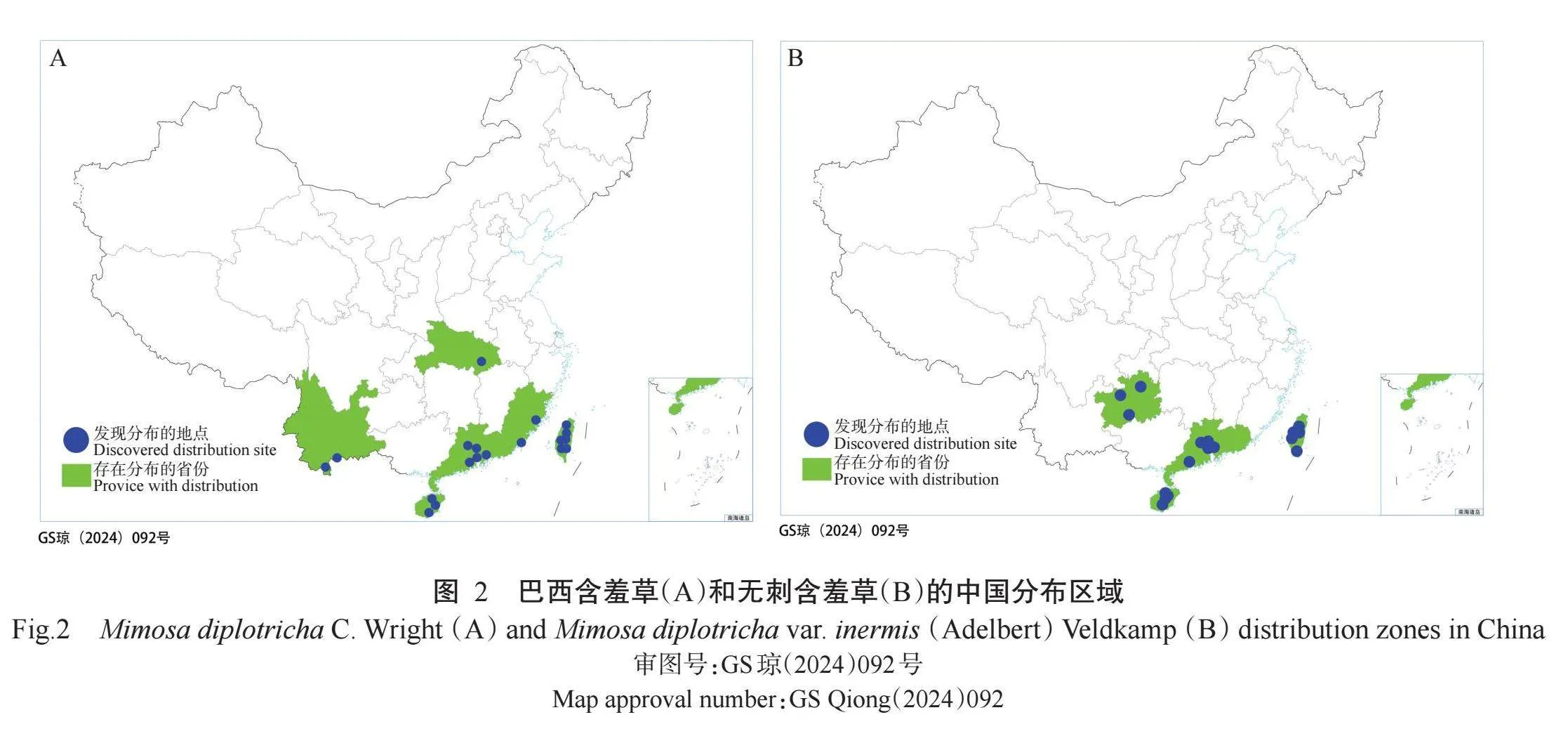
2. 1. 1 当前气候条件下巴西含羞草和无刺含羞草分布 巴西含羞草主要分布于美国、墨西哥、哥伦比亚、委内瑞拉、巴西、印度以及东南亚国家等(图1),我国已确认的分布地有台湾、香港、广州、东莞、三亚和海口等;无刺含羞草则主要分布于我国澳门、深圳、惠州、广州等珠三角城市及海南省(图2)。通过对2种入侵植物生长区域的对比分析,发现巴西含羞草主要分布在北纬50º~南纬50º,无刺含羞草则主要分布在北纬10º~30º;2种入侵植物在我国的适生区主要分布在南方,尤其以广东、海南、福建等省份分布最为密集。
2. 1. 2 巴西含羞草和无刺含羞草在海南省的适生区预测 为进一步提高预测结果的精确性,本研究使用MaxEnt模型进行了5次环境因子的重要性分析,结果显示,最冷月最低温度、最暖季度降水量、最湿季度降水量、最冷季度平均温度和最暖月最高温度等环境因子对2种入侵植物的生长影响较大。2种入侵植物受试者工作曲线面积(AUC)分别为0.956和0.996,结果远高于随机预测值0.500,表明模型预测结果具有较高的可信度。
将MaxEnt得出的预测结果导入到ArcGIS 10.8中,并对栅格数据进行可视化处理。具体如下:0~1的连续栅格数据中,0代表最低生存概率,1代表最高生存概率。生存概率越接近1,表示生存适宜程度越高。采用自然间断点分级法将预测结果划分为多个等级,设定入侵植物生存概率在[0,0.3)区间内的地区为低适生区,生存概率在[0.3,0.7)区间内的地区为中适生区,生存概率在[0.7,1]区间内的地区为高适生区。同时,将生存概率≥0.7作为高适生区的判别阈值,对2种入侵植物的适生性等级图进行二值化处理,并将已入侵区域和潜在入侵区域进行比较分析,结合实地踏查数据,划定2种入侵植物的管控区、高风险入侵区和监测区域。
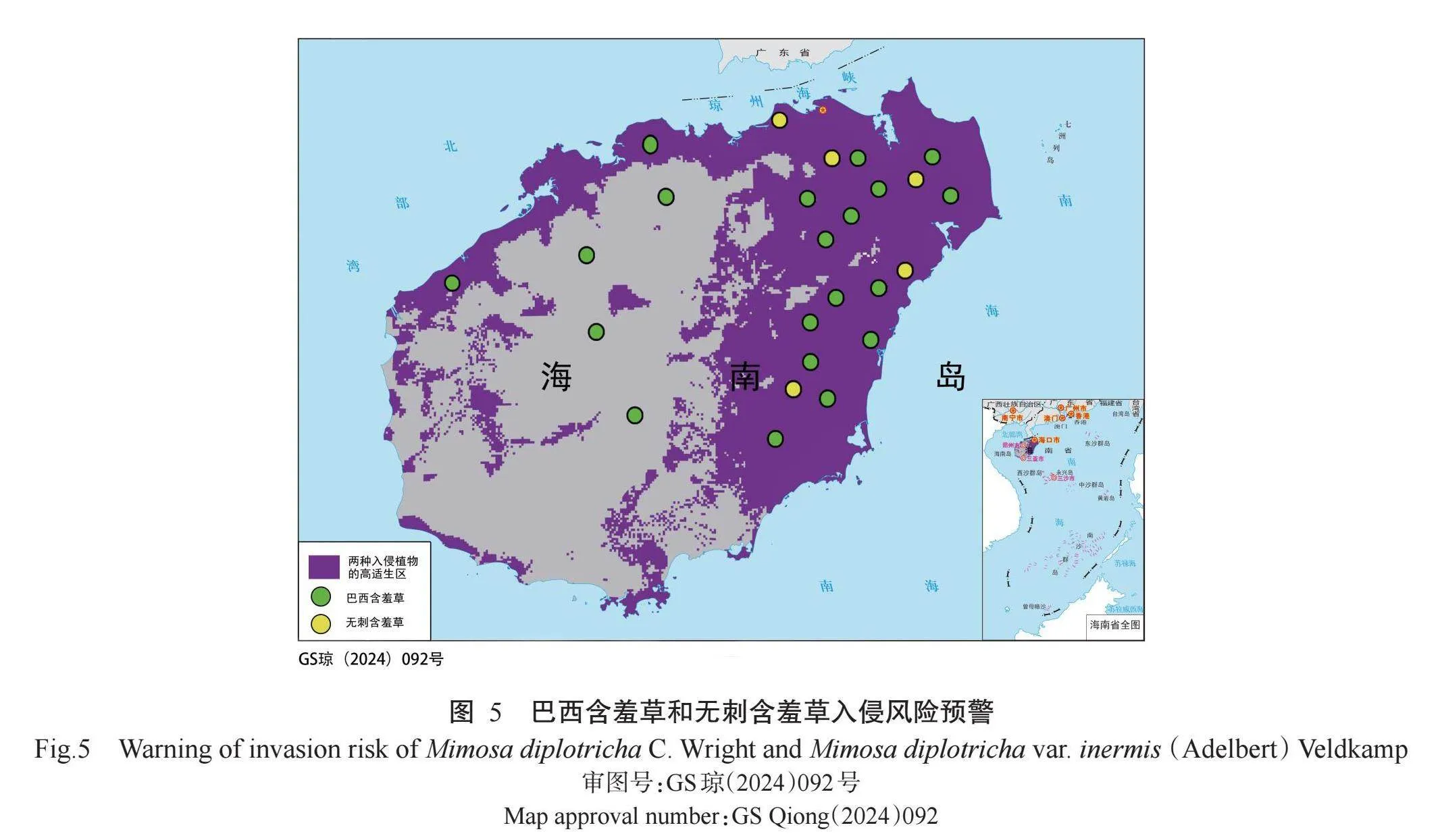
从图3可看出,当前气候背景下,除乐东、三亚、琼中黎族苗族自治县、保亭黎族苗族自治县、五指山市、陵水黎族自治县、白沙黎族自治县和昌江黎族自治县等地2种入侵植物的适生等级存在差异外,其余地区的适生区基本相同。数据库以及实际调研所得到巴西含羞草和无刺含羞草的分布数据基本集中在高热、高湿地区,证明模型预测与2种植物的实际分布吻合,预测结果有较高的可信度。从图3还可看出,巴西含羞草和无刺含羞草入侵风险较大的地区均集中在海南省的东北部地区,但无刺含羞草的入侵风险区域面积更大。
