
Stratigraphy of the Alveolina elliptica group from the Middle Eocene of Iran: Calibration with calcareous nannofossils biozones and description of Alveolina ozcani n.sp.
2024-01-25 07:26:28MehiHiLorenzoConsortiAliBhrmiMohmmPrnvrSeeKhorrmel
Journal of Palaeogeography 2024年1期
Mehi Hi , Lorenzo Consorti , Ali Bhrmi ,*,Mohmm Prnvr , See Khorrmel
a Department of Geology, Faculty of Sciences, University of Isfahan, Isfahan, Iran
b Institute of Marine Sciences (ISMAR-CNR), Area Science Park, Basovizza, 34149 Trieste, Italy
c National Iranian Oil Company, Exploration Directorate, Sheikh Bahaei Square, Tehran, Iran
d Department of Geology, Faculty of Science, Ferdowsi University of Mashhad, Mashhad, Iran
Abstract Precise taxonomy and the chronostratigraphic calibration of the Middle Eocene Alveolina from Central Iran is here undertaken from the Chah-Talkh section of the southern Sabzevar region(Central Iran).We have identified Alveolina kieli, Alveolina stercusmuris and Alveolina nuttalli along with the new species Alveolina ozcani n.sp.that we include into the Alveolina elliptica group.We have also found Nummulites uroniensis,Nummulites obesus and Nummulites cf.verneuili and associated calcareous nannofossils that look reliable to make thoughtful correlations with the Shallow Benthic Zones (SBZ).The foraminiferal biostratigraphy suggests an assignment to the upper part of the lower Lutetian—lower part of the middle Lutetian,SBZ13 (Middle Eocene), further strengthened through the identification of the calcareous nannofossil NP14b—NP15b or CNE8—CNE10 biozones,providing a solid correlation with the global stratigraphic standards.
Keywords Alveolina, Calcareous nannofossil, Central neo-tethys, Central Iran, Middle Eocene,Biostratigraphy
1.Introduction
The Sabzevar region of Central Iran represents a challenging area for retrieving significant successions holding Cenozoic larger foraminifera.This is especially related to the tectonic and magmatic events occurred through its geological history that have complicated the good preservation of larger-foraminifera-rich shallow-water limestone outcrops (e.g.Hadi et al.,2019a, 2021; Deveciler et al., 2022; Jafari and Ghasemi, 2023).These successions usually show a limited thickness and/or a scarce vertical continuity,with consequent difficulties to recover precise stratigraphic information.However,although scattered and rare, some of these outcrops host abundant larger foraminifera that if studied in detail could bear important taxonomical and stratigraphic implications,especially about the relationships with coeval larger foraminifera from the western Tethys platforms.Alveolina is one of the most significant genera with a remarkable diversification rate from the Late Paleocene (Thanetian) to the late Middle Eocene (Bartonian) and a wide geographical distribution (Hottinger,1963; Sirel and Acar, 2008; Drobne et al., 2011)throughout the tropical and subtropical shelves of the Neo-Tethys Ocean (e.g.Hottinger, 1960; Zhang et al.,2013).Alveolina is polyphyletic, characterized by a high number of parallel linages characterized by increase in biometric parameters through time,including the size of the test and proloculus and the number of chambers per whorl (Hottinger, 1963,1974).The growth of body size through time could be considered in accordance with the Cope's rule, or based on phenotypic effects(Brasier,1984;Hottinger,2001; Guex, 2003; Hadi et al.,2021).Representatives of the Alveolina elliptica group sensu Hottinger(1974)have been globally reported along the Neo-Tethys carbonate platforms (e.g.Hottinger, 1960, 1974;Drobne, 1977; Boukhary, 1985; Samanta, 1993; Serra-Kiel et al., 2016; Haig et al., 2019; Hadi et al.,2020a; Rahman et al., 2021).Hottinger (1974)thought that there is no practical reason to split the Alveolina elliptica group into many different taxa and recognized only two species (A.stercusmuris and A.elliptica), regarding all the other morphotypes with flosculinized basal layer such as A.elliptica nuttali and A.pillai as synonyms of A.elliptica.Samanta (1993),however,had a different view supporting the contrary.Such a discrepancy is likely due to the lack of a comprehensive framework of Alveolina phylogeny that,even though introduced by Hottinger(1963),has still not been fully worked out.Therefore,additional,globally distributed stratigraphic successions should be investigated, in order to extract a sort of standard trend through time(Hottinger and Drobne,1988;Hadi et al., 2021).
Investigations of the central Neo-Tethys biogeographic province of Iran dealt with some Lower Eocene Alveolina assemblages (e.g.Hadi et al.,2019b, 2019c, 2020b, 2021), but the Middle Eocene remains poorly known.Thus, the aim of the present contribution is to undertake a detailed study of the Alveolina occurrences in the Sabzevar region,including Alveolina ozcani n.sp.We have corroborated their stratigraphic occurrences by using the Shallow Benthic Zones (SBZs) of Serra-Kiel et al.(1998) and a detailed micropaleontological study of the standard calcareous nannofossil biozones for correlation with the global chronostratigraphic scale(Geological Time Scale, Gradstein et al., 2020).References for Paleogene calcareous nannofossils include Martini (1971; NP biozones), Okada and Bukry (1980;CP biozones), Varol (1989; NNT zones), and Agnini et al.(2014; CNP/CNE/CNO biozones) that integrate biozones with the geomagnetic polarity chrons(Berggren et al., 1995; Serra-Kiel et al., 1998;Berggren and Pearson, 2005).SBZs are built by Oppelian zones that integrate parallel scales of different systematic groups (Alveolina, Nummulites,orthophragminids and soritids, among others) and correlations with the global standards, although recalibrations might be still necessary for some intervals (Serra-Kiel et al., 2016; Pignatti and Papazzoni, 2017; Silva-Casal et al., 2021; Rodriguez-Pint′o et al., 2022; Benedetti et al., 2023).In the central Neo-Tethys, the Eocene Iranian platform for instance, may not so straightforwardly correspond with the SBZs (Pignatti and Papazzoni, 2017).Alveolina may shows of a certain degree of paleogeographydriven biases (related to endemism and/or timing of dispersal rate) that would lead to discrepancies in stratigraphic distributions (e.g., Hadi et al., 2020b,2021; BouDagher-Fadel and Price, 2021) when comparing the ranges of the same species recovered from distant successions.To the contrary,Nummulites seems less influenced by such factors and could be used,depending on each individual case,as a control against those Alveolina species that may show such influence (Racey, 1992; BouDagher-Fadel and Price,2014).By the integration of independent biostratigraphic scales we aim to confirm the reliability of key Nummulites species for correlation of widely spaced shallow-water successions, whereas west—east correlations based on Alveolina may need to be improved and considered case-by-case with greater care.
