植物次生代谢的区室化研究进展
2024-02-05 06:12关智晶孙超
生物技术通报 2024年1期
关智晶 孙超
(1. 天津中医药大学,天津 301617;2. 中国医学科学院药用植物研究所,北京 100193)
植物次生代谢产物不仅可以作为防御分子对抗自然环境中的生物和非生物胁迫,而且许多次生代谢产物具有显著的生物活性,为医药、农业、化工、食品和化妆品等行业提供了宝贵原料,对人类的生产生活具有重要意义[1]。次生代谢途径中一系列化学反应的顺利完成需要特定的反应条件,区室化正是植物应对这一难题的巧妙方法。此外,植物次生代谢区室化过程中还需要转运蛋白参与跨膜运输。近年来,随着单细胞转录组、空间转录组和质谱成像等新技术的出现,极大地提高了我们对于次生代谢空间分布的解析能力,区室化研究已经成为次生代谢研究领域里的一个新热点。
1 植物次生代谢在不同水平上的区室化
植物次生代谢存在多层次的区室化现象,这些不同的区室构成了相互作用的区室系统,是植物为了生存和抵抗环境胁迫,长期适应性进化的结果,对植物生长发育具有重要意义。同时区室化策略还能为代谢工程和合成生物学研究提供新的思路[2]。例如,Shi等[3]发现人参皂苷合成的关键酶原人参二醇合酶(protopanaxadiol synthase, PPDS)在酵母中异源表达时定位于内质网膜上,而其底物达玛烯二醇存在于脂滴当中。研究人员通过将PPDS靶向脂滴中,为酶反应建立了新的化学区室,从而显著提高了人参皂苷的生产效率,底物转化率达到86%。
1.1 分子水平的区室化
代谢区室(metabolon)是植物次生代谢中最小的区室,是由催化“级联”反应的酶通过非共价作用聚合而成的瞬时多酶复合体。在代谢区室内可形成底物通道,防止不稳定中间产物的降解和副反应的发生,提高酶的催化效率。代谢区室的形成可以增强途径反应,而代谢区室的解离可以下调途径反应,通过代谢区室的聚合和解离,植物可以对代谢流进行动态调节[4]。目前,已发现多种次生代谢产物的合成过程中存在分子水平的区室化现象,包括苯丙素类、生氰糖苷、萜类和生物碱等[5]。例如,蜀黍氰甙(dhurrin)是一种由高粱合成的具有防御功能的生氰糖苷类化合物。在植物体内,细胞色素P450CYP79A1和CYP71E1、糖苷转移酶UGT85B1能够形成代谢区室,催化L-酪氨酸合成蜀黍氰甙(图1-A)[6]。此外,苯丙素合成途径中的苯丙氨酸解氨酶(PAL)和肉桂酸-4-羟化酶(C4H)可以形成代谢区室,催化L-苯丙氨酸生成香豆酸[7]。