2. 1. 3 影响巴西含羞草和无刺含羞草分布格局的气候因子 为更全面地了解巴西含羞草和无刺含羞草的生存环境需求,本研究分析了2种入侵植物原产地和入侵地的分布数据以及全球环境数据。同时,根据MaxEnt模型运算结果,确定其高适生区4个主要环境因子的变化范围,并计算其在模型中的贡献率。结果(表1)显示,2种入侵植物生长环境因子有较高的相似性,即包含最冷月最低温度、最暖季度降水量、最湿季度降水量、最冷季度平均温度等4个高贡献率环境因子,且4个高贡献率环境因子对MaxEnt模型的总贡献率在70.0%以上,说明上述4个环境因子是影响巴西含羞草和无刺含羞草分布的主要依赖环境因子,且影响因子存在高度相似;同时也表明两者的生长环境相似,因此两者有极大可能生长在同一片区域。
2. 1. 4 巴西含羞草和无刺含羞草在海南省的预警分析 在MaxEnt分析结果基础上对巴西含羞草和无刺含羞草的适宜生境等级图进行叠加处理,识别出高度适宜的生境区域。这一过程产生了代表高适宜区域的栅格数据集,随后将栅格数据集与行政区域图层结合,生成巴西含羞草和无刺含羞草适生性风险预测图(图4和图5)。从图4和图5可看出,巴西含羞草和无刺含羞草已扩散到海南省东部高适生区的边缘,继续向适生度较低的西南方向蔓延。建议在琼海市、万宁市、定安县和澄迈县等市(县)建立阻截带,同时在琼中黎族苗族自治县和陵水黎族自治县等建立入侵检测机制,并且在其他未被入侵地区开展检测调查。
2. 2 巴西含羞草和无刺含羞草入侵风险分析结果
2. 2. 1 定性分析
2. 2. 1. 1 全球范围内的入侵与影响 (1)巴西含羞草原产巴西,为东南亚和太平洋岛屿上常见的严重杂草,广布于非洲中部、太平洋地区多个岛屿、加勒比海地区以及毛里求斯、越南、柬埔寨、马来西亚、印度尼西亚、菲律宾和澳大利亚等地(车晋滇,2010),是太平洋沿岸的一种严重危害杂草。在萨摩亚西部,巴西含羞草已成为农业、牧场、荒地和路边等地的主要杂草;在乌波卢岛,有高达85%的村庄受到巴西含羞草侵袭;在澳大利亚,巴西含羞草已成为热带牧场和北昆士兰甘蔗园的主要杂草;巴西含羞草最初作为花卉被引入我国,随后逸生至野外,其最早的记载可追溯到1956年的《广州植物志》(侯宽昭,1956)和《海南植物志 第二卷》(陈焕镛,1965)。