2.Geological setting
The Iranian platform was involved in the Alpine-Himalayan orogeny and has been classically subdivided into eight sedimentary—structural provinces,each one characterized by a different geological history (St¨ocklin, 1968).From the north to the south these are: (1) the Kopet-Dagh zone in the NE Iran, (2)the Alborz zone in N-NW Iran,(3)the Central Iran zone(CIZ), (4) the eastern Iranian suture zone, (5) the Urumieh-Dokhtar (Sahand-Bazman) magmatic arc, (6)the Zagros zone,(7)the Sanandaj-Sirjan zone,and(8)the Makran zone(Fig.1A).The geological history of the CIZ is perhaps the most complex one (Fig.1A).The studied section is located in the Sabzevar magmatic belt of the CIZ, which is a fossil volcanic arc mainly composed of Cenozoic volcanoclastic deposits intercalated with marine shallow-water carbonates(Shafaii Moghadam and Stern, 2014).The Eocene magmatic rocks are placed mainly south of the Late Cretaceous Sabzevar—Torbat-e-Heydarieh ophiolites, but also crosscutting the ophiolites and to the north (Shafaii Moghadam et al., 2015) (Fig.1B), which includes intermediate to felsic lava flows, adakites and pyroclastic rocks of Eocene age (Shafaii Moghadam et al.,2016).The Eocene carbonate rocks of the Sabzevar region are not defined under a formal lithostratigraphic nomenclature.Moreover, the exact stratigraphic relationships of the Eocene succession with the other lithostratigraphic units cropping out in the area remain unclear (Fig.1C).Shafaii Moghadam et al.(2015) noted that the Paleogene deposits of the southern Sabzevar region may show pulses of subduction-related(Cretaceous—Paleocene)and postorogenic magmatism (Eocene-Oligo-Miocene) within the Sabzevar magmatic belt.The oldest deposits of the southern Sabzevar region are Jurassic in age,composed of light brown oolitic limestone with Torinosuella peneropliformis Yabe and Hanzawa, Epistomina sp.,and Nautiloculina sp.The Lower Cretaceous rocks consist of gray argillaceous limestones and gray dolomitic-organodetrital limestones, with Orbitolina spp., Textulariella sp., Dicyclina sp., Bryozoa and algae (Rahaghi, 1980).The Upper Cretaceous is represented by different argillaceous and marly limestones, radiolarites and cherty limestones with abundant larger foraminifera (Siderolites calcitrapoides Lamarck, Orbitoides apiculatus Schlumberger, and Lepiorbitoides sp.) along with planktonic foraminifera (Heterohelix sp., Hedbergella spp., and Globotruncana spp., G.contusa, G.elevata, G.elevata stuartiformis), then topped by a Paleocene shallow-water succession (Rahaghi, 1980) characterized by Glomalveolina primaeva Reichel, Laffitteina sp., and Distichoplax biserialis (Dietrich).The Lower Eocene was explored by Rahaghi(1980),who reported Cuvillierina vallensis (Ruiz De Gaona),and Hadi et al.(2021), who studied the early Ypresian (SBZ6—SBZ7,NP10, CNE2) larger foraminifera.Rahaghi (1980) also reported some nummulitids and alveolinids from the Middle Eocene of the southern Sabzevar region.
The Chah-Talkh section is located about 9 km south of Oriyan village, 70 km south of Sabzevar and about 240 km southwest of Mashhad(Fig.1B).It can be found in the Darin and Sheshtamad 1/100,000 quadrangles(Fig.1C; sheet-7461, after Daneshmand and Nadim,1999; sheet-7561, after Jafarian and Jalali, 2000)(coordinates: 35°39′08′′N, 57°30′08′′E).The section is 58 m thick and is composed of different lithologies including conglomerate, marl, marly limestone, argillaceous limestone, silty sandstone, and sandy limestone.The marly intervals contain abundant and wellpreserved nummulitids (predominantly Assilina) and orthophragminids.Abundant Alveolina and Nummulites species occur throughout the middle part of the section.The interval under study comprises the upper part of the Chah-Talkh section, which is composed of about 17-m-thick marly and argillaceous limestones with abundant and well-preserved Alveolina.This stratigraphic portion is characterized by three main microfacies: (i) rotaliids and Nummulites packstone,(ii) Alveolina and Nummulites rudstone, and (iii) an Alveolina packstone (Fig.2).
3.Material and methods
A total of seven limestone samples (CT6—CT12)have been collected in vertical succession at about 3-m spacing from the Chah-Talkh section in Sabzevar region of central Iran.We have prepared forty-five thin sections from these samples (almost six thin sections per sample) labelled CT- (thin section dimension: 2.5 cm × 7.5 cm).We have further made 184 thin sections of isolated Alveolina(labelled ACT-)and Nummulites(labelled NCT-)specimens,and have selected 116 of them in perfect axial or centered section for a detailed study.The thin sections containing the studied specimens have been digitally photographed by using a binocular microscope under transmitted-light (Olympus BX53).The biometric parameters (proloculus diameter, axial and equatorial diameters, and elongation index) were measured transferring in-scale pictures on a single plate by using Adobe Illustrator CS4 software.We have followed Hottinger (1960, 1974), Drobne (1977), Samanta(1993) and Sirel and Acar (2008) for the taxonomic background analysis to identify the Alveolina species.The species of Nummulites were determined according to Schaub (1981).The thin sections are housed in the collection of the National Iranian Oil Company Exploration Directorate (NIOCEXP), Tehran, Iran and labelled as D.Pal.BF-3508.The biostratigraphy used refers to the SBZs of Serra-Kiel et al.(1998) and subsequent integrations (e.g., Silva-Casal et al.,2021; Rodriguez-Pint′o et al., 2022; Benedetti et al.,2023).
Fourteen smear slides were prepared from the seven samples (CN6—CN12)for calcareous nannofossil analysis following the techniques of Bown and Young(1998; modified by Parandavar and Hadavi, 2019).The samples were then analyzed using a standard light microscope (Olympus BX53) in parallel-polarized light(PPL) and cross-polarized light (XPL) with adding the Quartz Plate (QP) and Gypsum Plate (GP) at 1250—2000 × magnifications.Calcareous nannofossils were determined following the taxonomy of Romein(1979), Perch-Nielsen (1985), Bown (2005), and Aubry(1989).The biozonal frames used in this work refer to the low-latitude scheme of Martini (1971; NP biozones)and Agnini et al.(2014; CNE biozones).

Fig.2 Early Lutetian microfacies identified in the Chah-Talkh section, including texture and dominant and subordinate components.A)Rotaliids—Nummulites packstone (sample CT7/2); subordinate components: benthic foraminifera (Asterigerina, miliolids), Alveolina, and peloids; texture: packstone and locally grainstone matrix; B) Alveolina—Nummulites rudstone (sample CT8/1); subordinate components:peloids, miliolids, bivalves, small benthic foraminifera, and quartz; texture: rudstone with packstone and locally grainstone matrix; C, D)Alveolina packstone—rudstone (sample CT9/2 and CT9/3); subordinate components: benthic foraminifera (miliolids, textulariids and rotaliids), Orbitolites, Coskinolina, bivalve and gastropod fragments, and peloids; texture: packstone and rudstone with packstone matrix.A:Alveolina; C: Coskinolina; G: Bivalve and gastropod fragments; N: Nummulites; O: Orbitolites; R: Rotaliid (Neorotalia sp.).
4.Considerations on the Alveolina elliptica group
The Eocene Alveolina were classically grouped on the basis of their morphological(typological)features and in accordance with the inferred evolutionary trends through time (Hottinger, 1960, 1963, 1974;Drobne, 1977; Sirel and Acar, 2008).Hottinger (1974)recognized that some groups are not sufficiently known, especially concerning the growth stages that could be parameterized or not, depending on the evolutionary ‘speed’ of each group.So, gradual steps would be harder to study than sudden changes through time.This is also probably related to the occurrence of some poorly illustrated types (i.e.A.stercusmuris)that hamper thoughtful taxonomic comparisons.Among all, the Alveolina elliptica group is one of the most controversial examples.Its representatives are characterized by oval to subcylindrical forms with or without a weak flosculinization (Hottinger, 1974).Six allied taxa, i.e.A.elliptica (Sowerby), A.elliptica nuttalli Davies, A.pillai Checchia-Rispoli, A.stercusmuris Mayer-Eymar, A.cf.minuta Checchia-Rispoli,and A.lehneri Hottinger have been recognized by Hottinger (1960) within that group.The same author(Hottinger,1974),however,transferred A.lehneri and

Fig.3 Megalospheric forms of Alveolina specimens from the Chah-Talkh section(Sabzevar region,Central Iran).A—G)A.kieli Sirel and Acar,specimens number ACT7/1, ACT8/4, ACT8/5, ACT9/3, CT10/1, CT10/4, and CT11/4, respectively.