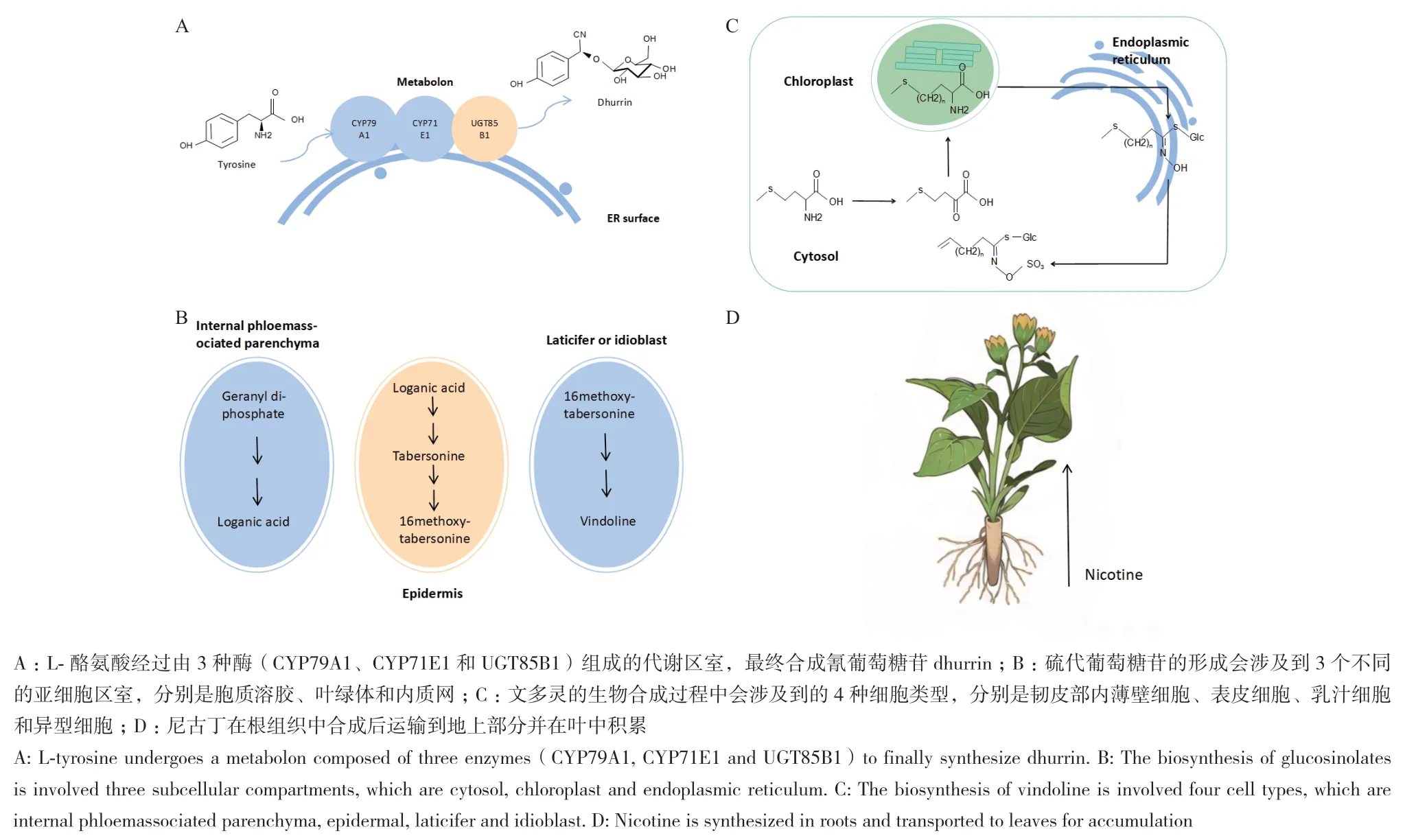
图1 植物不同水平上的次生代谢区室化Fig. 1 Plant specialized metabolic compartmentalization on different levels
1.2 亚细胞水平的区室化
由于次生代谢途径中的酶和中间产物往往会定位于不同细胞器中,所以次生代谢会在亚细胞水平上出现区室化现象。例如,抗癌药物长春碱和长春新碱在长春花叶中的生物合成至少涉及5个亚细胞区室,分别是细胞质、液泡、内质网、叶绿体和细胞核[8]。硫代葡萄糖苷在植物体内的生物合成途径分为3个阶段:R侧链的延伸、核心结构的合成和侧链的修饰。首先,甲硫氨酸在胞质溶胶中发生由BCAT4催化的初始脱氨反应,随后进入叶绿体中完成侧链延伸,之后在内质网中形成脱硫硫苷,脱硫硫苷运输至胞质溶胶中形成硫苷核心结构,最后继续在胞质溶胶中进行侧链修饰,形成硫代葡萄糖苷[9](图1-B)。有些化合物可以在多个亚细胞区室中产生,例如,所有萜类物质的共同前体异戊烯焦磷酸可以在细胞质中通过甲羟戊酸(MVA)途径合成,也可以在质体中通过甲基赤藓糖醇磷酸(MEP)途径合成[10]。除了常规的细胞器之外,苯质体(phenyloplasts)和单宁体(tannosomes)分别是酚类和缩合单宁(高聚原花青素)的生产和储存的特殊细胞器[11-12]。
1.3 细胞水平的区室化
每个细胞都是一个单独的区室,每种细胞类型都有自己的代谢活动,次生代谢途径会在不同的细胞类型之间进行分工[13]。例如,长春花中长春碱的生物合成过程中涉及到的酶反应分布在4种不同的细胞类型当中,起始于韧皮部内薄壁细胞,中间产物马钱苷酸从韧皮部内薄壁细胞转移至表皮细胞当中,乳汁细胞和异形细胞是文多灵和其他单萜吲哚类生物碱合成的场所[14](图1-C)。罂粟中吗啡的生物合成过程至少涉及韧皮部的3种细胞类型,分别是伴胞、筛管分子和乳汁细胞。吗啡的两种合成原料多巴胺和四羟基乙醛会在筛管分子中形成7-氧二氢蒂巴因,一部分7-氧二氢蒂巴因会在筛管分子中继续生成蒂巴因(筛管分子中的途径酶都是在伴胞中合成后转移至筛管分子的),随即蒂巴因会通过苄基异喹啉摄取通透酶(BUPs)家族转移至乳汁细胞中完成吗啡的合成;而另一部分7-氧二氢蒂巴会在BUPs家族的介导下直接转移至乳汁细胞中合成最终产物[15]。