目前,巴西含羞草在我国的广东、广西、海南、台湾、香港和云南等地均有分布。(2)无刺含羞草原产印度尼西亚,现广布于全世界热带地区。《广西植物名录 第二册》(广西植物研究所,1971)《广东植物志 第五卷》(吴德邻,2003)《广西植物志 第二卷》(李树刚,2005)均收载本变种,其标本采集于海南万宁。目前,无刺含羞草在我国澳门、香港、福建、广东、广西、海南和香港均有分布,因其对农业生产和生物多样性的严重影响而被列为我国二级严重入侵物种。
2. 2. 1. 2 传播性与繁殖能力 巴西含羞草和无刺含羞草因具有种子数量多、寿命持久、传播途径多样和繁殖能力强等特性而被广泛传播(韦春强等,2013)。巴西含羞草荚果上有刺毛,易黏附在动物的皮毛上随动物移动而传播;其种子常混杂于各种器物或土壤中,并随交通工具扩散至各个角落;也可借助水流进行传播;此外,人为无意识的携带行为也是其传播的一条途径。无刺含羞草种子可随苗木、草皮交易传播;也可通过人为引种栽培,逸生。巴西含羞草和无刺含羞草在热带、亚热带地区生态适应性强,生长迅速,能在多种土壤和环境条件下生存。根据MaxEnt模型分析结果可知,巴西含羞草和无刺含羞草的适生区包含了海南省的大部分农业产区、山林、生态保护区,故2种入侵植物均可在海南省形成完整的传播链并进行大规模繁殖。
2. 2. 1. 3 经济与生态影响 巴西含羞草和无刺含羞草与农作物竞争光、水、养分和空间,降低农业生产的效率和质量,且在大量生长后影响农业机械的正常运行,增加农业生产的成本和难度;同时,也会破坏土壤结构和肥力,导致土壤侵蚀和退化;此外,巴西含羞草和无刺含羞草还会改变土壤的微生物群落,影响土壤的生物多样性和功能。随着2种植物入侵的加剧,会排挤和取代本地的植物物种,降低植被的多样性和稳定性,甚至影响其他生物,如昆虫、鸟类和野生动物等的生存和繁衍,破坏生态系统的平衡。
2. 2. 1. 4 检疫与防治难易度 巴西含羞草和无刺含羞草由于种子数量多、寿命长、传播途径多、生态适应性强、抗药性强,容易形成单一的群落且难以根除;其种子还可以通过水流、风力、动物、车辆、衣物等方式进行远距离传播,因而难以有效控制(韦春强等,2013)。此外,巴西含羞草和无刺含羞草的植株具有很强的适应性和再生能力,可以在各种环境条件下生长,难以杀灭。目前,巴西含羞草和无刺含羞草的防治方法主要包括物理、化学和生物防治等(马金双,2014)。