A.cf.minuta to the Alveolina levantina and Alveolina minuta group, respectively.Thus, he then formally placed only A.stercusmuris and A.elliptica in the Alveolina elliptica group,regarding A.javana Verbeek and A.elliptica nuttalli Davies as synonyms of A.elliptica (Sowerby).Sirel (1976) assigned the Lower Eocene A.haymanaensis Sirel to the Alveolina elliptica group.Later, Drobne (1977) and Drobne et al.(2011)placed only two species, A.elliptica nuttalli and A.stercusmuris,within this group.Hottinger and Drobne(1988) included three species in the group, A.elliptica, A.elliptica nuttalli, and A.stercusmuris,removing A.pillai from it.The most complete study on the Alveolina elliptica group from India (eastern Neo-Tethys) was carried out by Samanta (1993).He postulated the occurrence of four members:A.elliptica,A.nuttalli,A.pillai,and A.javana.Sirel and Acar(2008)introduced A.kieli Sirel and Acar supporting some close relationship with the members of the Alveolina elliptica group.The recognition of A.kieli Sirel and Acar from several central-eastern Neo-Tethys localities (Serra-Kiel et al., 2016; Hadi et al., 2020a;Rahman et al.,2021)could be considered to confirm its validity.The biometry of A.ozcani n.sp.is comparable to that of A.nuttalli, supporting its inclusion into the Alveolina elliptica group sensu Hottinger(1960,1974).This is,in particular,revealed by the parameters of the elongation index per whorl,illustrated in the following chapters of this work.A.ozcani n.sp.could be considered as an intermediate form between A.kieli and A.nuttalli, but the lack of meaningful elements for comparison with other allied forms such as A.elliptica (Sowerby) and A.pillai Checchia-Rispoli still hampers a precise evolutionary assessment.Ibrahimpaˇsi′c (2012) figured A.pillai along with A.nuttalli, A.elliptica, A.stercusmuris, and A.drobnella within the A.elliptica group from the Adriatic-Dinaridic carbonate platform.
5.Systematic paleontology
PhylumForaminifera d'Orbigny (1826).
OrderMiliolida Delange and Herouard, 1896
FamilyAlveolinidae Ehrenberg,1839
GenusAlveolina d’Orbigny (1826);
Type species.Oryzaria boscii Defrance in Bronn(1825), designated according to Opinion 1501 in the Bulletin of Zoological Nomenclature (1988), vol.45,part 3.The lectotype of Alveolina boscii has been selected by Tr′eguier et al.(2019).
Remarks.Alveolina occurs in shallow-marine sediments of the Late Paleocene and Early to Middle Eocene of the whole Neo-Tethys (e.g.Hottinger and Drobne,1988; Drobne et al., 2011; Zhang et al., 2013; Serra-Kiel et al., 2016; Haig et al., 2019).Alveolina species have been included in different groups on the basis of shell morphology and variations of the spiral growth in accordance with their growth stages, so that the evolutionary trends of these groups could most likely be arranged in parallel lineages (Hottinger, 1963, 1974;Drobne,1977;Sirel and Acar,2008).
Alveolina kieliSirel and Acar, 2008
Fig.3A—G.
1980 Alveolina stercusmuris-Rahaghi, pl.2, fig.5;1992 Alveolina stercusmuris-White, pl.3, figs.2—3;2008 Alveolina kieli-Sirel and Acar, p.81, pl.74, figs.1—3;2016 Alveolina kieli-Serra-Kiel et al.,p.18,textfig.17, 1—6; 2018 Alveolina sp.-¨Ozcan et al., p.181,text-fig.17,D—E;2020a Alveolina kieli-Hadi et al.,p.328, pl.5, figs.4—5; 2021 Alveolina kieli-Rahman et al., p.23, pl.3, figs.1—3; 2023 Alveolina kieli-Forouzande et al., pl.9,figs.1—5;pl.10, fig.1.Number of specimens examined: 53.
Description.The test is elongated-ovoid to slightly sub cylindrical with truncated poles.The proloculus is small and slightly oval with a diameter ranging between 110 μm and 212 μm (mean value:149.49 μm), followed by three to five tight whorls at the nepionic stage.The adult stage is composed of eight to ten whorls and the axial thickening is a bit thicker than that observed in the equatorial direction.The coiling of the last five to eight whorls is tight and the thickness of the basal layer stays constant till the final whorl.The intercalary chamberlets are somewhat recognizable in the axial region of the senile stage.The chamberlets are small, circular and regularly arranged, becoming large and upright ovoid in the outer whorls.Moreover,the secondary passages are barely observable in the axial region.The mean axial diameter is 4.73 mm, the mean equatorial diameter is 3.05 mm, and the mean elongation index(EI) is 1.63 at the 18thwhorl.Detailed measurements and ranges are illustrated in Table 1 and Supplementary Table S1.
Remarks.This species was introduced by Sirel and Acar (2008) from Turkey.Thereafter A.kieli has been reported elsewhere (e.g., Serra-Kiel et al., 2016;Rahman et al., 2021) and recognized as an endemic species of the Middle East (Hadi et al., 2020b).Ibrahimpaˇsi′c(2012,pl.32,Fig.5)informally described A.drobnella that,for us,closely resembles A.kieli with respect to its internal structure and test shape, and differs by its larger proloculus.A.stercusmuris Mayer-Eymer shows a very close relationship with A.kieli,with slight differences in the test shape(Figs.3 and 5)and elongation index(Fig.4).A.kieli is recorded from the central and eastern part of the Neo-Tethys Ocean of Turkey, Oman,Iran,and Pakistan,and is clearly distinguishable from A.stercusmuris by its lower elongation index(Fig.4)and marked ovoid test(Figs.3 and 5).

Table 1 Numerical characteristics in the axial section of Alveolina kieli Sirel and Acar A-forms from the Chah-Talkh section,and comparison with previous publications.The detailed data of the present study are available in the Supplementary Table S1.
Alveolina stercusmurisMayer-Eymar, 1886
Fig.5A—H.
1886 Alveolina stercusmuris-Mayer-Eymar, pl.4,Fig.10;1974 Alveolina stercusmuris-Hottinger,pl.61,fig.6; pl.71; 1977 Alveolina stercusmuris-Drobne, p.50, pl.10, figs.12—14; 1980 Alveolina stercusmuris-Rahaghi, pl.2, figs.1—4, 6—10; 1992 Alveolina stercusmuris-White, pl.3, fig.1; 2008 Alveolina stercusmuris-Sirel and Acar, p.83—84, pl.76, fig.1;2010 Alveolina stercusmuris-Deveciler, p.194, pl.3,fig.1; 2011 Alveolina stercusmuris-Drobne et al., p.751, pl.3; 2014 Alveolina stercusmuris-Deveciler, p.142, pl.4, fig.10; 2016 Alveolina stercusmuris-Serra-Kiel et al., p.344, fig.17 (7—9); 2020a Alveolina stercusmuris-Hadi et al., p.332, pl.3, figs.13—14;2021 Alveolina stercusmuris-Rahman et al.,p.23,fig.17 (e—g); 2023 Alveolina stercusmuris-Forouzande et al., pl.10, figs.2—5;pl.11, figs.1—2.