1.4 组织器官水平的区室化
植物体的器官是植物体内最大的区室,次生代谢产物的储存常常集中在某种器官当中,例如青蒿素主要储存在黄花蒿的叶片中,杜仲绿原酸主要存在于杜仲叶片中,人参皂苷主要存在于人参和三七的根中,这些积累的器官通常为药用植物的“药用部位”。尽管植物能在特定的器官积累次生代谢产物,但是积累的地点并不总是合成的地点。植物体内存在一个将“源”组织器官中合成的次生代谢产物运输到“库”组织器官中的转运系统[13]。例如,木薯中产生的生氰糖苷在叶片中合成,最终会运输到根部。黄连在根中产生小檗碱,随后会转移至根茎当中。拟南芥中硫代葡萄糖甙在运输到种子之前在叶片和角果中产生。尼古丁是一种烟草属的吡咯烷生物碱,主要在根组织中合成,随即被运输到地上部分并在叶中积累[16](图1-D)。
2 次生代谢产物的跨膜转运
次生代谢的区室化必然伴随着中间代谢产物的跨膜运输,转运蛋白在次生代谢产物的跨膜运输过程中发挥着关键作用。目前已报道的参与植物次生代谢产物转运的转运蛋白来源于4个蛋白家族,分别是ATP结合盒转运蛋白(ABC)、多药和毒素外排家族蛋白(MATE)、硝酸盐/多肽转运家族蛋白(NPF)和嘌呤通透酶(PUP)。
2.1 ABC转运蛋白对植物次生代谢物的转运
ABC蛋白家族转运蛋白是数量最多、功能最广泛的超家族蛋白之一。高等植物中的ABC蛋白分为8个亚家族[1],参与次生代谢转运的转运蛋白主要集中在ABC B、ABC C和ABC G 三个亚家族中,转运的次生代谢产物包括生物碱、黄酮类化合物、萜类化合物、挥发性化合物和类胡萝卜素等。目前已鉴定的ABC G亚家族转运蛋白都是负责向胞外的跨细胞膜运输;ABC C亚家族主要负责花青素类物质由胞质向液胞内的跨膜运输;ABC B亚家族既可以向细胞内,也可以向细胞外运输次生代谢产物。
2.2 MATE转运蛋白对植物次生代谢物的转运
MATE转运蛋白家族成员几乎存在于所有的植物中,含量丰富且具有各种各样的功能,参与次生代谢产物和植物激素的运输、外源性物质的解毒等[17]。据报道,MATE家族成员参与了生物碱、黄酮类和萜类化合物等次生代谢产物在不同区室之间的转运。定位于液泡膜上的转运蛋白负责向液泡内转运次生代谢产物,定位于细胞膜上转运蛋白向细胞外运输次生代谢产物。
2.3 PUP转运蛋白对植物次生代谢物的转运