2. 2. 2 定量分析 根据外来植物风险指标体系和判断标准,并参照相关外来物种风险评估方法(丁晖等2006;张国良等,2020),提出巴西含羞草和无刺含羞草风险指标体系和判断标准,进行定量分析。该定量分析指标体系准则层包括4个指标层(R1、R2、R3和R4)的17个具体指标和32个问题,根据评价指标的赋分值表(张岳等,2022),设定2种入侵植物R为100,其中R1赋值40、R2赋值15、R3赋值20、R4赋值25。按R=R1×0.40+R2×0.15+R3×0.20+R4×0.25计算风险值,确定危险级别(表2)。通过计算得出巴西含羞草R=69.15,超过了阈值61.00,为特别危害物种;无刺含羞草R=58.00,介于阈值29.50~61.00,为高度危害物种。针对特别危害的巴西含羞草应坚决禁止其引入,以防对本地生态系统造成不可逆的损害;而对于高度危害的无刺含羞草,虽不完全禁止其引入,但应严格限制其数量,并采取足够的监管和防范措施,以控制其逃逸和扩散,确保本地生态安全。
3 讨论
本研究利用MaxEnt模型结合GIS技术分析了巴西含羞草和无刺含羞草在海南省的分布格局,发现2种入侵植物的潜在适宜区域主要集中在海南东北部地区,并有向西南方向扩散的趋势,与其现有的分布区域基本一致。说明MaxEnt模型能较好地预测入侵植物的分布范围,与其他研究者的结论相符(岳茂峰等,2016;李璇等,2018)。MaxEnt模型预测结果还显示,2种入侵植物的分布受到多个环境因子影响,其中最重要的是最冷月最低温度、最暖季度降水量、最湿季度降水量和最冷季度平均温度,这些环境因子与入侵植物的生态适应性有关,也与海南的
气候和地形特征相吻合。
外来植物风险评估包括分析外来入侵植物特性、环境和人为因素,评价其入侵可能性和影响,从社会、经济和政治等方面确定可接受的危害程度,制定和实施降低风险措施(张国良等,2020;张岳等,2022)。本研究对巴西含羞草和无刺含羞草进行了分布、危害及防治等分析,建立合适的风险评估体系,通过定量分析的办法确定其危险级别。研究结果表明,巴西含羞草和无刺含羞草在海南省的入侵风险均较高,其中巴西含羞草的风险略高于无刺含羞草,与2种入侵植物的生物学特性和危害现状等相一致。