Number of specimens examined: 30.
Description.Test is subcylindrical with wellrounded and occasionally somewhat truncated poles.Proloculus is small with a diameter ranging from 110 μm to 190 μm (mean value: 154 μm), followed by three to five tightly coiled whorls at the nepionic stage.The following eight to nine whorls of the adult stage are coiled loosely displaying an increase in the thickness of the basal layer, especially axial thickening.The maximum thickness of the basal layer is 3.7 times greater than the height of the relative chamberlets.The last five to six whorls of the senile stage are coiled tightly whereas the thickness of the basal layer remains constantly thinner till the last whorl.The chamberlets are small and circular in the early stages; their size increases gradually through the ontogeny becoming significantly larger and oval to upright oval in the last stage of growth.The secondary passages and the intercalary chamberlets also occur in the axial sector of the middle and outer whorls.The mean axial and equatorial diameters are 6.40 mm and 3.48 mm at the 18thwhorl, with a mean elongation index of 1.84.Detailed measurements and ranges are illustrated in Table 2 and Supplementary Table S2.
Remarks.Hottinger (1974) noted that A.stercusmuris Mayer and Eymar differs from all other taxa included into the Alveolina elliptica group by the lack of flosculinization (except of A.kieli Sirel and Acar, 2008).This feature is observable in our specimens especially if comparing the flosculinized forms of A.nuttalli and A.ozcani n.sp.In addition, this species differs from A.kieli by having a more developed basal layer that, in turn, is reflected by the test shape (Fig.5) and the greater elongation index (Fig.4).

Fig.4 Comparison of biometric parameters between Alveolina kieli Sirel and Acar and Alveolina stercusmuris Mayer-Eymar from the Chah-Talkh section(Sabzevar region,Central Iran).A)Variation of mean axial and equatorial diameters in successive whorls in A.kieli;B)Variation of mean axial and equatorial diameters in successive whorls in A.stercusmuris;C)Elongation index in successive whorls of A.kieli compared with that in A.stercusmuris; minimum and maximum values are plotted.The source data are available in Supplementary Tables S1 and S2.
Alveolina nuttalli (Davies, 1940)
Fig.6A—H.
1940 Alveolina elliptica nuttalli-Davies,p.219—220,pl.12, figs.1—4; 1960 Alveolina elliptica nuttalli-Hottinger, pl.12, fig.4; 1977 Alveolina elliptica nuttalli-Drobne, pl.10, figs.9—11; 1980 Alveolina elliptica nuttalli-Rahaghi, pl.5, figs.1—10; 1993 Alveolina nuttalli-Samanta,p.65—72,pl.10,figs.1—5;pl.11,figs.2—3;2010 Alveolina nuttalli-Deveciler,pl.3,figs.3—5;2011 Alveolina elliptica nuttalli-Drobne et al.,pl.3;2014 Alveolina nuttalli-Deveciler,pl.4,figs.1—3;2016 Alveolina elliptica nuttalli-Serra-Kiel et al.,p.344,text-fig.17 (10—13);2018 Alveolina spp.-¨Ozcan et al.,p.146,text-fig.8(16,19—20);2019c Alveolina elliptica nuttalli-Hadi et al.p.151,text-fig.6.4;2020a Alveolina elliptica nuttalli-Hadi et al., pl.5, figs.6—8; 2021 Alveolina elliptica nuttalli-Rahman et al.,p.23,fig.18(a—i); 2023 Alveolina elliptica nuttalli-Forouzande et al.,pl.11,figs.3—5.

Present study 110—190 16—20 3.6—6.5 1.74—3.47 1.87—2.1 Forouzande et al.(2023)125—225 13—22 5.7—8.7 2.84—4.69 1.85—2 Rahman et al.(2021)120—200 13—18 4.61—6.08 2.41—3.36 1.68—1.91 Hadi et al.(2020a)180—200 15—20 7.8—8.4 4.1—4.4 1.8—1.9 Serra-Kiel et al.(2016)230—300 15—18 7.35 (17th whorl)2.80 (17th whorl)2.58 (17th whorl)Deveciler(2014)148 134.97 2.75 1.88 Deveciler(2010)255 155.63.11.81 200(2008)Sirel and Acar 165.52.91.89 Drobne(1977)125—200 16—18 6—7.5 3.2—4.2(16th—18th whorl)150—225(1974)13—20——1.75—2.8 1.55—2.15 Number of whorls Alveolina stercusmuris Hottinger Proloculus diameter(μm)Axial diameter(mm)Equatorial diameter(mm)Elongation index
Number of specimens examined: 21.
Description.The test is elongated-ovoid to slightly subcylindrical in shape with rounded poles.The proloculus is small with a diameter of 100—210 μm(mean value: 147.5 μm).The first four to six whorls of the nepionic stage are coiled tightly.The adult stage is composed of three to four highly flosculinized whorls.The maximum thickness of the basal layer is five to eight times greater than the height of the axial chamberlets.The senile stage of growth is composed of four to nine whorls and shows an abrupt thinning of the basal layer.Poles are slightly depressed inwards,in the axial sector of the senile stage.The chamberlets are circular and densely packed in the inner whorls,then they become larger and oval in the outer whorls.The mean axial diameter is 4.54 mm and the mean equatorial diameter is 3.00 mm at the 13thwhorl,with a mean elongation index of 1.49.Detailed measurements and ranges are illustrated in Table 3 and Supplementary Table S3.
Remarks.A.nuttalli is easily distinguished from allied forms (e.g.the advanced form of A.elliptica sensu Hottinger, 1974) by having a smaller test and three more distinguishable growth stages.It also differs from A.pillai in its smaller size and lower number of whorls in the nepionic and senile stages.However,Hottinger (1974) regarded A.pillai as a junior synonymy of A.elliptica.Our specimens are identical to all the various morphotypes of A.nuttalli (Davies) so far reported, which strictly show fewer whorls in the nepionic stage than A.ozcani n.sp.
Alveolina ozcaniHadi and Consorti n.sp.
Figs.7A—G, 8A—D.
1993 Alveolina pillai-Samanta, pl.13, fig.5; 2010 Alveolina nuttalli-Deveciler, pl.3, fig.3; 2012 Alveolina pillai- Ibrahimpaˇsi′c, pl.28, figs.1—5, and pl.29,figs.1 and 3.
Derivation of name.The species is dedicated to Ercan ¨Ozcan, who extensively worked on larger foraminifera.
Type material.Holotype (Fig.7A and 9) and paratypes (Fig.7B—G) from samples ACT8/1 and ACT9/3 are deposited in the collection of the National Iranian Oil Company Exploration Directorate (NIOCEXP), Tehran, Iran under the label D.Pal.BF-3508.

Fig.6 Megalospheric forms of Alveolina specimens from the Chah-Talkh section (Sabzevar region, Central Iran).A—H) A.nuttalli (Davies),specimens number ACT7/9, ACT8/1.24, ACT9/3.41, ACT9/43, ACT9/45, ACT9/4851, ACT9/53, and ACT9/54, respectively.
Type locality.Chah-Talkh section, southern Sabzevar region, Central Iran.
Type level.Early Lutetian, SBZ13.
Material.Samples CT8 and CT9 from the Chah-Talkh section (Figs.7 and 8).
Number of specimens examined: 14.