先前研究认为植物中的PUP转运蛋白家族主要参与含嘌呤环化合物(腺嘌呤、胞嘧啶和细胞分裂素)和含吡啶环化合物(吡哆醇和吡哆醛)的运输[1]。除此之外,现在已经有研究证明PUP蛋白及其同源物NUP1(主要介导尼古丁Nicotine转运)和BUP1(主要介导苄基异喹啉类生物碱BIA转运)也参与着植物中次生代谢产物——生物碱的运输,这些转运蛋白都定位于细胞膜上,参与生物碱向细胞内的跨膜运输。
2.4 NPF转运蛋白对植物次生代谢物的转运
NPF是一个大的转运蛋白家族,这些蛋白质最初被认为是一类在植物硝酸盐吸收和运输中具有独特功能的转运蛋白[18],目前多项研究证明,NPF转运蛋白也在次生代谢产物的转运中发挥作用。高等植物中NPF家族分为8个亚家族NPF1-8[19],其中已鉴定的参与次生代谢转运的转运蛋白主要集中在NPF1和NPF2两个亚家族中,转运的次生代谢产物包括硫代葡萄糖苷、生氰糖苷、黄酮醇苷和生物碱。NPF1蛋白负责向液泡外的跨膜运输,NPF2蛋白大多向细胞内转运次生代谢产物,也有少数向液泡外运输(表1)。
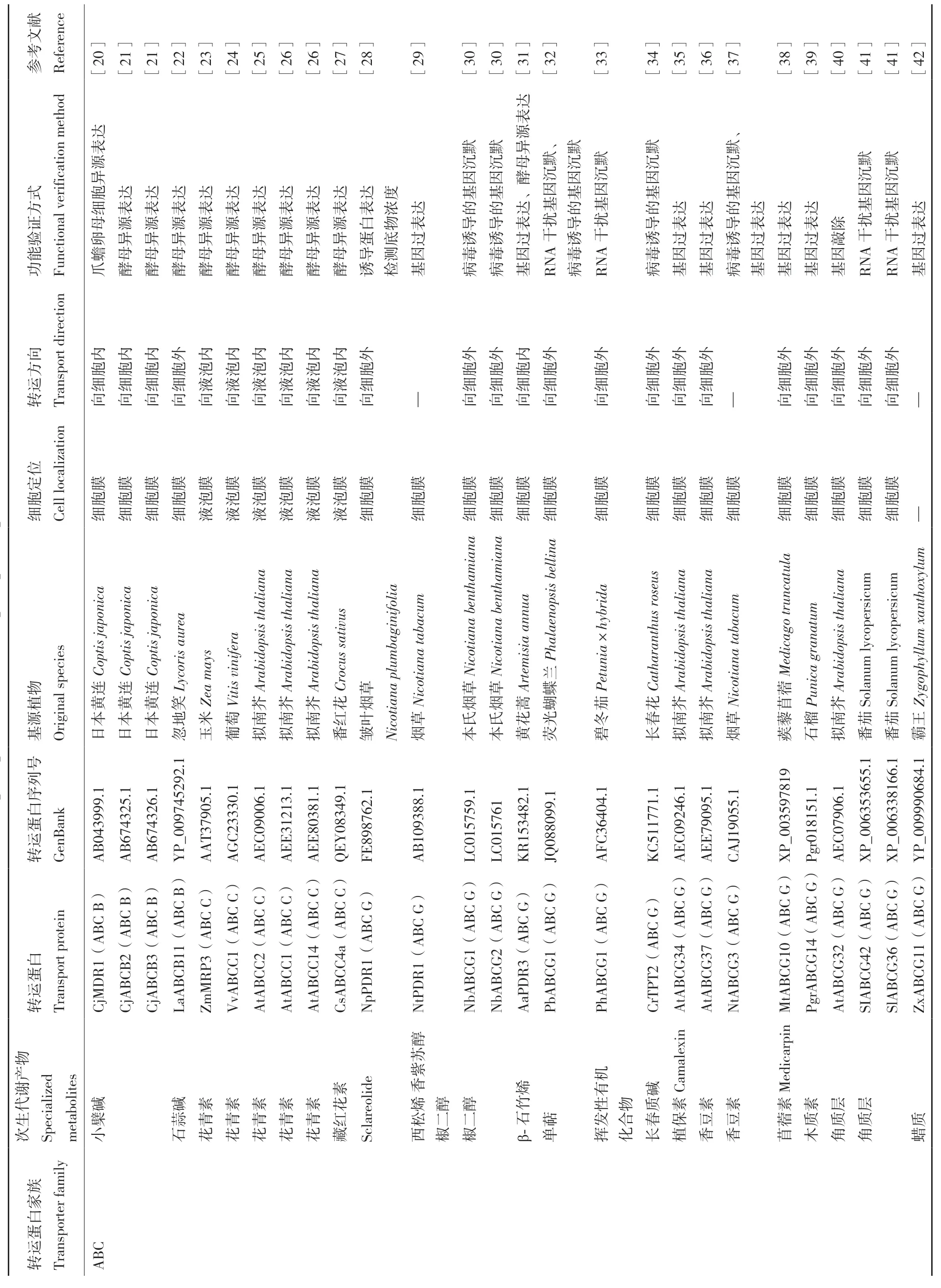
表1 运输植物次生代谢产物的转运蛋白Table 1 Transport proteins that transport plant specialized metabolites
3 次生代谢区室化对植物产生的影响
次生代谢的区室化现象在植物界中广泛存在,可以有效降低酶抑制效应、控制代谢流的大小和方向、减轻中间产物的细胞毒性、提高中间产物的稳定性、分散细胞的代谢负担、防止不必要的代谢串扰,在植物的生长发育和胁迫响应中发挥着重要作用。
3.1 降低酶抑制效应
代谢产物可以抑制产生代谢产物的酶,这是一种在真核生物中普遍存在的现象,而区室化最大限度的减少了这种不可避免的酶抑制,并减轻了自我抑制对次生代谢的限制[72]。Alam 等[72]将酶定位到不同细胞器,来研究区室化对抑制剂网络的影响,研究结果表明次生代谢是通过利用亚细胞区室减少酶的抑制作用。
3.2 减轻中间产物的细胞毒性
植物中所产生的次生代谢产物一般以低浓度形式存在,因为某些次生代谢产物的浓度升高会产生细胞毒性[1]。而为了避免这种自身毒性,植物中已经进化出存在各种各样的解毒机制。例如,细胞外排、液泡隔离、囊泡运输、胞外生物合成以及以无毒形式积累代谢产物等[73]。可以看出,区室化作用在这些解毒机制中占了很大的比例。例如在长春碱合成过程中,参与单体缩合反应的酶,即异胡豆苷合酶和过氧化物酶,定位在液泡中,这表明细胞将产生的有害终产物隔离在液泡中,从而保护细胞免受自身毒性[74]。