巴西含羞草具有较强的竞争力、繁殖力和适应力等,当前已在海南省形成了大面积的单一群落,对本地生物多样性和生态系统功能带来严重的威胁。无刺含羞草入侵性虽然没有巴西含羞草强,但也具有较高的生长速度、扩散能力和耐干旱性等特性,且在海南省的分布范围不断扩大,对农业生产和自然保护区造成了不良影响。因此,建议在关键区域建立生物阻截带,并设立长期监测站点;同时,加强公众教育和跨学科合作,政府出台政策法规支持对巴西含羞草和无刺含羞草的管理,控制其扩散,保护本地生物多样性。
本研究使用的入侵植物分布数据是基于文献和网络记录,可能存在一定的误差和不完整性,未能反映2种入侵植物的实际分布状况。因此,有必要通过实地调查和监测来获取更准确和更新的入侵植物分布数据,以提高模型的预测精度和可靠性。此外,本研究使用的环境因子是基于全球尺度的数据集,可能存在一定的粗糙度和不适用性,未能充分考虑海南省的地域特征和微观环境变化。因此,有必要使用更高分辨率和更具代表性的环境因子数据,以提高模型的预测灵敏度和适应性。
本研究使用的定性或定量指标风险评估方法可能存在一定的主观性和模糊性,未能充分反映入侵植物的风险程度和影响范围。因此,有必要使用更客观和综合的风险评估方法,以提高风险评估的科学性和有效性。
4 结论
MaxEnt模型分析结果,巴西含羞草和无刺含羞草在海南省的潜在适宜区域主要集中在海南省东北部地区,与其现有的分布区域基本一致。海南省气候总体上较适合巴西含羞草和无刺含羞草生长,且随着农业生产、动植物活动等因素,其扩散范围仍在不断扩大。根据2种入侵植物的入侵风险分析结果,巴西含羞草和无刺含羞草分别为特别危害物种和高度危害物种,对海南省原生生态环境和农业生产均可能造成不良影响,因此,迫切需要采取措施,开展检疫、监测和灭除等,防止其进一步扩散和造成更大的危害。
参考文献((References)):
车晋滇. 2010. 中国外来杂草原色图鉴[M]. 北京:化学工业出版社. [Che J D. 2010. A color atlas of alien weeds in China[M]. Beijing:Chemical Industry Press.]
陈焕镛. 1965. 海南植物志:第2卷[M]. 北京:科学出版社:204-215. [Chen H Y. 1965. Flora of Hainan,Vol. 2[M]. Bejing:Science Press:204-215.]