Description.The test is ovoid to subcylindrical in shape, with rounded poles.The diameter of the proloculus is small,ranging between 110 and 205 μm,followed by seven to eleven oval and tight, nonflosculinized whorls of the nepionic stage of growth.This is followed by the adult stage that is flosculinized and composed of two to three loose whorls,showing a marked increase of the basal thickening.Basal layer can be five to six times thicker than the height of the chamberlets in the axial region, where the secondary passages are visible.The last four to seven whorls of the senile stage are coiled tightly with a constantly thin basal layer.Few of the intercalary chamberlets occur axially, in the senile stage.The chamberlets are small and circular, irregularly arranged and densely packed.The size of the chamberlets increases gradually during ontogeny becoming larger and oval to upright oval in the outer whorls.The mean axial and equatorial diameters are 4.89 mm and 2.61 mm, respectively, at the 15thwhorl, with a mean elongation index of 1.97.Detailed measurements and ranges are illustrated in Table 4 and Supplementary Table S4.

Table 3 Numerical characteristics in the axial section of A.nuttalli (Davies)A-forms from the Chah-Talkh section,and comparison with previous publications.The detailed data of the present study are available in the Supplementary Table S4.
Differential diagnosis.A.ozcani n.sp.is biometrically similar to A.nuttalli (Davies) with respect to the proloculus size, test shape, and elongation index, but it differs by having a greater number of tightly coiled whorls in the nepionic stage of growth (Table 4; Figs.9 and 10).There are some similarities between A.ozcani n.sp.and A.kieli regarding the style of the early stage of growth(nepionic whorls), but the adult basal layer of the axial and equatorial zones of A.ozcani n.sp.shows a significant development(Fig.10).A.pillai Checchia-Rispoli shows a similar arrangement of the whorls throughout ontogeny - with emphasis on 5—14 tightly to loosely coiled whorls of the nepionic stage- although A.ozcani n.sp.differs by the smaller and fewer whorls of the senile stage.Furthermore, A.ozcani differs from A.pillai by having a wider nepionic stage and loosely coiled senile whorls arranged in a short series of coarser chamberlets.We wish to stress that A.ozcani n.sp.differs from A.pillai also in possessing a generally coarser internal structure and by the occurrence of some intercalary chamberlets that are somewhat recognizable in the senile stage as well.
6.Biostratigraphy
The biostratigraphic framework of the studied section is erected by integrating the record of larger foraminifera and calcareous nannofossils.This is usually applied where both fossil groups are found together (e.g., in NE Italy and Turkey; see Benedetti et al., 2023; Yücel et al., 2023) and may represent an improved step for the Eocene successions of Iran,the studies of which usually provide only a low resolution (e.g., Babazadeh and Cluzel, 2023).Regarding larger foraminifera, we focused on the stratigraphic value of species of Alveolina and Nummulites that are used for correlations with the SBZs of Serra-Kiel et al.(1998).Although SBZs are fairly well positioned against the global standards (linked both to calcareous plankton and nannofossil scales and to the magnetostratigraphic units), Rodriguez-Pint′o et al.(2022) and Benedetti et al.(2023), among others, have pointed out that some of SBZs boundaries need to be calibrated more carefully.The calcareous nannofossil standard biozonation (Martini, 1971; Okada and Bukry, 1980;Agnini et al., 2014) represents one of the most reliable tools for correlating the larger foraminifera biostratigraphy to the global time scale (Serra-Kiel et al., 1998; Berggren and Pearson, 2005), which may satisfy some of requirements to fill the stratigraphic uncertainties of the shallow-water successions(e.g., Pignatti and Papazzoni, 2017; Simmons and Aretz, 2020).
6.1.Larger foraminifera

Fig.7 Megalospheric forms of Alveolina ozcani n.sp.from the Chah-Talkh section(Sabzevar region,Central Iran).A)Holotype;axial section,specimen number ACT8/1.10;B—G)Paratypes;axial sections,specimens number ACT8/1.13,ACT8/1.25,ACT8/1.26,ACT8/1.27,ACT8/1.29,and ACT9/3.44, respectively.
Four Alveolina species, A.kieli Sirel and Acar, A.stercusmuris Mayer-Eymar,A.nuttalli(Davies),and A.ozcani n.sp.have been found along with Nummulites uroniensis (de la Harpe) emend.Heim, N.obesus Sheng and Zhang, and N.cf.verneuili d'Archiac and Haime (Fig.11).Coskinolina sistanensis Schlaginweit and Hadi, Fabiania cassis Oppenheim, Orbitolites sp.,and Asterigerina sp.also occur all together, which represent an inedited assemblage,first time recorded simultaneously in the central Neo-Tethys region.Based on the occurrence of the indices N.uroniensis and N.obesus, the stratigraphic succession studied here can be placed into the SBZ13 zone (early Lutetian; Serra-Kiel et al., 1998) (Fig.12).Concerning Alveolina,Sirel and Acar (2008), Deveciler (2010, 2014) and Forouzande et al.(2023)placed the occurrences of A.kieli, A.stercusmuris, and A.nuttalli approximately into the middle Lutetian—early Bartonian(SBZ15—SBZ17).In the southern basins of the central Neo-Tethys of Oman, the stratigraphic range of A.nuttalli can be extended up to SBZ16 (late Lutetian;Serra-Kiel et al., 2016).Hadi et al.(2020a) and Rahman et al.(2021) supported the occurrence of these Alveolina species into the SBZ13 of Serra-Kiel et al.(1998) of the central Neo-Tethys (SE Iran) and the eastern Neo-Tethys (Pakistan) realms.This suggests that the stratigraphic range of A.kieli, A.stercusmuris, and A.nuttalli is wide, ranging throughout the SBZs 13 to 17, whereas the Nummulites assemblage looks reliable to make more precise correlations with the SBZs.A.ozcani n.sp.occurs in association with Nummulites and calcareous nannofossil markers of the SBZ13 (Fig.12), but assessment of its full stratigraphic range needs further research.

Fig.8 Megalospheric forms of Alveolina ozcani n.sp.from the Chah-Talkh section (Sabzevar region, Central Iran) in axial and sub-axial section.Specimens number ACT8/1.40, ACT8/1.8, ACT9/1.18, and ACT9/3.51, respectively from A to D.
6.2.Calcareous nannofossils
The calcareous nannofossil assemblages are well preserved and suffered only limited diagenesis as justsome etching, dissolution or calcite overgrowth are observed.The preservation index can be assigned to the E1 and O1 of Roth(1973)scheme.The assemblages are dominated by placoliths such as Coccolithus, Reticulofenestra, and Cyclicargolithus (together up to 70 %of the total specimens) along with some reworked Cretaceous specimens (e.g.Watznaueria, Prediscosphaera,and Cribrosphaerella).In Figs.13 and 14,we have illustrated the most significant taxa, along with some common species.Coccolithus pelagicus(Wallich),Reticulofenestra dictyoda (Deflandre), and R.minuta Roth are the most abundant taxa (about 75 %) in the studied samples.

Table 4 Comparison of growth stages of A-forms generations of A.ozcani n.sp.and A.nuttalli(Davies).
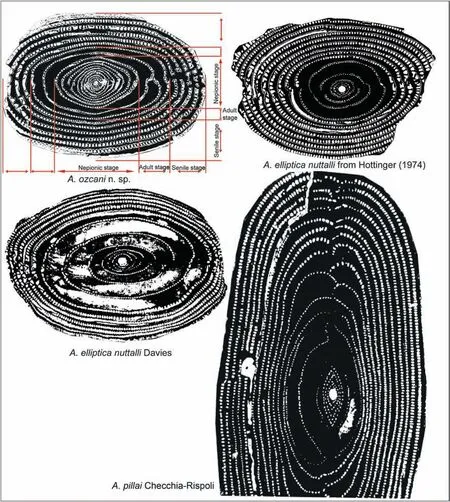
Fig.9 Holotype of A.ozcani n.sp.(with biometric parameters), the holotype of A.nuttalli (Davies), the A.nuttalli (Davies) figured in Hottinger (1974), and A.pillai Checchia-Rispoli (specimen in fig.12 of Checchia-Rispoli, 1909).