3.3 提高中间产物的稳定性
区室化可以将次生代谢途径中不能稳定存在的中间体转化为稳定的中间体。如L-酪氨酸经过由3种酶(细胞色素P450酶CYP79A1和CYP71E1、葡萄糖苷转移酶UGT85B1)组成的代谢区室最终合成dhurrin。在这个过程中,L-酪氨酸先通过CYP79A1和CYP71E1转化为氰醇,氰醇在细胞质基质中不稳定,之后通过UGT85B1酶可以转化为稳定的氰苷[4]。
3.4 防止不必要的代谢串扰
区室化可以有效防止次生代谢途径之间的相互干扰[75]。例如,异戊二烯是所有萜类物质的前体,植物可以通过细胞质中的MVA途径和质体中的MEP途径合成异戊二烯,细胞质中合成的异戊二烯主要用于倍半萜和三萜的合成,而质体中的异戊二烯主要用于单萜和二萜的合成,从而有效避免了不同萜类物质合成之间的相互干扰。
4 植物次生代谢区室化及相关转运蛋白的应用研究
对植物次生代谢区室化的深入研究为通过生物技术提高次生代谢产物含量提供了新的思路和策略。利用生物技术将代谢途径限制在质体和线粒体等亚细胞区室内,可以有效提高酶和底物的浓度,减少支路代谢和复杂调控网络的干扰,提高目标产物的产量。例如,Wu等[76]通过将法尼基焦磷酸合酶与紫穗槐二烯合酶定位到烟草叶绿体上,使得紫穗槐二烯的产量提高了1 000倍以上。植物次生代谢转运蛋白的发现为转运工程和合成生物学研究提供了新的靶点。通过删除一些有毒或有害物质的转运蛋白基因可以降低植物组织中相应物质的积累,提高农作物或其他经济作物的品质。例如,Nour-Eldin等[77]通过突变删除油菜的GTR转运蛋白基因,可以使油菜种子中的抗营养性物质硫代葡萄糖苷的含量下降了60%-70%。
在天然产物合成生物学研究中,通过在底盘细胞中表达植物转运蛋白或改造底盘细胞的转运体系,可以将合成的次生代谢产物转运到培养基中或隔离到细胞器中,降低产物对细胞的毒性,并减轻产物对酶的反馈抑制作用,从而增加次生代谢产物的产量。例如,Yamada等[78]在大肠杆菌中导入番荔枝碱生物合成途径的基础上,再转入具有番荔枝碱转运活性的拟南芥转运蛋白AtDTX1发现,番荔枝碱的产量增加了11倍。通过模拟植物细胞水平的区室化,微生物的模块化共培养工程将代谢途径划分成不同的模块,在不同的细胞中表达,可以有效减轻细胞的代谢负担,减少产物的反馈抑制,从而提高目标产物的产量。例如,Zhou等[79]通过大肠杆菌和酿酒酵母的共培养系统成功合成了氧化紫衫烷类物质,产量可达33 mg/L。