陈思明. 2023. 互花米草(Sparina alernilora)潜在分布格局的空间尺度效应[J]. 生态学报,43(14):6058-6068.[ Chen S M. 2023. Spalial seale elect of potential distribution pat‐tern of Sparina alermifora[J]. Acta Eeologica Sinica,43(14):6058-6068.] doi:10.5846/stxb202207162032.
丁晖,石碧清,徐海根. 2006. 外来物种风险评估指标体系和评估方法[J]. 生态与农村环境学报,22(2):92-96.[ Ding H,Shi B Q,Xu H G. 2006. Index system and methodology for risk assessment of alien species[J]. Joumal of Ecology and Rural Environment,22(2):92-96.] doi:10.3969/j.issn.1673-4831.2006.02.020.
方铧,陈星彤,刘明月,张永彬,苗正红,满卫东,张清文,寇财垚,李想. 2023. 基于MaxEnt 模型的互花米草潜在生境适宜性分析[J]. 草地学报,31(11):3514-3524.[ Fang H,Chen X T,Liu M Y,Zhang Y B,Miao Z H,Man W D,Zhang Q W,Kou C Y,Li X. 2023. Analysis on the poten‐tial habitat suitability of Spartina alterniflora based on MaxEnt model[J]. Acta Agrestia Sinica,31(11):3514-3524.] doi:10.11733/i.issn.1007-0435.2023.11.030.
广西植物研究所. 1971. 广西植物名录 第二册[M]. 桂林:广西植物研究所:280.[ Guangxi Institute of Botany. 1971. A checklist of plants in Guangxi,Vol. 2[M]. Guilin:Guangxi Institute of Botany:280.]
侯宽昭. 1956. 广州植物志[M]. 北京:科学出版社:311.[ Hou K Z. 1956. Flora of Guangzhou[M]. Beijing:Science Press:311.]
李慧琪,赵力,祝培文,张嫣,朱耿平,刘强. 2015. 入侵植物长芒苋在中国的潜在分布[J]. 天津师范大学学报(自然科学版),35(4):57-61.[ Li H Q,Zhao L,Zhu P W,Zhang Y,Zhu G P,Liu Q. 2015. Potential distribution of invasive palmer amaranth in China( Amaranthus palmeri)[J]. Jour‐nal of Tianjin Normal University( Natural Science Edition),35(4):57-61.] doi:10.3969/j.issn.1671-1114.2015.04.013.
李树刚. 2005. 含羞草科[M]//中国科学院广西植物研究所. 广西植物志 第二卷. 南宁:广西科学技术出版社:416-441. [Li S G. 2005. Mimosaceae[M]//Flora of Guangxi,Vol. 2. Guangxi Institute of Botany,Chinese Academy of Sciences. Nanning:Guangxi Science and Technology
Press:416-441.]
李璇,李垚,方炎明. 2018. 基于优化的MaxEnt模型预测白栎在中国的潜在分布区[J]. 林业科学,54(8):153-164.[ Li X,Li Y,Fang Y M. 2018. Prediction of potential suitable distribution areas of Quercus fabri in China based on an optimized MaxEnt model[J]. Scientia Silvae Sinicae,54(8):153-164.] doi:10.11707/j.1001-7488.20180817.
李晓霞,胡宽义,曾安逸,董定超. 2024. 基于MaxEnt生态位模型的小花十万错在中国的潜在分布[J]. 西南农业学报,37(4):860-868.[ Li X X,Hu K Y,Zeng A Y,Dong D C. 2024. Potential distribution of Asystasa gangetiea ssp. micrantha in China based on the MaxEnt ecological miche model[J]. Southwest China Journal of Agricultural Seien-ces,37(4):860-868.] doi:10.16213/j.enki.scjas.2024.4.020.
林敬梧,桂东伟. 2024. 火炬树在北美和中国空间分布差异对比及防范入侵措施[J]. 生态学报,44(4):1692-1699.[ Lin J W,Gui D W. 2024. Comparing spatial distribution of torch tree in North America and China and its preventing measures[J]. Acta Ecologica Sinica,44(4):1692-1699.] doi:10.20103/j.stxb.202204100939.
马金双. 2014. 中国外来入侵植物调研报告(下卷)[M]. 北京:高等教育出版社:592-593.[ Ma J S. 2014. The survey reports on Chinese alien invasive plants(Vol. 2)[M]. Bei‐jing:Higher Education Press:592-593.]