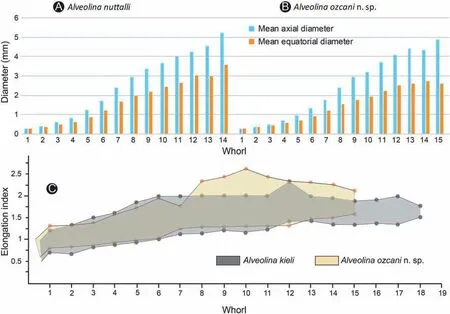
Fig.10 Biometric parameters of Alveolina ozcani n.sp.compared to that of A.nuttalli(Davies)and A.kieli,Chah-Talkh section(Sabzevar region,Central Iran).A)Variation of mean axial and equatorial diameters in successive whorls of A.nuttalli; B) Variation of mean axial and equatorial diameters in successive whorls of A.ozcani n.sp.;C)Elongation index in successive whorls of A.ozcani compared with that of A.kieli; minimum and maximum values are plotted.The source data are available in the Supplementary Tables S1, S3, and S4.
Further species determined are Coccolithus formosus (Kamptner), C.pauxillus Bown, Chiasmolithus sp., Discoaster cf.kuepperi, D.saipanensis Bramlette and Riedel, Nannotetrina sp., Micrantholithus disculus(Bramlette and Riedel),Sphenolithus moriformis(Bronnimann and Stradner), Sphaerogypsina editus Perch-Nielsen, S.radians Deflandre, S.spiniger Bukry,Toweius callosus Perch-Nielsen,T.pertusus(Sullivan),T.gammation (Bramlette and Sullivan), Cyclicargolithus floridanus(Roth and Hay),Reticulofenestra erbae(Fornaciari et al.),R.bisecta(Hay et al.),R.wadeae Bown,and Helicosphaera reticulata Bramlette and Wilcoxon.We suspect that some of these species were most likely reworked from older stratigraphic levels(e.g.,Ypresian stage)thusweprefertorelyontheyoungerassemblages,among them C.formosus,T.pertusus,and S.editus.
The NP14 zone of Martini (1971) is defined by the lowest appearance (LA) of Discoaster sublodoensis up to the LA of Nannotetrina fulgens and/or the highest appearance (HA) of Blackites inflatus (= Rhabdosphaera inflata).Okada and Bukry (1980) divided this zone into two subzones CP12a (Ypresian age) and CP12b (Lutetian age) (equivalent to NP14a and NP14b subzones)based on the LA of Blackites inflatus and/or any species of the genus Nannotetrina.Subsequently,Perch-Nielsen (1985), Bown and Jones (2012) and Howe (2021) have shown the LA of Sphenolithus spiniger in the NP14b subzone,within the Lutetian stage.Moreover, The LA of D.sublodoensis to HA of D.lodoensis,and then the LA of N.cristata and/or LA of B.inflatus were used to define the zones CNE6 and CNE7 (equivalent to the NP14a subzone) of Agnini et al.(2014), respectively (Fig.11).In addition,Agnini et al.(2014)used the LA of N.cristata and LA of N.alata for defining the lower and upper boundaries of the CNE8 zone (equivalent to the NP14b subzone),respectively.
The LA of Nannotetrina(N.fulgens or N.quadrata)was classically thought as the lower boundary of the NP15 zone (Martini, 1971; Perch-Nielsen, 1985), but Varol (1989) and Agnini et al.(2014) found such taxa within the NP15 zone.The LAs of N.alata and B.gladius point to the lower boundaries of CNE9 and CNE8b zones of Agnini et al.(2014) and Varol (1989),respectively(Fig.12).It should be mentioned that the base of CNE9 can be correlated with the lower boundary of the NP15 zone of Martini (1971) and with the lower boundary of the CP13a subzone of Okada and Bukry (1980) (Fig.12).Thus, based on the subdivision of the NP15 zone made by Okada and Bukry (1980)throughout the total vertical range of Chiasmolithus gigas (NP15a to NP15c and/or CP13a and CP13b subzones),the LA of Coccolithus gigas(=Ch.gigas)could be used to define the base of CP13b and CNE10 zones of Okada and Bukry (1980) and Agnini et al.(2014),respectively (Fig.12).

Fig.11 Equatorial and axial sections of megalospheres of Nummulites specimens from the Chah-Talkh section (Sabzevar region, Central Iran).A—D)N.uroniensis(de la Harpe)emend.Heim:(A1)external view,(A2—B2)equatorial sections,(C,D)axial sections,sample NCT9/1,NCT9/2,NCT9/4,and NCT9/6,respectively;E,F)N.cf.verneuili d’Archiac and Haime:(E1 and F1)external view,(E2,E3 and F2)equatorial sections, sample NCT12/1 and NCT12/2, respectively; G) N.obesus Sheng and Zhang: (G1 and G2) equatorial section, sample NCT12/3.
The lowermost samples of our succession(samples CT6 and CT7) can be assigned to the NP14b subzone based on the occurrence of Nannotetrina sp.and S.spiniger(Fig.12).The intermediate part of the studied column (sample CT8) can be assigned to the NP15a subzone based on the occurrence of C.floridanus along with R.wadeae.Although the latter species do not necessarily suggest zonal separation, it is inserted to approximately compare our record with that of Bown and Jones (2012).Our first record of H.reticulata could be considered as an LA,thus suggesting that the upper part of studied interval (samples CT9 to CT12)can be assigned to the NP15b subzone of Martini(1971).The whole biozonal scheme here recognized indicates an early Lutetian age for the studied interval.This is equivalent to the CNE8—CNE10 zones of Agnini et al.(2014), to the CP12b—CP13b zones of Okada and Bukry(1980)from low latitudes,and to the NNTE8 zone of Varol(1989)from mid to high latitudes(Fig.12).

Fig.12 A)Lithological column of the Chah-Talkh section(Sabzevar region,Central Iran);B)Biostratigraphy and correlation between larger foraminifera and calcareous nannofossils biozones; C) Biozonational schemes around the Ypresian—Lutetian boundary with chronostratigraphic attribution of the studied succession.
7.Discussion

Fig.13 Index and commonly recorded calcareous nannofossils from the Chah-Talkh section(Sabzevar region,Central Iran).Scale bar is 2 μm for all images.A,B)Coccolithus pelagicus(A:XPL,B:GP);C,D)Reticulofenestra dictyoda(C:XPL,D:GP);E—H)Sphenolithus spiniger(E and G:XPL,F and H:GP,E and F:0°,G and H:45°);I—L)Sphenolithus villae(I and K:XPL,J and L:GP,I and J:0°,K and L:45°);M-P)Sphenolithus radians(M and O:XPL,N and P:GP,M and N:0°,O and P:45°);Q—T)Sphenolithus moriformis(Q and S:XPL,R and T:GP,Q and R:0°,S and T:45°).Abbreviations: XPL = Cross-polarized light; GP = Gypsum plate.