5 总结与展望
随着多组学技术的快速发展,植物次生代谢产物的合成与调控研究取得了巨大进步,长春碱和诺斯卡品等一些重要天然药物的生物合成途径相继得到解析。但是目前对于植物次生代谢的研究主要集中在途径相关酶和转录因子的发掘和鉴定方面,对于大多数次生代谢途径的关注和认知还停留在级联的酶促反应阶段。但是在植物中次生代谢途径往往具有复杂的空间组织结构,代谢中间产物需要在不同区室之间进行运输,这种空间分布会对次生代谢流的方向和大小产生重要影响。虽然对于次生代谢空间分布的研究还比较匮乏,但是随着单细胞转录组、空间转录组和质谱成像等新技术的出现,为次生代谢空间分布研究提供了有力的工具。例如,本研究团队利用单细胞转录组技术成功对长春碱生物合成的空间分布进行了研究,表明单细胞和空间多组学技术在研究次生代谢空间分布方面具有巨大的应用潜力。
植物次生代谢的区室化在植物中普遍存在,对其进行深入研究可以为代谢工程和合成生物学研究提供新的策略和思路。目前,已经有研究人员利用区室化策略,通过质体工程、转运蛋白工程和共培养等方式,在原植物或微生物底盘中高效合成目标天然产物。但是区室化策略在合成生物学中的应用还有许多亟待探索的问题。例如,植物次生代谢产物转运蛋白在微生物底盘系统中的亚细胞定位和转运活性是否与植物中相同;如何构建非天然的代谢区室以提高代谢途径的反应效率;在底盘系统中如何通过多个转运蛋白的协同作用控制代谢流的大小和方向;以及如何通过多层次的区室化协同提高目标产物的产量等。
猜你喜欢
小哥白尼(军事科学)(2019年9期)2019-12-21
中学生数理化·七年级数学人教版(2019年10期)2019-11-25
天然产物研究与开发(2019年10期)2019-11-05
电影(2019年3期)2019-04-04
中学生数理化·高一版(2018年9期)2018-10-09
湖南教育·C版(2018年3期)2018-06-05
阅读(低年级)(2018年11期)2018-05-14
少儿科学周刊·少年版(2017年3期)2017-06-29
中国民族医药杂志(2016年2期)2016-05-14
河北医科大学学报(2011年1期)2011-03-25