潘铭心,朱思睿,张震. 2022. 外来入侵植物加拿大一枝黄花在中国的适生区预测[J]. 西安文理学院学报(自然科学版),25(1):90-96. [Pan M X,Zhu S R,Zhang Z. 2022. Prediction of suitable habitat of invasive plant of Solidago canadensis L. in China[J]. Journal of Xi’an University (Natural Science Edition),25(1):90-96.] doi:10.3969/j.issn.1008-5564.2022.01.016.
塞依丁·海米提,努尔巴依·阿布都沙力克,许仲林,阿尔曼·解思斯,邵华,维尼拉·伊利哈尔. 2019. 气候变化情景下外来入侵植物刺苍耳在新疆的潜在分布格局模拟[J]. 生态学报,39(5):1551-1559.[ Sayit H,Nurbay A,Xu Z L,Arman J,Shao H,Vinira Y. 2019. Simulation of potential distribution patterns of the invasive plant species Xanthium spinosum L. (Bathurst burr) in Xinjiang under climate change[J]. Acta Ecologica Sinica,39(5):1551-1559.] doi:10.5846/stxb201802040299.
陶永祥,赵建伟,王兰新,郭贤明. 2017. 西双版纳自然保护区外来入侵植物现状调查[J]. 山东林业科技,(1):58-61. [Tao Y X,Zhao J W,Wang L X,Guo X M. 2017. Investi‐gation on the present situation of alien invasive plants in Xishuangbanna Nature Reserve[J]. Shandong Forestry Scinence and Technology,(1):58-61.] doi:10.3969/j.issn. 1002-2724.2017.01.014.
王聪,蔡普默,仪传冬,陈家骅. 2018. 2007—2017年外来入侵害虫风险评估的文献分析及常用风险评估模型介绍[J]. 中国农业大学学报,23(8):225-238.[ Wang C,Cai P M,Yi C D,Chen J H. 2018. Bibliometric based analysis of the risk assessment of alien invasive pest from 2007 to 2017 and introduction of several risk assessment models[J]. Journal of China Agricultural University,23(8):225-238.] doi:10.11841/i.issn.1007-4333.2018.08.24.
韦春强,赵志国,丁莉,潘玉梅. 2013. 广西新记录入侵植物[J]. 广西植物,33(2):275-278. [Wei C Q,Zhao Z G,Ding L,Pan Y M. 2013. Newly recorded invasive plants in Guangxi[J]. Guihaia,33(2):275-278.] doi:10.3969/i.issn.1000-3142.2013.02.026.
吴德邻. 2003. 含羞草科[M]//中国科学院华南植物研究所. 广东植物志 第五卷. 广州:广东科技出版社:141-163. [Wu D L. 2003. Mimosaceae[M]//South China Institute of Botany,Chinese Academy of Sciences. Flora of Guang‐dong,Vol. 5. Guangzhou:Guangdong Science and Tech‐nology Press:141-163.]
岳茂峰,冯莉,崔烨,张纯,田兴山. 2016. 基于MaxEnt模型的入侵植物白花鬼针草的分布预测及适生性分析[J]. 生物安全学报,25(3):222-228. [Yue M F,Feng L,Cui Y,Zhang C,Tian X S. 2016. Prediction of the potential distri‐bution and suitability analysis of the invasive weed,Bidens alba( L.) DC[J]. Jouranl of Biosafety,25(3):222-228.] doi:10.3969/j.issn.2095-1787.2016.03.013.
张国良,付卫东,孙玉芳,王忠辉,张瑞海. 2020. 外来入侵物种监测与控制[M]. 北京:中国农业出版社.[ Zhang G L,Fu W D,Sun Y F,Wang Z H,Zhang R H. 2020. Monito-ring and control of alien invasive species[M]. Beijing:China Agriculture Press.]
张瑞海,张国良,宋振,王忠辉,付卫东. 2019. 印加孔雀草入侵生态风险评估及其管理措施[J]. 生物安全学报,28(1):71-75.[ Zhang R H,Zhang G L,Song Z,Wang Z H,Fu W D. 2019. The invasive assessment and management measure of Tagetes minuta L[. J]. Journal of Biosafety,28(1):71-75.] doi:103969/i.issn.2095-1787.2019.01.013.