The stratigraphic arrangements published in the Geological Time Scale (Gradstein et al., 2020) for the Eocene Series suggest placing the Ypresian—Lutetian transition, in agreement with Serra-Kiel et al.(1998), at the SBZs12/13 biozonal boundary.This boundary, however, looks doubtful (Speijer et al.,2020, Fig.28.6) and indeterminately assigned, at the Lutetian GSSP locality, to a generic upper Ypresian—lower Lutetian timespan.By integrating parallel stratigraphic methods (magnetostratigraphy and nannofossil biostratigraphy) against the SBZs of Serra-Kiel et al.(1998), Rodriguez-Pint′o et al.(2022)and Benedetti et al.(2023) have finally suggested to place this line within the lower Lutetian stage.The biostratigraphy presented here made by integrating the vertical distribution of larger foraminifera and calcareous nannofossils point to the SBZ13.This is particularly supported by the occurrence of the SBZs biozonal markers Nummulites uroniensis and N.obesus, which can be considered more reliable to make correlations among the western and central Neo-Tethys than the Alveolina species recovered here.This is supported by integrating the occurrence of nannofossil taxa referable to the NP14 and NP15 zones, and more in particular the CNE8 to 10 zones and CP12b to CP13b subzones.By comparing our results with those displayed in Fig.12, we are able to confirm the placement of our section in the SBZ13, especially if assuming the SBZ12/13 boundary, as suggested by Rodriguez-Pint′o et al.(2022) and Benedetti et al.(2023), in the NP14 zone or in the upper part of the CP12b subzone (see Speijer et al., 2020).A refined correlation would suggest that the base of the studied section would fall into the uppermost part of the CP12b subzone, more likely above the suggested SBZ12/13 boundary.The rest of the section comprises the NP15a and NP15b subzones, therefore ranging from the upper part of the lower Lutetian possibly to at least the lower part of the middle Lutetian.
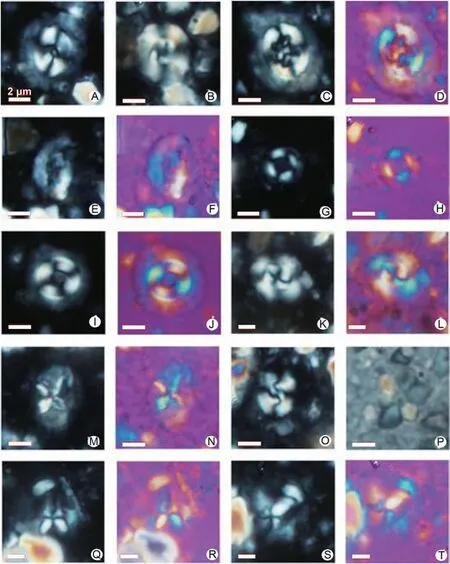
Fig.14 Index and commonly recorded calcareous nannofossils from the Chah-Talkh section(Sabzevar region,Central Iran).Scale bar is 2 μm for all images.A) Coccolithus pelagicus (XPL); B) Cyclicargolithus floridanus (XPL); C, D) Chiasmolithus sp.(C: XPL, D: GP); E, F) Helicosphaera reticulata (E: XPL, F: GP); G, H) Coccolithus pauxillus (G: XPL, H: GP); I, J) Coccolithus formosus (I: XPL, J: GP); K, L) Reticulofenestra erbae(K:XPL,L:GP);M,N)Nannotetrina sp.(M:XPL,N:GP);O)Reticulofenestra bisecta(XPL);P)Discoaster cf.kuepperi(QP);Q—T) Sphenolithus spiniger (Q and S: XPL, R and T: GP, Q and R: 0°, S and T: 45°).Abbreviations: XPL = Cross-polarized light; GP = Gypsum plate; QP = Quartz plate.
Some ambiguities still exist on how to define the members of Alveolina elliptica group,as the clustering philosophy changed at times (e.g., Hottinger, 1974;Samanta, 1993; Sirel and Acar, 2008).It is, however,possible to state that some members of the group possess a wide geographic distribution throughout the Neo-Tethys domain, from Europe and the Mediterranean(western Neo-Tethys)to the Indo-Pacific(eastern Neo-Tethys) basins (e.g., Hottinger, 1960; Drobne,1977; Samanta, 1993) (Fig.15).A.nuttalli appears having a wider biogeographic range (from the Mediterranean to Indo-Pacific regions) compared to other members of the group (Table 5).This species is markedly flosculinized in contrast with the weakly or/and non-flosculinized specimens included into the Alveolina elliptica group such as A.elliptica and A.stercusmuris sensu Hottinger (1974).Therefore, the close morphological and biometrical correspondences between A.nuttalli and A.ozcani n.sp.suggest to focus on the paleobiogeographic significance and stratigraphic distribution of the well-known A.nuttalli.
A.nuttalli was first described as A.elliptica var.nuttalli from the Indio—Pakistan region by Davies(1940).He considered both of A.elliptica var.a and A.elliptica var.b of Nuttall (1925) as synonyms of his new form from Kohat,while Samanta(1993)suggested that they belong to two different species: A.nuttalli(Davies)and A.ovoidea d’Orbigny.In the western Neo-Tethys in contrast, there are only a few reports from the Middle Eocene, mostly concentrated on the Adriatic Carbonate Platform of the Mediterranean area(Hottinger, 1960; Drobne, 1977).There, Drobne et al.(2011) demonstrated that the stratigraphic range of A.nuttalli (Davies) can be assigned to the Lutetian(SBZ13—SBZ16), whereas it is also reported from Turkey in association with Bartonian (SBZ17) markers such as A.fragilis Hottinger, A.fusiformis Stache, N.perforatus (de Montfort), and N.malatyaensis Sirel(Deveciler, 2010, 2014).Serra-Kiel et al.(2016) found A.nuttalli along with N.garganicus Tellini, N.discorbinus (Schlotheim), and N.somaliensis Nuttall and Brighton confirming that the stratigraphic range comprises the early Lutetian(SBZ 13)to late Lutetian(SBZ 16) timespan also in the central Neo-Tethys (Yemen and Oman).A.nuttalli was reported from the Alborz area (NW Iran) in the Ypresian (SBZ8—SBZ11) in association with A.gr.guidonis,A.cf.distefanoi Chechia-Rispoli,A.cf.minuta Chechia-Rispoli,Sphaerogypsina globulus (Reuss), and Coskinolina sistanensis (Hadi et al., 2019d).Hadi et al.(2020a) recorded A.nuttalli from the Hormak section(eastern Iran)along with other members of the A.elliptica group such as A.kieli and A.stercusmuris,from a level tentatively dated as early Lutetian (SBZ13).The same assemblage, which includes A.nuttalli, A.kieli, and A.stercusmuris, is also known from the Jahrum Formation(Zagros region)from the middle Lutetian (SBZ15) to the early Bartonian (SBZ17) by the occurrence of nummulitid and orthophragminid indices (Forouzande et al., 2023).
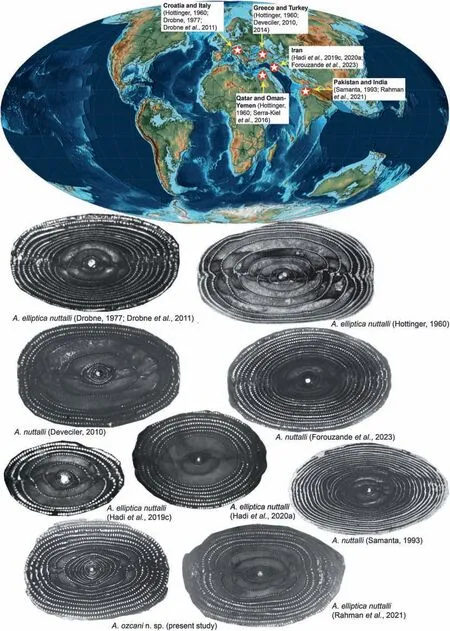
Fig.15 Biogeographic distribution of Alveolina nuttalli Davies throughout the Neo-Tethys (paleogeographic map from Scotese, 2014).