张岳,张国良,袁至立,宋振,王忠辉,付卫东. 2022. 速生槐叶蘋入侵风险评估及管理措施分析[J]. 生物安全学报,31(4) :364-374.[ Zhang Y,Zhang G L,Yuan Z L,Song Z,Wang Z H,Fu W D. 2022. Invasive risk assessment and management measure analysis of Salvinia molesta D. S. Mitchel[l J]. Journal of Biosafety,31(4):364-374.] doi:10.3969/j.issn.2095-1787.2022.04.010.
Anand V,Oinam B,Singh I H. 2021. Predicting the current and future potential spatial distribution of endangered Rucer‐vus eldii eldii( Sangai) using MaxEnt mode[l J]. Environ‐mental Monitoring and Assessment,193:147-165. doi:10.1007/s10661-021-08950-1.
Arogoundade A M,Odindi J,Mutanga O. 2020. Modelling Parthenium hysterophorus invasion in KwaZulu-Natal Province using remotely sensed data and environmental variables[J]. Geocarto International,35(13):1450-1465. doi:10.1080/10106049.2019.1581268.
Bacher S,Blackburn T M,Essl F,Cenovesi P,Heikkila J,Jeschke J M,Jones C,Keller R,Kenis M,Kueller C,Marti‐nou A F. Nentwig W,Perl J,Pyšek P,Rabitsch W,Richard‐son D M,Roy H E,Saul W C,Scalera R,Vila M,Wilson J R U,Kumschick S. 2018. Socio-economic impact classifi‐cation of alien taxa (SEICAT)[J]. Methods in Ecology and Evolution,9(1):159-168. doi:10.1007/10265-015-0738-3.
Masood E. 2018. The battle for the soul of biodiversity[J]. Nature,560(7719):423-425. doi:10.1038/d41586-018-05984-3.
Paini D R,Sheppard A W,Cook D C,De Barro P J,Worner S P,Thomas M B. 2016. Global threat to agriculture from invasive species[J]. Proceedings of the National Academy of Sciences,113(27):7575-7579. doi:10.1073/pnas.1602 205113.
Song X Y,Milne R I,Fan X Y,Xie S Y,Zhang L,Zheng H L,Fan L Q,Chung J M,Chung M G,Ma T Xu X T,Wang J,M K. 2020. Blow to the Northeast?Intraspecific differen‐tiation of Populus davidiana suggests a north-eastward skew of a phylogeographic break in East Asia[J]. Journal of Biogeography,48(1):187-201. doi:10.1111/jbi.13992.
Xian X G,Zhao H X,Wang R,Zhang H B ,Chen B X,Liu W X,Wan F H. 2023. Evidence of the niche expansion of crofton weed followinginvasion in China[J]. Ecology and Evolution,13(1):e9708. doi:10.1002/ece3.9708.
Yang Y Bo,Liu G,Xiong S,Zhang W G,Cai X W. Ren Z L . Yao N N,Zhu Z H,Nie H. 2018. Where will invasive plants colonize in response to climate change:Predicting the invasion of Galinsoga quadriradiata in China[J]. Inter‐national Journal of Environmental Research,12:929-938. doi:10.1007/s41742-018-0146-3.
Zhang H,Song J Y,Zhao H X,Li M,Han W H. 2021. Predic-ting the distribution of the invasive species Leptocybe invasa:Combining MaxEnt and geodetector models[J]. Insects,12(2):92-109. doi:10.3390/insects12020092.
(责任编辑 麻小燕)
猜你喜欢
小主人报(2023年1期)2023-03-15 01:08:24
新教育(2022年16期)2022-06-22 07:04:04
党的生活(黑龙江)(2022年4期)2022-04-25 22:14:17
资源信息与工程(2021年5期)2022-01-15 05:37:32
中国财政年鉴(2018年0期)2018-07-08 08:12:20
成都信息工程大学学报(2017年4期)2018-01-22 02:08:33
中国卫生(2016年12期)2016-11-23 01:09:50
中国财政年鉴(2016年0期)2016-06-05 15:23:31
中国市场(2016年12期)2016-05-17 05:10:28
中学历史教学(2015年7期)2015-11-11 07:08:26