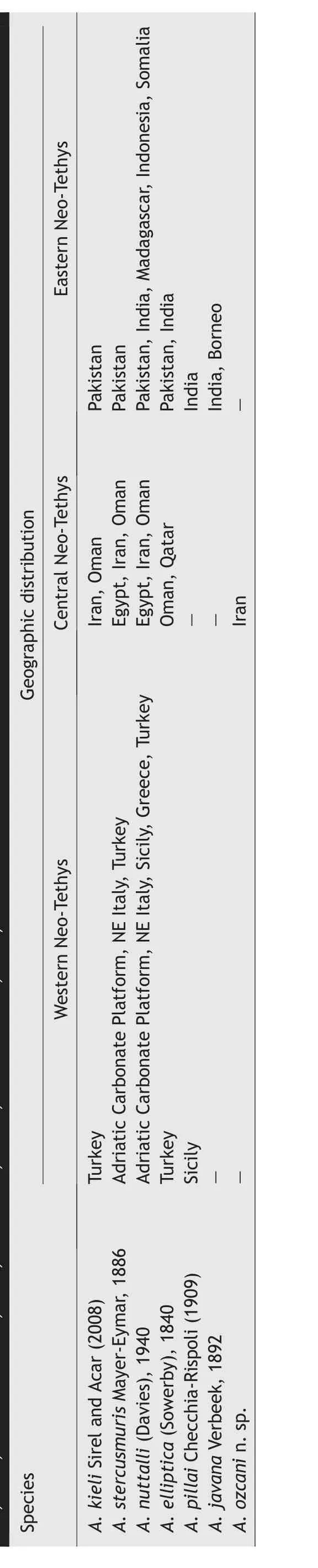
Eastern Neo-Tethys Pakistan Pakistan Pakistan, India, Madagascar,Indonesia,Somalia Pakistan, India India India,Borneo Egypt,Iran, Oman Iran,Oman Geographic distribution Egypt,Iran, Oman Oman,Qatar Turkey Central Neo-Tethys Adriatic Carbonate Platform,NE Italy,Turkey Western Neo-Tethys Adriatic Carbonate Platform,NE Italy,Sicily, Greece, Turkey Turkey Sicily——Iran——Species A.kieli Sirel and Acar(2008)A.stercusmuris Mayer-Eymar,1886 A.nuttalli(Davies),1940 A.elliptica(Sowerby), 1840 A.pillai Checchia-Rispoli(1909)A.javana Verbeek,1892 A.ozcani n.sp.—
Our study agrees with the Middle East records of A.nuttalli in the SBZ13 first given by Hottinger(1960)and Rahaghi(1980)from Qatar and Iran,respectively.In the eastern Neo-Tethys (Pakistan), A.nuttalli is reported within the SBZ12 to SBZ13 in association with A.cosinensis cosigena, A.aff.frumentiformis,A.aff.azzarolii Drobne, A.kieli, and A.stercusmuris (Rahman et al., 2021).Samanta (1993) provides the most complete collection of the Alveolina elliptica group members from the upper part of the Fulra limestone in India(eastern Neo-Tethys region) reporting A.nuttalli in association with A.elliptica,A.pillai,A.javana,and A.ovoidea, along with other nummulitid and orthophragminid markers,which are attributed to the Middle Eocene.From the same Fulra limestone, ¨Ozcan et al.(2018) illustrated some Bartonian (SBZ17) specimens assignable to A.nuttalli,A.elliptica, and A.ovoidea.The A.elliptica group is common throughout the Lutetian, their complete range may even cover the Ypresian—Bartonian timespan.The confirmed stratigraphic occurrences of A.nuttalli support this last statement.Conversely,A.ozcani n.sp.is only reported from the Chah-Talkh section so far,and may suggest a probable endemism in the Middle East platforms.
8.Conclusions
A detailed study of the Eocene shallow-water carbonate succession of the Chah-Talkh section(Central Iran) allowed identifying A.kieli, A.stercusmuris,and A.nuttalli as members of the Alveolina elliptica group.The assemblage is considered as early Lutetian in age by the occurrence of the indices Nummulites uroniensis and N.obesus, along with N.cf.verneuili (SBZ13).Such an attribution is corroborated by applying the calcareous nannofossil biostratigraphy, with which the NP14b—NP15b(CNE8—CNE10) zones were identified.Thus, for the first time so far, the index zonal calcareous nannofossil markers such as S.spiniger,Nannotetrina sp.,C.floridanus, R.wadeae, and H.reticulata were recognized from a shallow-water succession of Central Iran (central Neo-Tethys region) and correlated with SBZs.These results have been compared with recent data published on the Lower—Middle Eocene chronostratigraphy.This suggests the studied succession falls within the SBZ13,from the upper part of the early Lutetian to the lower part of the middle Lutetian.Our study also confirms that the genus Nummulites can be considered a more reliable marker than Alveolina to correlate Middle East shallow-water successions with the Shallow Benthic Zones.Furthermore, we have introduced a new Alveolina species, A.ozcani n.sp., as a further representative of the Alveolina elliptica group.It is characterized by more whorls in the nepionic stages with respect to its close allies.Unlike Alveolina nuttalli, we postulate the new species may be an endemic taxon such as A.kieli, restricted to Middle East platforms.However,more records are needed to confirm such a biogeographic affinity by considering populations in timeequivalent strata elsewhere (i.e.further east),which are currently unavailable.
Authors'contributions
MH: Fieldwork and sampling, conceptualization,methodology, formal analysis, investigation, writingoriginal draft, writing-review and editing.LC:Conceptualization, investigation, writing-original draft, writing-review and editing.AB: Investigation,writing-original draft and funding acquisition.MP:Conceptualization, investigation, writing-original draft.SK: Formal analysis.All authors read and approved the final manuscript.
Funding
This study was supported financially by the Vice Chancellor for Research and Technology at University of Isfahan.
Conflicts of interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgements
This study is carried out within the framework of a postdoc project given to MH.The authors are grateful to the Vice Chancellor for Research and Technology at University of Isfahan for financial and logistic supports.We are grateful to Katica Drobne and Ali Deveciler for the helpful suggestions and constructive comments that greatly improved the quality of this paper.Furthermore, we would especially like to thank the editor, Xiu-Fang Hu, for editorial comments.
Supplementary data
Biometric parameters of Alveolina kieli, A.stercusmuris, A.nuttalli, and A.ozcani n.sp.for this research are available in the Supplementary Tables S1—S4, respectively, and can be found online at https://doi.org/10.1016/j.jop.2023.11.002.
 Journal of Palaeogeography2024年1期
Journal of Palaeogeography2024年1期
- Journal of Palaeogeography的其它文章
- Biological sources, paleoenvironment, and organic matter enrichment in source rocks of the Ordovician Majiagou Formation, Ordos Basin, China: Evidence from biomarkers,microfossils, and inorganic geochemical analyses
- On the occurrence of Neoplanorbulinella Matsumaru, 1976 (Foraminifera) from the late Oligocene of Central Iran (Qom Formation):Palaeobiogeographic implications
- Circum-Indian Holocene sea-level database:A repository of distinct relative sea-level plots across the Indian Peninsula
- Evolution of Cenozoic sedimentary architecture in Central and Southern South China Sea basins
- Interpreting siliciclastic sedimentation in the upper Paleozoic Mulargia-Escalaplano Basin(Sardinia, Italy): influence of tectonics on provenance
- Trace fossil evidences of an Early Miocene paleoseismic event and depositional regime change from the Kutch (Kachchh) Basin